
Seasonal and Spatial Variations in Fish Assemblage in the Yangtze Estuary and Adjacent Waters and Their Relationship with Environmental Factors



Abstract
:1. Introduction
2. Materials and Methods
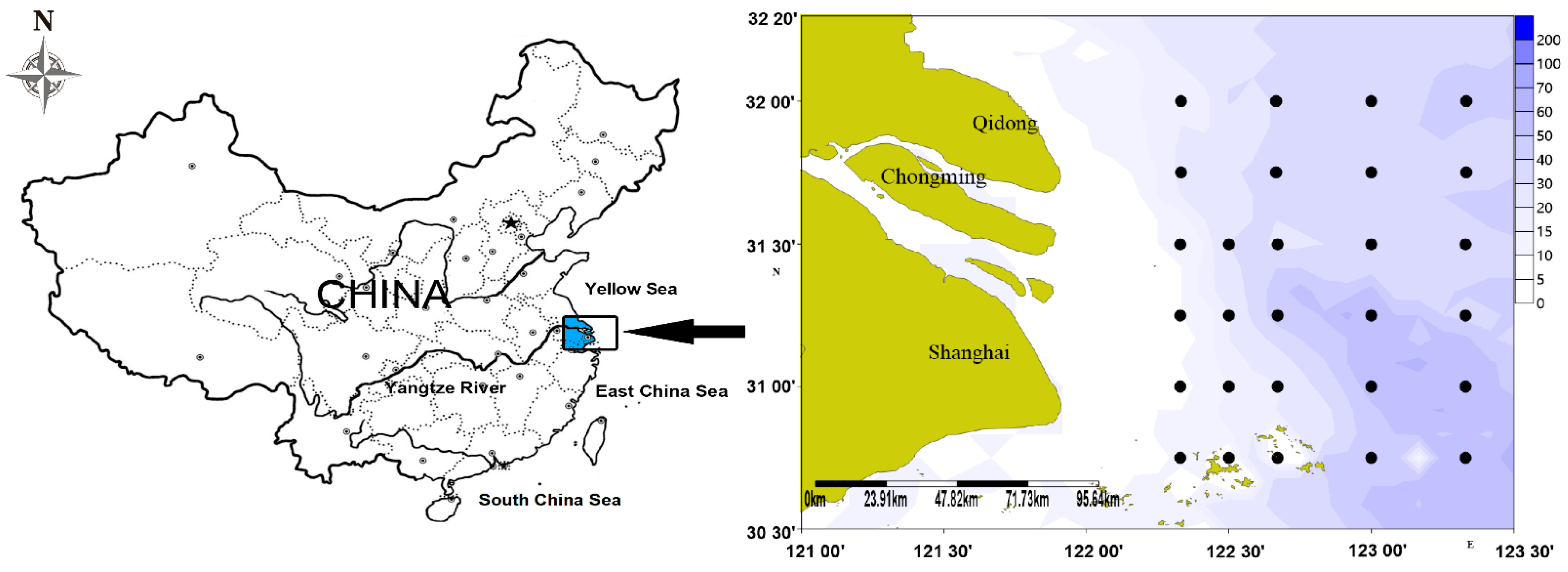
2.1. Study Area
2.2. Sampling
2.3. Data Analysis
3. Results
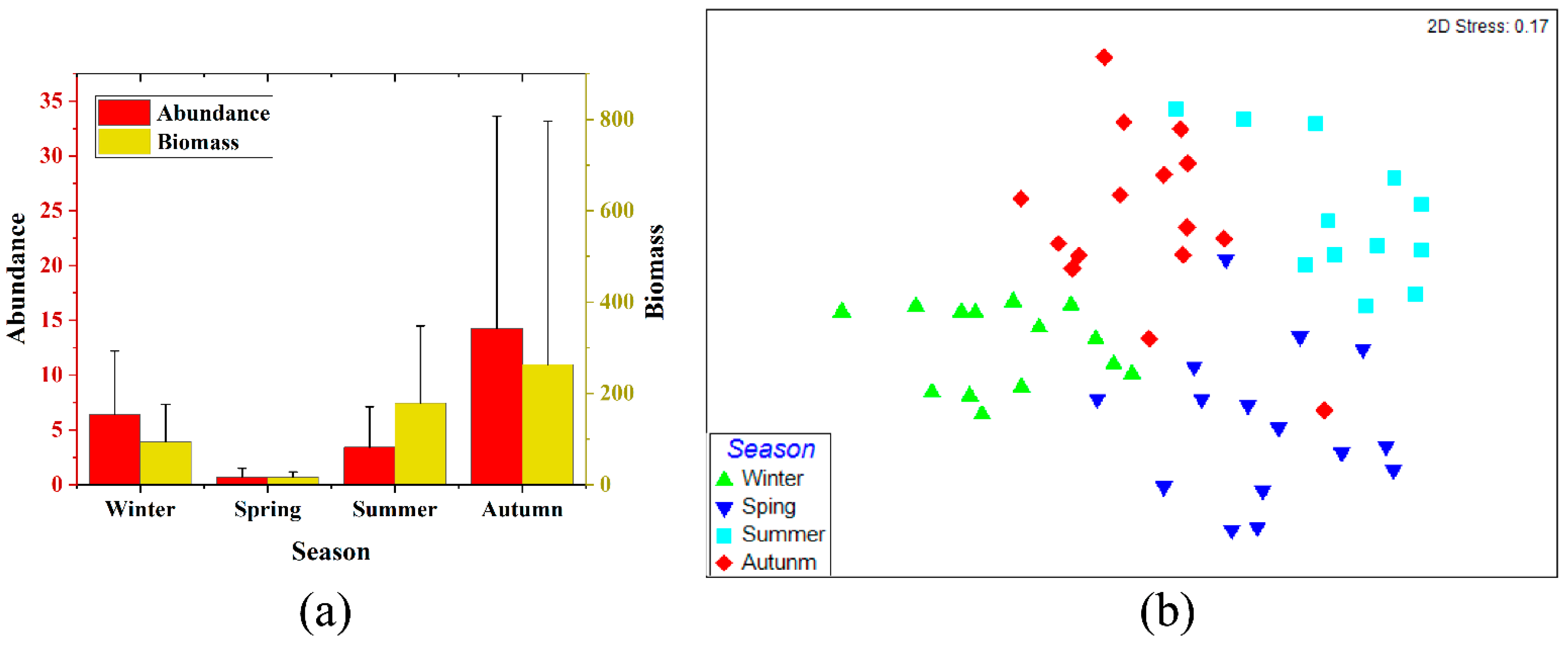
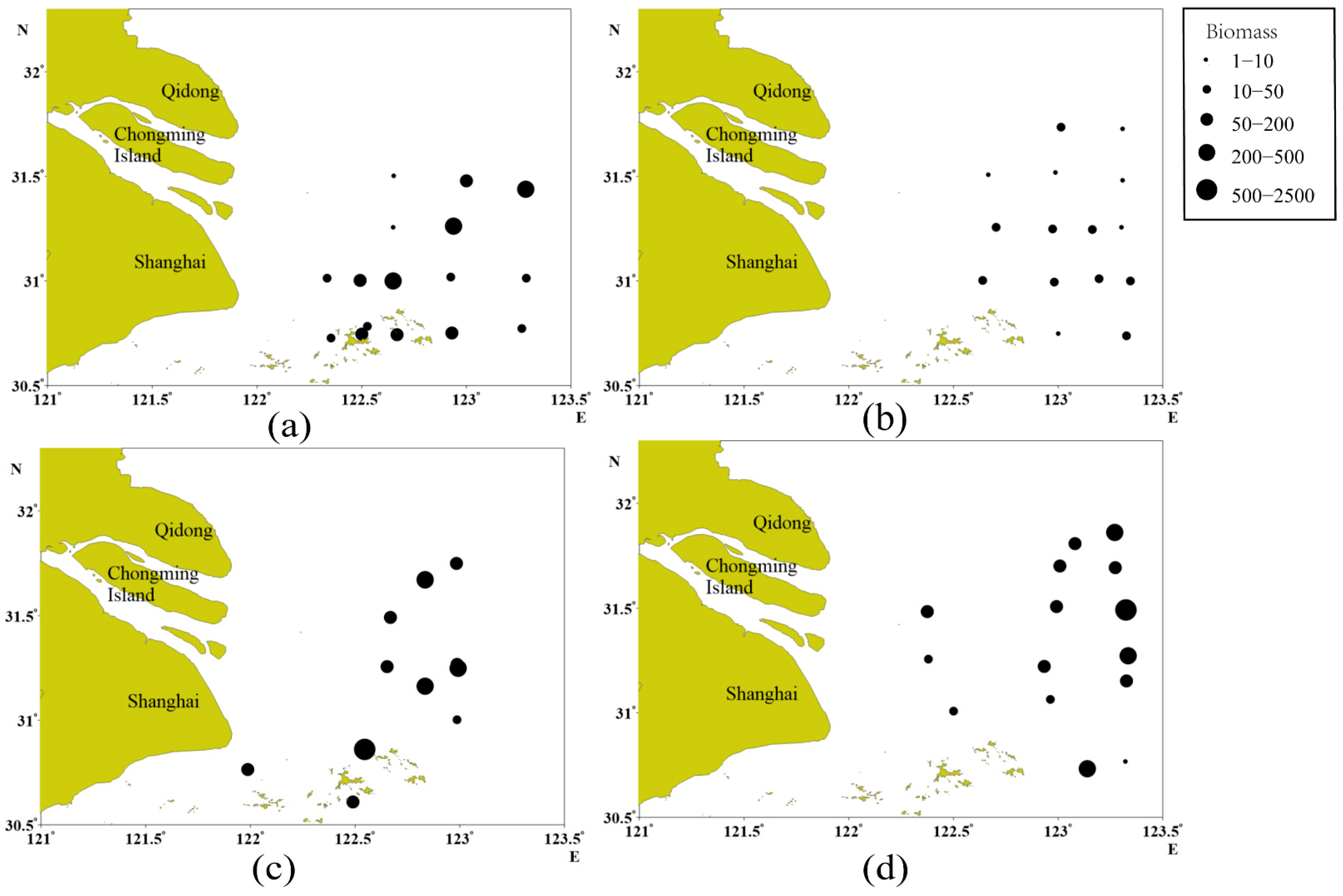
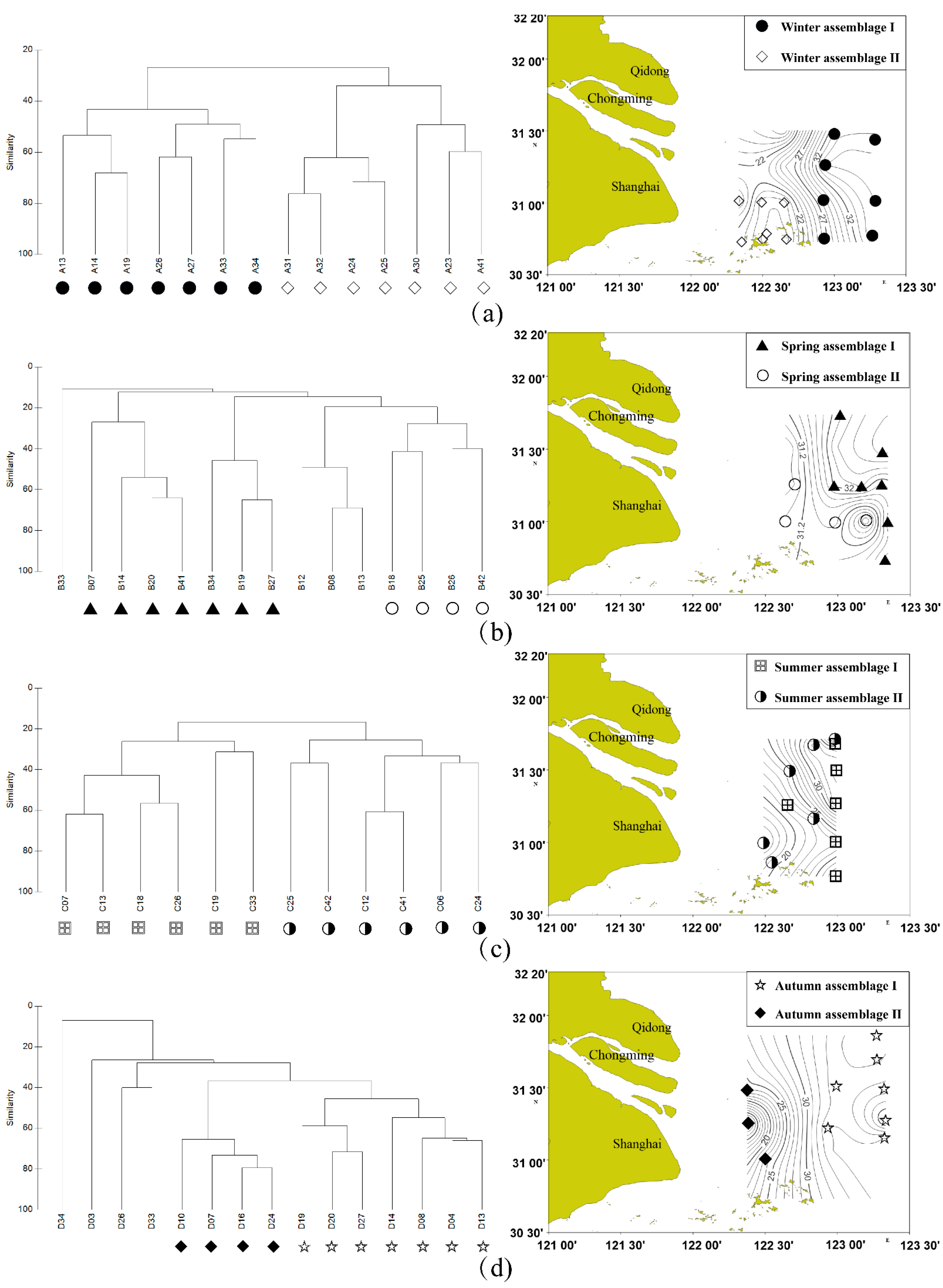
3.1. Seasonal and Spatial Variations in Fish Assemblages
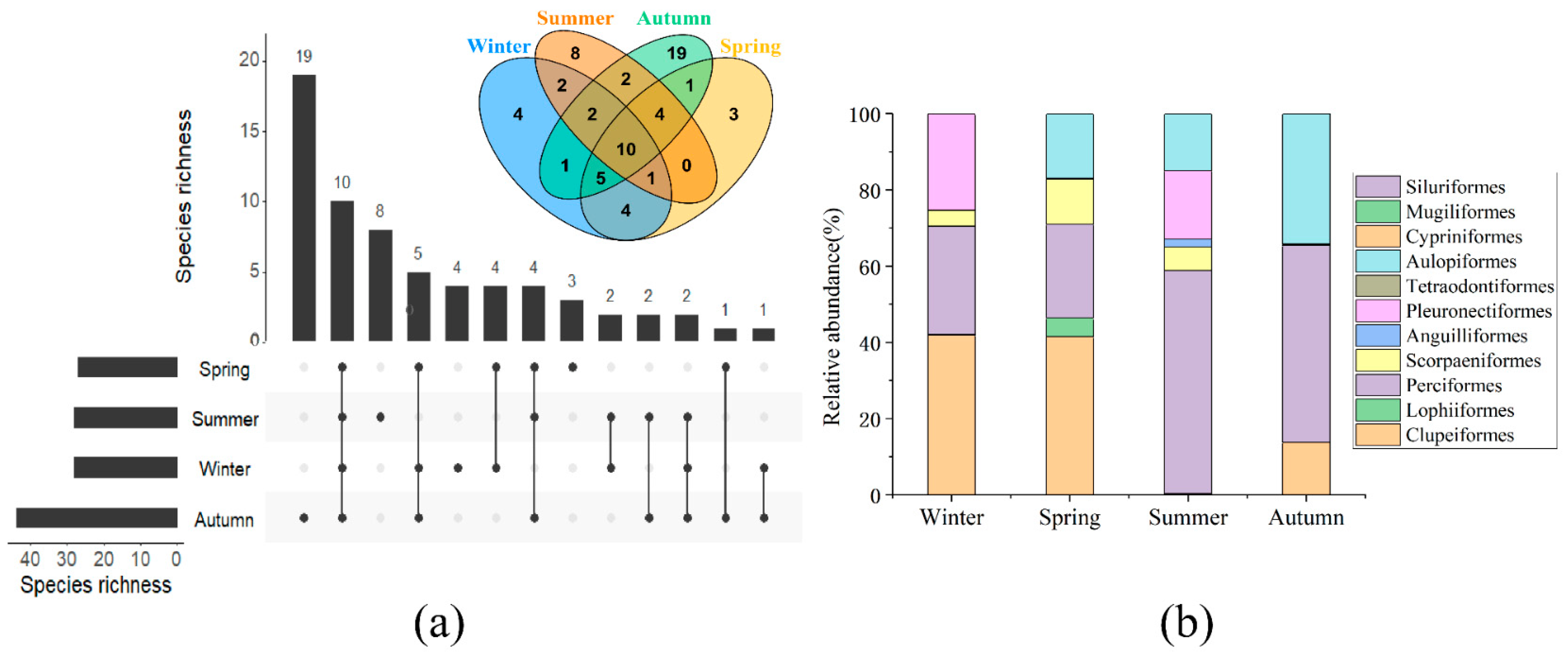
3.2. Species Composition
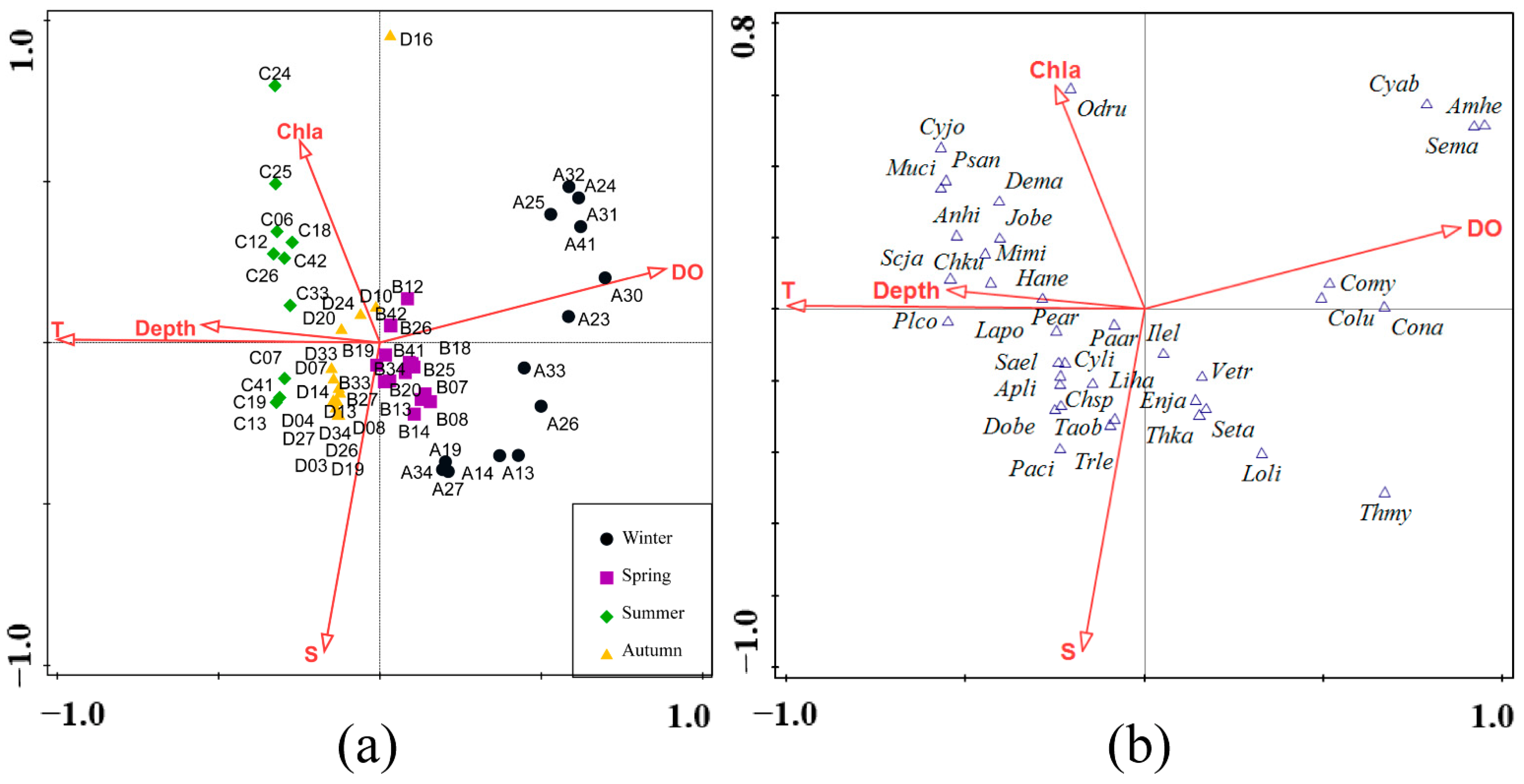
3.3. Fish Assemblage in Relation to Environmental Factors
4. Discussion
4.1. Seasonal and Spatial Variations in Fish Assemblages
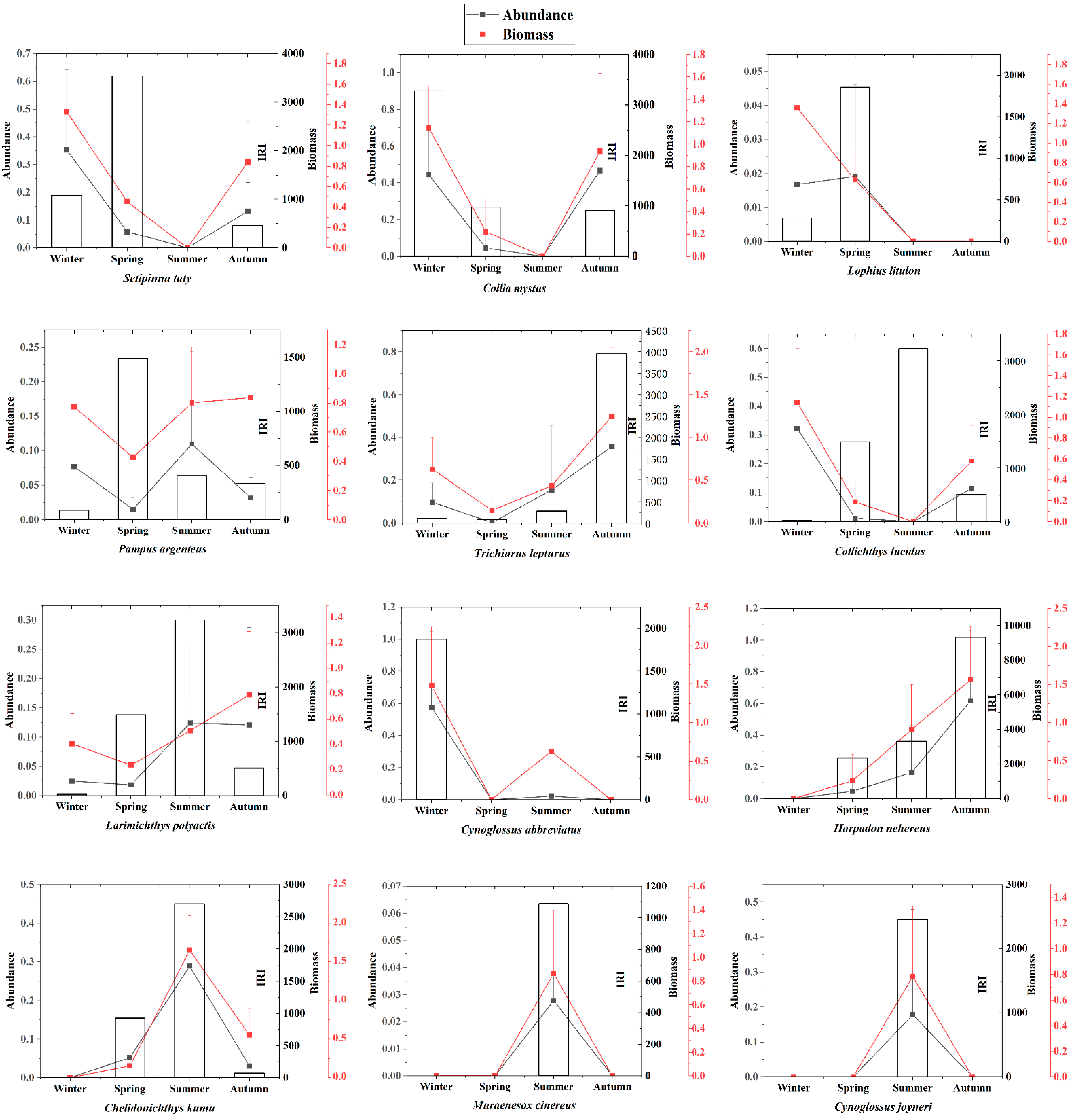
4.2. Seasonal Migration and Spatial Distribution of Dominant Species
4.3. Influence of Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, B.Z.; Shen, H.T. Three Gorges Project and Estuary Ecological Environment; Science Press: Beijing, China, 1994. [Google Scholar]
- Yang, D.Z.; Yin, B.S.; Liu, Z.L.; Bai, T.; Qi, J.F.; Chen, H.Y. Numerical study on the pattern and origins of Kuroshio branches in the bottom water of southern East China Sea in summer. J. Geophys. Res. Ocean. 2012, 117, C02014. [Google Scholar] [CrossRef]
- Xu, Y.; Ma, L.; Li, X.Z.; Sun, Y.; Gong, L. Demersal fish assemblage characteristics and their relationship with environmental variables in the sea off Changjiang river estuary. Oceanol. Limnol. Sin. 2017, 48, 1383–1391. [Google Scholar]
- Xu, Q.H.; Qian, J.H.; Chen, Z.Y.; Zhang, H.P.; Chen, L. Water Environment Status and Pollution Prevention and Control in Yangtze River Estuary and Adjacent Waters. China Dev. 2015, 15, 10–14. [Google Scholar]
- Song, Z.K.; Cheng, H.Q.; Hu, H.; Li, J.F.; Jiang, Y.P. Numerical simulation analysis of influence of reclamation of north branch in the Yangtze River estuary on hydrodynamic characteristics. Yangtze River 2012, 43, 59–63. [Google Scholar]
- Zhuang, P. Fishes of the Yangtze Estuary; China Agriculture Press: Beijing, China, 2018. [Google Scholar]
- Wang, X.C.; Xie, P.F.; Li, Q.; Zhang, J.Y.; Li, H.J. Ecological Environment of the Yangtze Estuary and Protection Countermeasures. Res. Environ. Sci. 2020, 33, 1197–1205. [Google Scholar]
- Xu, Z.L.; Yuan, Q.; Jiang, M.; Zang, Z.J. An investigation of fish eggs, larvae and juveniles in the Changjiang esturary. J. Fish. Sci. China 1999, 6, 63–64. [Google Scholar]
- Zhang, G.X.; Zhang, X.S. A survey on the fisheries of set stow net in the Chang Jiang Estuary. J. Fish. China 1985, 9, 185–198, (In Chinese with English abstract). [Google Scholar]
- Zhang, H.; He, W.S.; Tong, C.F.; Lu, J.J. Species composition and biodiversity of the intertidal fishes in Chongxi Wetland in winter in Changjiang Estuary. Resour. Environ. Yangtze Basin 2007, 16, 308–313. [Google Scholar]
- Yu, H.C.; Xian, W.W. Fish Assemblage Structure in Adjacent Sea of Yangtze Estuary and Its Relationshp with Envirorm ental Factors. J. Yangtze River Sci. Res. Inst. 2010, 27, 88–92. [Google Scholar]
- Shi, Y.R.; Chao, M.; Quan, W.M.; Huang, H.J.; Shen, X.Q. Fish community diversity analyses in the Yangtze Estuary, China. J. Fish. Sci. China 2012, 19, 1051–1059. [Google Scholar]
- Zhang, Y.Q.; Xian, W.W.; Li, W.L. Fish Assemblage Structure in Adjacent Sea of Changj iang Estuary in Spring of 2004 and 2007 and Its Association With Environmental Factors. Pericdical Ocean Univ. China 2013, 43, 67–74. [Google Scholar]
- Cheila, A.; Rui, C.; Marco, S.; Luís, B.; Pedro, M.; Joaquim, R.; Karim, E.; Jorge, M.S.G. Use of different intertidal habitats by faunal communities in a temperate coastal lagoon. Estuar. Coast. Shelf Sci. 2008, 80, 357–364. [Google Scholar]
- Ren, Q.Q.; Xian, W.W.; Zhang, Y.; Liu, C.L.; Li, W.L. Invertebrate assemblage structure associated with key environmental factors in the Yangtze Estuary, China. Chin. J. Appl. Ecol. 2018, 29, 3067–3077. [Google Scholar]
- Chu, C.; Jones, N.E. Spatial variability of thermal regimes and other environmental determinants of stream fish communities in the Great Lakes Basin, Ontario, Canada. River Res. Appl. 2011, 27, 646–662. [Google Scholar] [CrossRef]
- Sun, P.F.; Shan, X.J.; Wu, Q.; Chen, Y.L.; Jin, X.S. Seasonal variations in fish community structure in the Laizhou Bay and the Yellow River Estuary. Acta Ecol. Sin. 2014, 34, 367–376. [Google Scholar]
- Whitfield, A.K. Fish species diversity in Southern African estuarine systems: An evolutionary perspective. Environ. Biol. Fishes 1994, 40, 37–48. [Google Scholar] [CrossRef]
- Stoner, A.W. Community structure of the demersal fish species of Laguna Joyuda, Puerto Rico. Estuaries 1986, 9, 142–152. [Google Scholar] [CrossRef]
- Flores-Verdugo, F.; González-Farías, F.; Ramírez-Flores, O.; AmezcuaLinares, F.; Yánez-Arancibia, A.; Alvarez-Rubio, M.; Day, J.W. Mangrove ecology, aquatic primary productivity, and fish community dynamics in the Teacapán-Agua Brava lagoonestuarine system (Mexican Pacific). Estuaries 1990, 13, 219–230. [Google Scholar] [CrossRef]
- Fraser, T.H. Abundance, seasonality, community indices, trends and relationships with physicochemical factors of trawled fish in upper Charlotte Harbor, Florida. Bull. Mar. Sci. 1997, 60, 739–763. [Google Scholar]
- Mukherjee, S.; Chaudhuri, A.; Kundu, N.; Mitra, S.; Homechaudhuri, S. Comprehensive Analysis of Fish Assemblages in Relation to Seasonal Environmental Variables in an Estuarine River of Indian Sundarbans. Estuaries Coasts 2013, 36, 192–202. [Google Scholar] [CrossRef]
- Beaudreau, A.H.; Bergstrom, C.A.; Whitney, E.J.; Duncan, D.H.; Lundstrom, N.C. Seasonal and interannual variation in high-latitude estuarine fish community structure along a glacial to non-glacial watershed gradient in Southeast Alaska. Environ. Biol. Fishes 2022, 105, 431–452. [Google Scholar] [CrossRef]
- Yu, H.C. Fish Community Structure in the Changjiang Estuary and Adjacent Waters; Institute of Oceanology, Chinese Academy of Sciences: Qingdao, China, 2008. [Google Scholar]
- Sheaves, M.; Johnston, R. Ecological drivers of spatial variability among fish fauna of 21 tropical Australian estuaries. Mar. Ecol. Prog. Ser. 2009, 385, 245–260. [Google Scholar] [CrossRef]
- Maes, J.; Stevens, M.; Ollevier, F. The composition and community structure of the ichthyofauna of the upper Scheldt estuary: Synthesis of a 10-year data collection (1991–2001). J. Appl. Ichthyol. 2005, 21, 86–93. [Google Scholar] [CrossRef]
- Whitfield, A.K. Biology and Ecology of Fishes in Southern African Estuaries, 2nd, ed.; Ichthyological Monogro; Smith Institute of Ichthyology: Grahamstown, South Africa, 1998. [Google Scholar]
- Zhuang, P. Fishes of Yangtze Estuary; Shanghai Scientific & Technical Publishers: Shanghai, China, 2006. [Google Scholar]
- Wu, J.H.; Wang, J.Q.; Dai, X.J.; Tian, S.Q.; Liu, J.; Chen, J.H.; Wang, X.F. An analysis of spatial co-occurrence pattern of fish species of Yangtze Estuary based on probabilistic model. South China Fish. Sci. 2019, 15, 1–9. [Google Scholar]
- Shi, Y.R. Studies on Fish Community Diversity and Community Dynamics Based on Multivariate Analysis in the Yangtze Estuary; Shanghai Ocean University: Shanghai, China, 2012. [Google Scholar]
- Luo, H.Z.; Zhang, H.D.; Li, P.F.; Zhou, Y.D. Analysis of the Current Situation of Fishery Biology of Harpodon nehereus in the East China Sea. J. Zhejiang Ocean Univ. Nat. Sci. 2012, 31, 202–205+233. [Google Scholar]
- Laffaille, P.; Feunteun, E.; Lefeuvre, J.C. Composition of fish communities in a European macrotidal salt marsh (the Moint Saint-Michel bay, France). Estuar. Coast. Shelf Sci. 2000, 51, 429–438. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, J.H.; Li, S.F. A study on the distribution of Setipinna taty in the East China Sea. Mar. Fish. 2004, 26, 255–260. [Google Scholar]
- Shen, X.Q.; Shi, Y.R.; Chao, M.; Huang, H.J.; Tang, F.H. Fish community structure of the Yangtze Estuary in summer and autumn. J. Fish. China 2011, 35, 700–710. [Google Scholar]
- Shen, G.Y.; Shi, B.Z. Marine Ecology; Science Press: Beijing, China, 2003; pp. 153–183. [Google Scholar]
- Martino, E.J.; Able, K.W. Fish assemblages across the marine to low salinity transition zone of a temperate estuary. Estuar. Coast. Shelf Sci. 2003, 56, 969–987. [Google Scholar] [CrossRef]
- Yang, G. Fish Community Structure and Correlation to Major Environmental Factors in Yangtze Estuary; Shanghai Ocean University: Shanghai, China, 2012. [Google Scholar]
- David, J.H.; Kirk, O.W.; Jenny, S.B. Local and regional determinants of stream fish assemblage structure: Inferences based on taxonomic vs. functional groups. J. Biogeogr. 2007, 34, 324–338. [Google Scholar]
- Márcia, C.C.A.; Francisco, G.A.; André, L.M.P.; Márcio, A.S. Co-occurrence of demersal fishes in a tropical bay in southeastern Brazil: A null model analysis. Estuar. Coast. Shelf Sci. 2006, 66, 315–322. [Google Scholar]
- Remmert, H. Studies and thoughts about the zonation along the rocky shores of the Baltic. Zoologica 1983, 22, 121–125. [Google Scholar]
- Hu, Y.Y.; Zhao, L.; Guo, X.Y.; Wei, H. Variations of the water temperature offshore the Changjangriver Estuary in winters and sprincs. Oceanol. Et Limnol. Sin. 2012, 43, 655–661. [Google Scholar]
- Zhou, X.Y.; Hu, D.B.; Wang, C.Z.; Hu, X.J.; Yao, S.K. Seasonal and Interannual SST Var iations in the Changjiang Estuary. Period. Ocean. Univ. China 2005, 35, 357–362. [Google Scholar]
- Wang, H.C.; Yu, J.G. Effect of water temperature on fish. Heilongjiang Fish. 2011, 42, 30–31. [Google Scholar]
- Wang, X.L. Temporal and Spatial Variations of the Fish Community Structure in Haizhou Bay and Adjacent Waters; Ocean University of China: Qingdao, China, 2013. [Google Scholar]
- Li, Y.Z.; Chen, G.B.; Sun, D.R. Analysis of the composition of fishes in the Pearl River estuarine waters. J. Fish. China 2000, 24, 312–317. [Google Scholar]
- Peterson, M.S.; Ross, S.T. Dynamics of littoral fishes and decapods along a coastal river-estuarine gradient. Estuar. Coast. Shelf Sci. 1991, 33, 467–483. [Google Scholar] [CrossRef]
- Simier, M.; Laurent, C.; Ecoutin, J.M.; Albaret, J.J. The Gambia River estuary: A reference point for estuarine fish assemblages studies in West Africa. Estuar. Coast. Shelf Sci. 2006, 69, 615–628. [Google Scholar] [CrossRef]
- Garcia, A.M.; Vieira, J.P.; Winemiller, K.O. Effects of 1997–1998 El Nino on the dynamics of the shallow-water fish assemblage of the Patos Lagoon Estuary (Brazil). Estuar. Coast. Shelf Sci. 2003, 57, 489–500. [Google Scholar] [CrossRef]
- Whitfield, A.K. Ichthyofaunal assemblages in estuaries: A South African case study. Rev. Fish Biol. Fish. 1999, 91, 51–86. [Google Scholar]
- Ran, F.X.; Jin, W.J.; Huang, S.; Liu, C.X.; Li, Z.X.; Li, C.Z. Research progress on the effects of salinity change on fish. J. Northwest AF Univ. Nat. Sci. Ed. 2020, 48, 10–18. [Google Scholar]
- Ansari, Z.A.; Chatterji, A.; Ingole, B.S.; Sreepada, R.A.; Rivonkar, C.U.; Parulekar, A.H. Community structure and seasonal variation of an inshore demersal fish community at Goa, west coast of India. Estuar. Coast. Shelf Sci. 1995, 41, 593–610. [Google Scholar] [CrossRef]
- Claridge, P.N.; Potter, I.C.; Hardisty, M.W. Seasonal changes in movements, abundance, size composition and diversity of the fish fauna of the Sevem Estuary. J. Mar. Biol. Assoc. 1986, 66, 229–258. [Google Scholar] [CrossRef] [Green Version]
- Hyndes, G.A.; Platell, M.E.; Potter, I.C.; Lenanton, R.C.J. Does the composition of the demersal fish assemblages in temperate coastal waters change with depth and undergo consistent seasonal changes? Mar. Biol. 1999, 134, 335–352. [Google Scholar] [CrossRef]
- Wang, P.H.; Li, B. Historical Changes and Mechanism of Hypoxia in the Changjiang Estuary and Its Adjacent Waters. J. Zhejiang Ocean Univ. Nat. Sci. 2019, 38, 401–406. [Google Scholar]
- Wang, K.; Li, B.; Lu, J.J.; Wang, P.H. Temperature and salinity characteristics of summer hypoxia area in the Yangtze River Estuary adjacent sea area. J. Xiamen Univ. Nat. Sci. 2020, 59, 32–38. [Google Scholar]
- Wei, Q.S.; Yu, Z.G.; Xia, J.Y.; Ran, X.B.; Zhang, X.L. A preliminary analysis on the dynamic characteristics of the hypoxic zone adjacent to the Changjiang Estuary in summer. Acta Oceanol. Sin. 2011, 33, 100–109. [Google Scholar]
- Menge, B.A.; Olson, A.M. Role of scale and environmental factors in regulation of community structure. Trends Ecol. Evol. 1990, 5, 52–57. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factor | Winter | Spring | Summer | Autumn | |
|---|---|---|---|---|---|
| Depth (D) | M | 36.75 ± 27.16 | 41.80 ± 9.68 | 31.83 ± 10.20 | 36.40 ± 15.69 |
| Temperature (T) | °C | 7.92 ± 2.52 c | 14.41 ± 1.07 bc | 28.62 ± 0.84 a | 20.65 ± 1.38 ab |
| Salinity (S) | 24.77 ± 6.90 ab | 30.81 ± 0.86 ab | 24.69 ± 5.82 b | 29.61 ± 6.64 a | |
| Dissolved oxygen (DO) | mg/L | 10.05 ± 1.06 a | 9.03 ± 0.66 b | 7.25 ± 0.59 c | 7.35 ± 0.49 c |
| pH | FTU | 8.01 ± 0.05 b | 8.00 ± 0.26 b | 8.18 ± 0.05 a | 8.04 ± 0.06 b |
| Total nitrogen (TN) | μmg/L | 37.13 ± 15.93 a | 36.43 ± 7.24 a | 61.81 ± 49.66 a | 20.20 ± 12.70 b |
| Total phosphorus (TP) | μmg/L | 2.27 ± 1.71 a | 0.49 ± 0.18 b | 1.97 ± 2.64 ab | 0.93 ± 0.54 ab |
| Turbidity (Turb) | 55.25 ± 60.96 a | 12.72 ± 29.40 b | 3.63 ± 5.69 b | 7.73 ± 7.30 b | |
| Chlorophyll a (Chl a) | μg/L | 0.94 ± 0.62 | 1.91 ± 1.95 | 2.74 ± 2.58 | 0.81 ± 0.53 |
| Axes | 1 | 2 | 3 | 4 | Total Inertia |
|---|---|---|---|---|---|
| Eigenvalues | 0.592 | 0.217 | 0.195 | 0.095 | 3.367 |
| Species–environment correlations | 0.956 | 0.705 | 0.852 | 0.700 | |
| Cumulative percentage variance | |||||
| of species data | 17.59 | 24.02 | 29.82 | 32.65 | |
| of species–environment relation | 51.34 | 70.11 | 87.03 | 95.31 | |
| Sum of all eigenvalues | 3.367 | ||||
| Sum of all canonical eigenvalues | 1.154 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Ren, Q.; Liu, C.; Xian, W. Seasonal and Spatial Variations in Fish Assemblage in the Yangtze Estuary and Adjacent Waters and Their Relationship with Environmental Factors. J. Mar. Sci. Eng. 2022, 10, 1679. https://doi.org/10.3390/jmse10111679
Chen Z, Ren Q, Liu C, Xian W. Seasonal and Spatial Variations in Fish Assemblage in the Yangtze Estuary and Adjacent Waters and Their Relationship with Environmental Factors. Journal of Marine Science and Engineering. 2022; 10(11):1679. https://doi.org/10.3390/jmse10111679
Chicago/Turabian StyleChen, Zhaomin, Qingqiang Ren, Chunlong Liu, and Weiwei Xian. 2022. "Seasonal and Spatial Variations in Fish Assemblage in the Yangtze Estuary and Adjacent Waters and Their Relationship with Environmental Factors" Journal of Marine Science and Engineering 10, no. 11: 1679. https://doi.org/10.3390/jmse10111679