
Influences of Seasonal Variability and Potential Diets on Stable Isotopes and Fatty Acid Compositions in Dominant Zooplankton in the East Sea, Korea
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of Environmental Parameter
2.2. Zooplankton Sampling
2.3. Stable Isotope Analysis of Zooplankton
2.4. Fatty Acid Composition Analysis
2.5. Statistical Analysis
3. Results
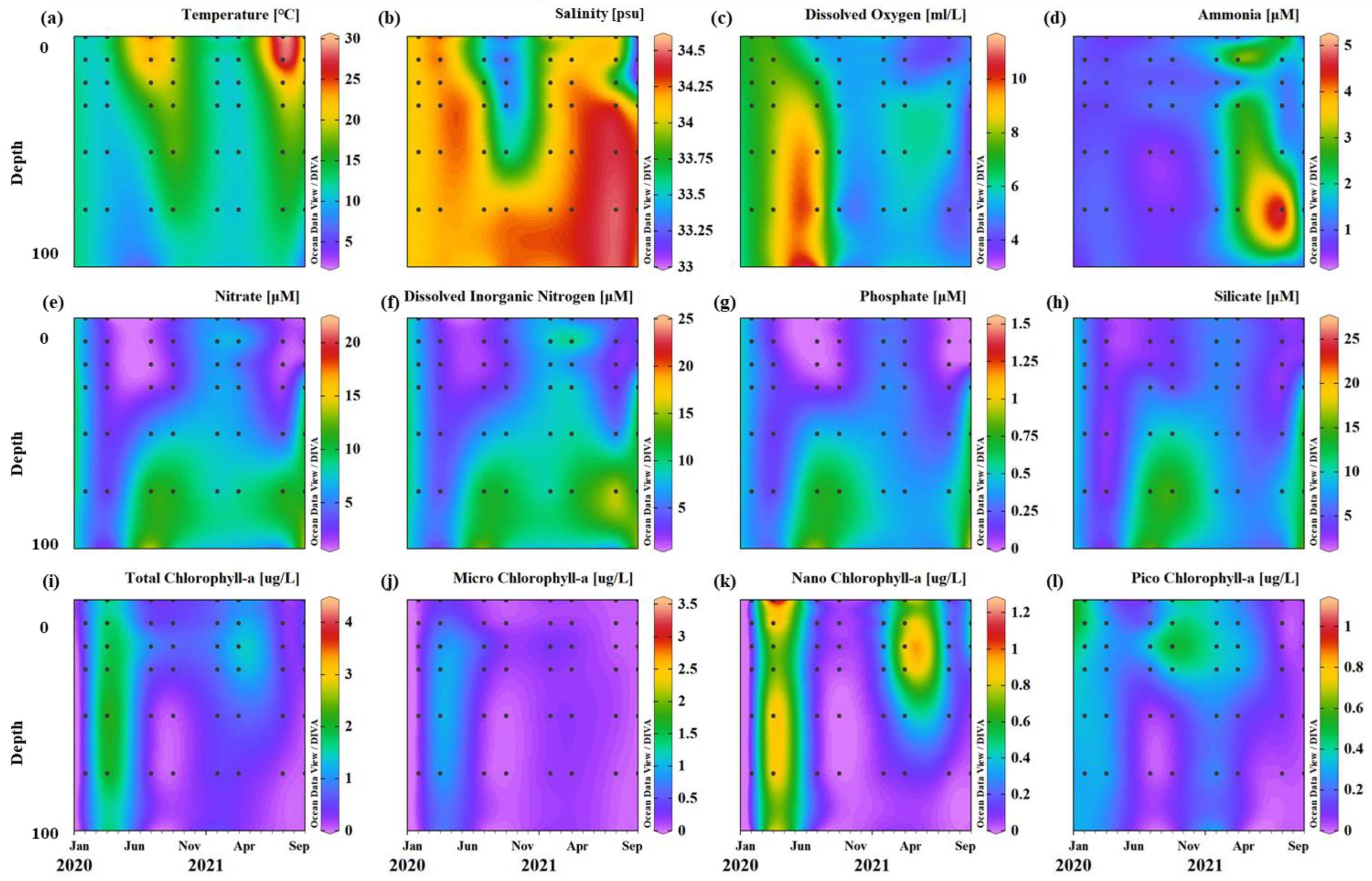
3.1. Environmental Conditions
3.2. Spatiotemporal Isotope Fluctuations in Zooplankton Taxa
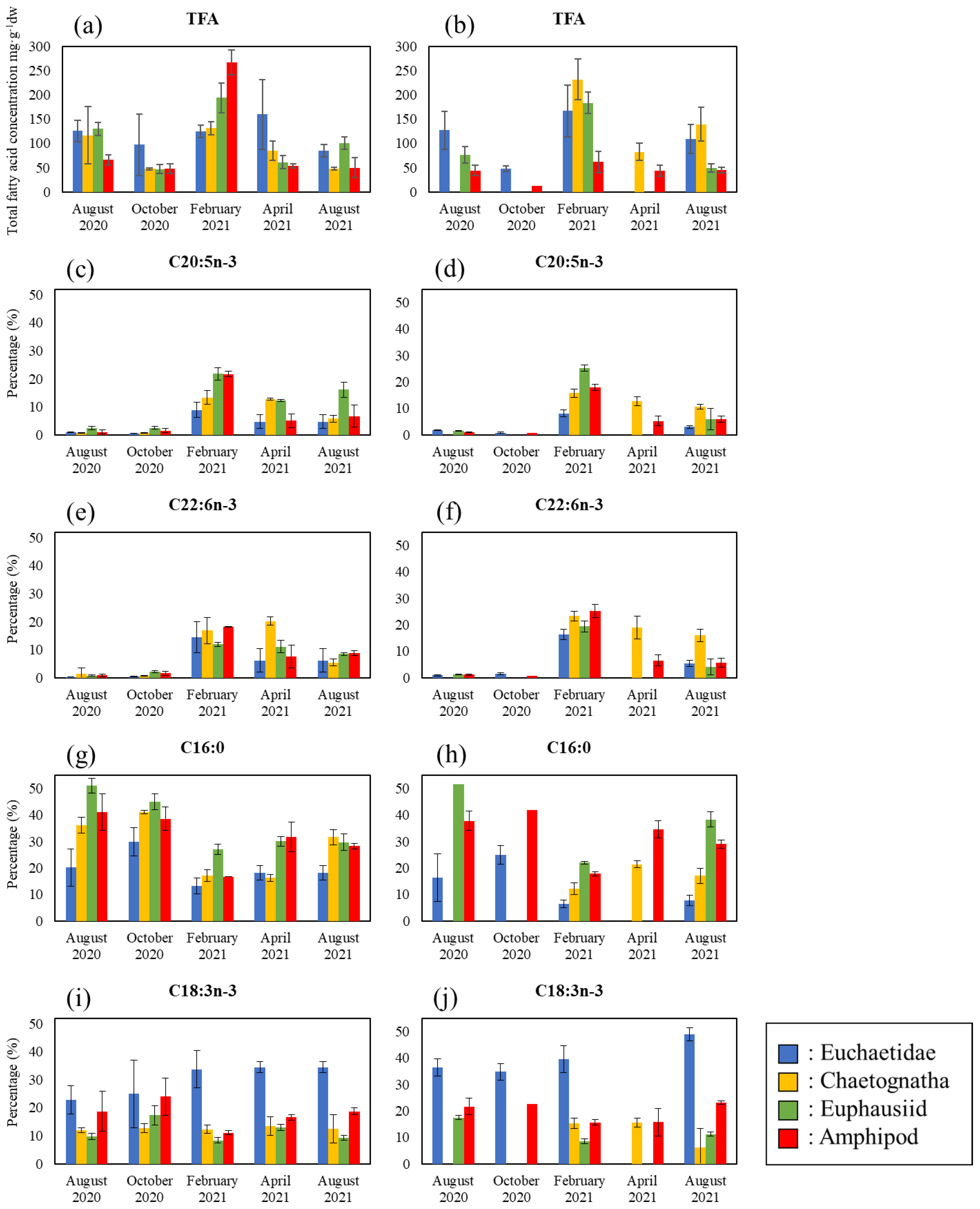
3.3. Changes in Total Fatty Acid Concentrations and the Proportion of Major Fatty Acids in Zooplankton Taxa
3.4. Estimating Changes in the Ecological Niche of Zooplankton
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M. The key role of zooplankton in ecosystem services: A perspective of interaction between zooplankton and fish recruitment. Ecol. Indic. 2021, 129, 107867. [Google Scholar] [CrossRef]
- Sterner, R. Role of zooplankton in aquatic ecosystems. In Encyclopedia of Inland Waters; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 678–688. [Google Scholar] [CrossRef]
- Winder, M.; Jassby, A.D. Shifts in Zooplankton Community Structure: Implications for Food Web Processes in the Upper San Francisco Estuary. Estuaries Coasts 2011, 34, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Ha, S.-Y.; Lee, S.; Kim, J.-H.; Shin, K.-H. Trophic dynamics of zooplankton before and after polar night in the Kongsfjorden (Svalbard): Evidence of trophic position estimated by δ15N analysis of amino acids. Front. Mar. Sci. 2020, 7, 489. [Google Scholar] [CrossRef]
- Stevens, C.; Sahota, R.; Galbraith, M.; Venello, T.; Bazinet, A.; Hennekes, M.; Yongblah, K.; Juniper, S. Total lipid and fatty acid composition of mesozooplankton functional group members in the NE Pacific over a range of productivity regimes. Mar. Ecol. Prog. Ser. 2022, 687, 43–64. [Google Scholar] [CrossRef]
- Athira, T.R.; Nefla, A.; Shifa, C.T.; Shamna, H.; Aarif, K.M.; Almaarofi, S.S.; Rashiba, A.P.; Reshi, O.R.; Jobiraj, T.; Thejass, P.; et al. The impact of long-term environmental change on zooplankton along the southwestern coast of India. Environ. Monit. Assess. 2022, 194, 316. [Google Scholar] [CrossRef]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Chen, H.; Govaert, L.; Wang, W.; Yang, J. Responses of zooplankton body size and community trophic structure to temperature change in a subtropical reservoir. Ecol. Evol. 2019, 9, 12544–12555. [Google Scholar] [CrossRef]
- Joo, H.; Son, S.; Park, J.-W.; Kang, J.J.; Jeong, J.-Y.; Kwon, J.-I.; Kang, C.-K.; Lee, S.H. Small phytoplankton contribution to the total primary production in the highly productive Ulleung Basin in the East/Japan Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 54–61. [Google Scholar] [CrossRef]
- Kang, J.J.; Jang, H.K.; Lim, J.-H.; Lee, D.; Lee, J.H.; Bae, H.; Lee, C.H.; Kang, C.-K.; Lee, S.H. Characteristics of Different Size Phytoplankton for Primary Production and Biochemical Compositions in the Western East/Japan Sea. Front. Microbiol. 2020, 11, 560102. [Google Scholar] [CrossRef]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5–16. [Google Scholar] [CrossRef]
- Han, I.-S.; Lee, J.-S. Change the annual amplitude of sea surface temperature due to climate change in a recent decade around the Korean Peninsula. J. Korean Soc. Mar. Environ. Saf. 2020, 26, 233–241. [Google Scholar] [CrossRef]
- Chang, K.-I.; Zhang, C.-I.; Park, C.; Kang, D.-J.; Ju, S.-J.; Lee, S.-H.; Wimbush, M. (Eds.) Oceanography of the East Sea (Japan Sea), 1st ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Marañón, E.; Cermeño, P.; Latasa, M.; Tadonléké, R.D. Temperature, resources, and phytoplankton size structure in the ocean. Limnol. Oceanogr. 2012, 57, 1266–1278. [Google Scholar] [CrossRef]
- Ashjian, C.J.; Davis, C.S.; Gallager, S.M.; Alatalo, P. Characterization of the zooplankton community, size composition, and distribution in relation to hydrography in the Japan/East Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 1363–1392. [Google Scholar] [CrossRef]
- Lee, B.R.; Park, W.; Kang, H.K.; Lee, H.W.; Ji, H.S.; Choi, J.H. Comparison of zooplankton communities in the East Sea, East China Sea and Philippine Sea. J. Environ. Biol. 2019, 40, 861–870. [Google Scholar] [CrossRef]
- Lee, H.; Choi, J.; Im, Y.; Oh, W.; Hwang, K.; Lee, K. Spatial–Temporal Distribution of the Euphausiid Euphausia pacifica and Fish Schools in the Coastal Southwestern East Sea. Water 2022, 14, 203. [Google Scholar] [CrossRef]
- Henschke, N.; Everett, J.D.; Suthers, I.M.; Smith, J.A.; Hunt, B.P.V.; Doblin, M.A.; Taylor, M.D. Zooplankton trophic niches respond to different water types of the western Tasman Sea: A stable isotope analysis. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2015, 104, 1–8. [Google Scholar] [CrossRef]
- Kozak, E.R.; Franco-Gordo, C.; Godínez-Domínguez, E.; Suárez-Morales, E.; Ambriz-Arreola, I. Seasonal variability of stable isotope values and niche size in tropical calanoid copepods and zooplankton size fractions. Mar. Biol. 2020, 167, 37. [Google Scholar] [CrossRef]
- Schoo, K.L.; Boersma, M.; Malzahn, A.M.; Löder, M.G.J.; Wiltshire, K.H.; Aberle, N. Dietary and seasonal variability in trophic relations at the base of the North Sea pelagic food web revealed by stable isotope and fatty acid analysis. J. Sea Res. 2018, 141, 61–70. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Variation in δ15N and δ13C trophic fractionation: Implications for aquatic food web studies. Limnol. Oceanogr. 2001, 46, 2061–2066. [Google Scholar] [CrossRef]
- Ahn, I.-Y.; Elias-Piera, F.; Ha, S.-Y.; Rossi, S.; Kim, D.-U. Seasonal Dietary Shifts of the Gammarid Amphipod Gondogeneia antarctica in a Rapidly Warming Fjord of the West Antarctic Peninsula. J. Mar. Sci. Eng. 2021, 9, 1447. [Google Scholar] [CrossRef]
- Ke, Z.; Li, R.; Chen, D.; Zhao, C.; Tan, Y. Spatial and seasonal variations in the stable isotope values and trophic positions of dominant zooplankton groups in Jiaozhou Bay, China. Front. Mar. Sci. 2022, 9, 968. [Google Scholar] [CrossRef]
- El-Sabaawi, R.; Dower, J.F.; Kainz, M.; Mazumder, A. Characterizing dietary variability and trophic positions of coastal calanoid copepods: Insight from stable isotopes and fatty acids. Mar. Biol. 2009, 156, 225–237. [Google Scholar] [CrossRef]
- Choi, H.; Won, H.; Kim, J.H.; Yang, E.J.; Cho, K.H.; Lee, Y.; Kang, S.H.; Shin, K.H. Trophic Dynamics of Calanus hyperboreus in the Pacific Arctic Ocean. J. Geophys. Res. Ocean. 2021, 126, e2020JC017063. [Google Scholar] [CrossRef]
- Hiltunen, M.; Strandberg, U.; Brett, M.T.; Winans, A.K.; Beauchamp, D.A.; Kotila, M.; Keister, J.E. Taxonomic, Temporal, and Spatial Variations in Zooplankton Fatty Acid Composition in Puget Sound, WA, USA. Estuaries Coasts 2022, 45, 567–581. [Google Scholar] [CrossRef]
- Parson, T.R.; Maita, Y.; Lalli, C.M. A Manual of Biological and Chemical Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST-palaeontological statistics, ver. 1.89. Palaeontol. Electron 2001, 4, 1–9. [Google Scholar]
- Kim, H.-C.; Yoo, S.; Oh, I.S. Relationship between phytoplankton bloom and wind stress in the sub-polar frontal area of the Japan/East Sea. J. Mar. Syst. 2007, 67, 205–216. [Google Scholar] [CrossRef]
- Lee, M.; Ro, H.; Kim, Y.-B.; Park, C.-H.; Baek, S.-H. Relationship of Spatial Phytoplankton Variability during Spring with Eutrophic Inshore and Oligotrophic Offshore Waters in the East Sea, Including Dokdo, Korea. J. Mar. Sci. Eng. 2021, 9, 1455. [Google Scholar] [CrossRef]
- Kürten, B.; Al-Aidaroos, A.M.; Kürten, S.; El-Sherbiny, M.M.; Devassy, R.P.; Struck, U.; Zarokanellos, N.; Jones, B.H.; Hansen, T.; Bruss, G. Carbon and nitrogen stable isotope ratios of pelagic zooplankton elucidate ecohydrographic features in the oligotrophic Red Sea. Prog. Oceanogr. 2016, 140, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Domingues, R.B.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Ammonium, nitrate and phytoplankton interactions in a freshwater tidal estuarine zone: Potential effects of cultural eutrophication. Aquat. Sci. 2011, 73, 331–343. [Google Scholar] [CrossRef]
- Tunēns, J.; Aigars, J.; Poikāne, R.; Jurgensone, I.; Labucis, A.; Labuce, A.; Liepiņa-Leimane, I.; Buša, L.; Vīksna, A. Stable Carbon and Nitrogen Isotope Composition in Suspended Particulate Matter Reflects Seasonal Dynamics of Phytoplankton Assemblages in the Gulf of Riga, Baltic Sea. Estuaries Coasts 2022, 45, 2112–2123. [Google Scholar] [CrossRef]
- Wu, Z.; Yu, Z.; Song, X.; Wang, W.; Zhou, P.; Cao, X.; Yuan, Y. Key nitrogen biogeochemical processes in the South Yellow Sea revealed by dual stable isotopes of nitrate. Estuar. Coast. Shelf Sci. 2019, 225, 106222. [Google Scholar] [CrossRef]
- Bode, A.; Lamas, A.F.; Mompeán, C. Effects of Upwelling Intensity on Nitrogen and Carbon Fluxes through the Planktonic Food Web off A Coruña (Galicia, NW Spain) Assessed with Stable Isotopes. Diversity 2020, 12, 121. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.; Wainright, S.C. Diatom sources of 13 C-rich carbon in marine food webs. Mar. Ecol. Prog. Ser. 1991, 76, 149–157. [Google Scholar] [CrossRef]
- Bardhan, P.; Karapurkar, S.G.; Shenoy, D.M.; Kurian, S.; Sarkar, A.; Maya, M.V.; Naik, H.; Varik, S.; Naqvi, S.W.A. Carbon and nitrogen isotopic composition of suspended particulate organic matter in Zuari Estuary, west coast of India. J. Mar. Syst. 2015, 141, 90–97. [Google Scholar] [CrossRef]
- Perry, R.I.; Thompson, P.A.; Mackas, D.L.; Harrison, P.J.; Yelland, D.R. Stable carbon isotopes as pelagic food web tracers in adjacent shelf and slope regions off British Columbia, Canada. Can. J. Fish. Aquat. Sci. 1999, 56, 2477–2486. [Google Scholar] [CrossRef]
- Rau, G.H.; Riebesell, U.; Wolf-Gladrow, D. A model of photosynthetic 13C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake. Mar. Ecol. Prog. Ser. 1996, 133, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Silori, S.; Sharma, D.; Chowdhury, M.; Biswas, H.; Bandyopadhyay, D.; Shaik, A.U.R.; Cardinal, D.; Mandeng-Yogo, M.; Narvekar, J. Contrasting phytoplankton and biogeochemical functioning in the eastern Arabian Sea shelf waters recorded by carbon isotopes (SW monsoon). Mar. Chem. 2021, 232, 103962. [Google Scholar] [CrossRef]
- Ying, R.; Cao, Y.; Yin, F.; Guo, J.; Huang, J.; Wang, Y.; Zheng, L.; Wang, J.; Liang, H.; Li, Z.; et al. Trophic structure and functional diversity reveal pelagic-benthic coupling dynamic in the coastal ecosystem of Daya Bay, China. Ecol. Indic. 2020, 113, 106241. [Google Scholar] [CrossRef]
- Budge, S.M.; Parrish, C.C. Lipid biogeochemistry of plankton, settling matter and sediments in Trinity Bay, Newfoundland. II. Fatty acids. Org. Geochem. 1998, 29, 1547–1559. [Google Scholar] [CrossRef]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Muller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine food environment. Adv. Mar. Biol. 2003, 46, 226–340. [Google Scholar] [CrossRef]
- El-Sabaawi, R.W.; Sastri, A.R.; Dower, J.F.; Mazumder, A. Deciphering the seasonal cycle of copepod trophic dynamics in the Strait of Georgia, Canada, using stable isotopes and fatty acids. Estuaries Coasts 2010, 33, 738–752. [Google Scholar] [CrossRef]
- Lee, R.F.; Hagen, W.; Kattner, G. Lipid storage in marine zooplankton. Mar. Ecol. Prog. Ser. 2006, 307, 273–306. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Lannig, G. Interactive effects of metal pollution and temperature on metabolism in aquatic ectotherms: Implications of global climate change. Clim. Res. 2008, 37, 181–201. [Google Scholar] [CrossRef] [Green Version]
- Werbrouck, E.; Van Gansbeke, D.; Vanreusel, A.; De Troch, M. Temperature affects the use of storage fatty acids as energy source in a benthic copepod (Platychelipus littoralis, Harpacticoida). PLoS ONE 2016, 11, e0151779. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Gonzàlez, G.; Agustí, S. Long-term exposure to increasing temperature can offset predicted losses in marine food quality (fatty acids) caused by ocean warming. Evol. Appl. 2020, 13, 2497–2506. [Google Scholar] [CrossRef]
- Shin, K.-H.; Hama, T.; Yoshie, N.; Noriki, S.; Tsunogai, S. Dynamics of fatty acids in newly biosynthesized phytoplankton cells and seston during a spring bloom off the west coast of Hokkaido Island, Japan. Mar. Chem. 2000, 70, 243–256. [Google Scholar] [CrossRef]
- Shin, K.-H.; Hama, T.; Handa, N. Effect of nutrient conditions on the composition of photosynthetic products in the East China Sea and surrounding waters. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2003, 50, 389–401. [Google Scholar] [CrossRef]
- Landry, M.R. Switching between herbivory and carnivory by the planktonic marine copepod Calanus pacificus. Mar. Biol. 1981, 65, 77–82. [Google Scholar] [CrossRef]
- López-Ibarra, G.A.; Bode, A.; Hernández-Trujillo, S.; Zetina-Rejón, M.J.; Arreguín-Sánchez, F. Trophic position of twelve dominant pelagic copepods in the eastern tropical Pacific Ocean. J. Mar. Syst. 2018, 187, 13–22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Station | Latitude | Longitude | Sample Collection Depth (m) | Bottom Depth (m) | Species |
|---|---|---|---|---|---|---|
| East Sea | 105-05 | 37.55 | 129.38 | 100 | 280 | Euchaetidae |
| Chaetognatha | ||||||
| Euphausiid | ||||||
| Amphipod | ||||||
| 105-11 | 37.55 | 131.24 | 100 | 1140 | Euchaetidae | |
| Chaetognatha | ||||||
| Euphausiid | ||||||
| Amphipod |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Yun, H.-Y.; Won, E.-J.; Choi, H.; Youn, S.-H.; Shin, K.-H. Influences of Seasonal Variability and Potential Diets on Stable Isotopes and Fatty Acid Compositions in Dominant Zooplankton in the East Sea, Korea. J. Mar. Sci. Eng. 2022, 10, 1768. https://doi.org/10.3390/jmse10111768
Kim J, Yun H-Y, Won E-J, Choi H, Youn S-H, Shin K-H. Influences of Seasonal Variability and Potential Diets on Stable Isotopes and Fatty Acid Compositions in Dominant Zooplankton in the East Sea, Korea. Journal of Marine Science and Engineering. 2022; 10(11):1768. https://doi.org/10.3390/jmse10111768
Chicago/Turabian StyleKim, Jieun, Hee-Young Yun, Eun-Ji Won, Hyuntae Choi, Seok-Hyeon Youn, and Kyung-Hoon Shin. 2022. "Influences of Seasonal Variability and Potential Diets on Stable Isotopes and Fatty Acid Compositions in Dominant Zooplankton in the East Sea, Korea" Journal of Marine Science and Engineering 10, no. 11: 1768. https://doi.org/10.3390/jmse10111768