
Sharks Do Not Always Grow Slowly: Tagging Data Reveal a Different Pattern of Growth, Longevity and Maturity for Threatened Smooth-Hounds in the Central Mediterranean Sea
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
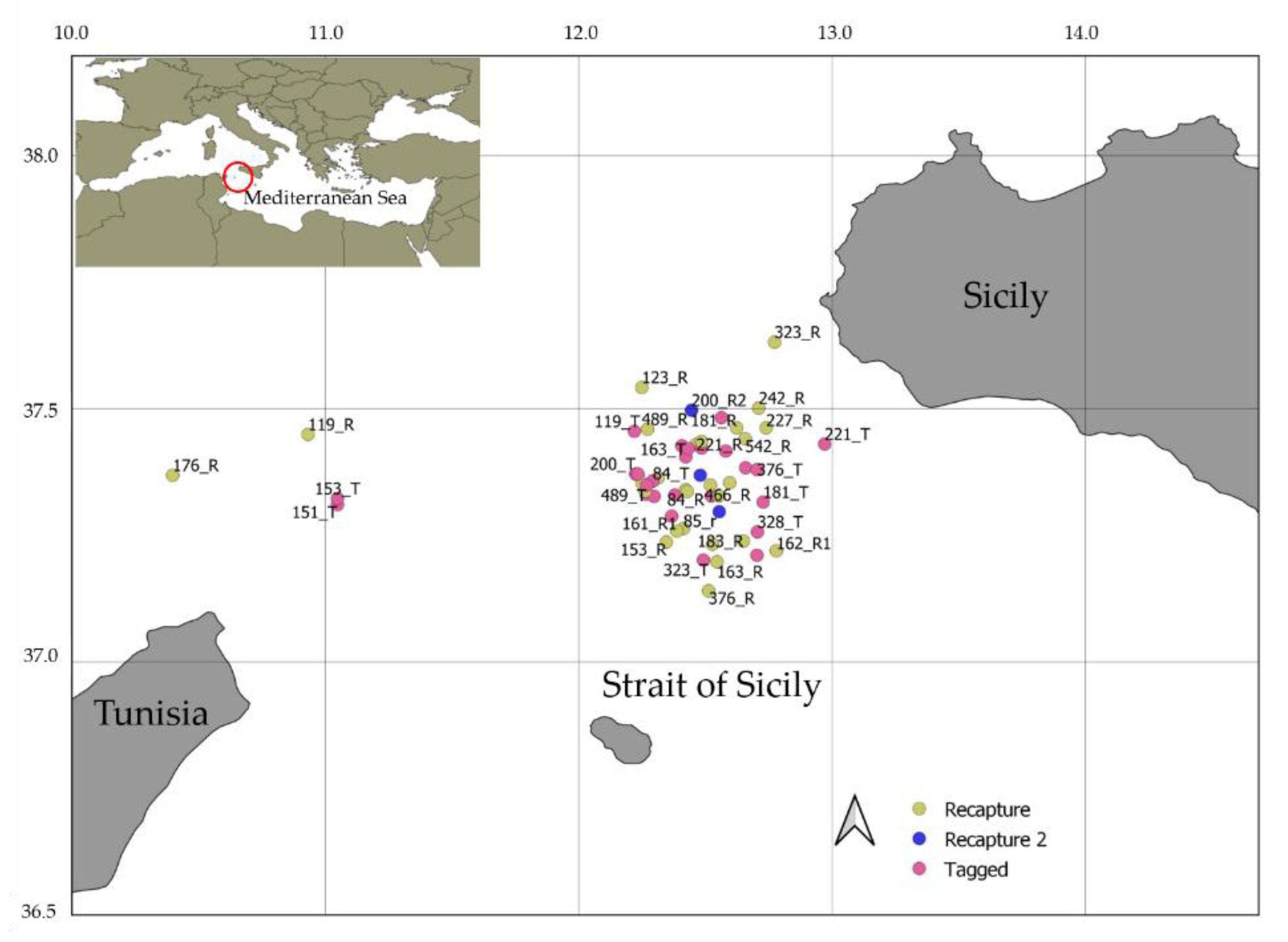
2.1. Study Area, Sample Collection, and Ethical Statement
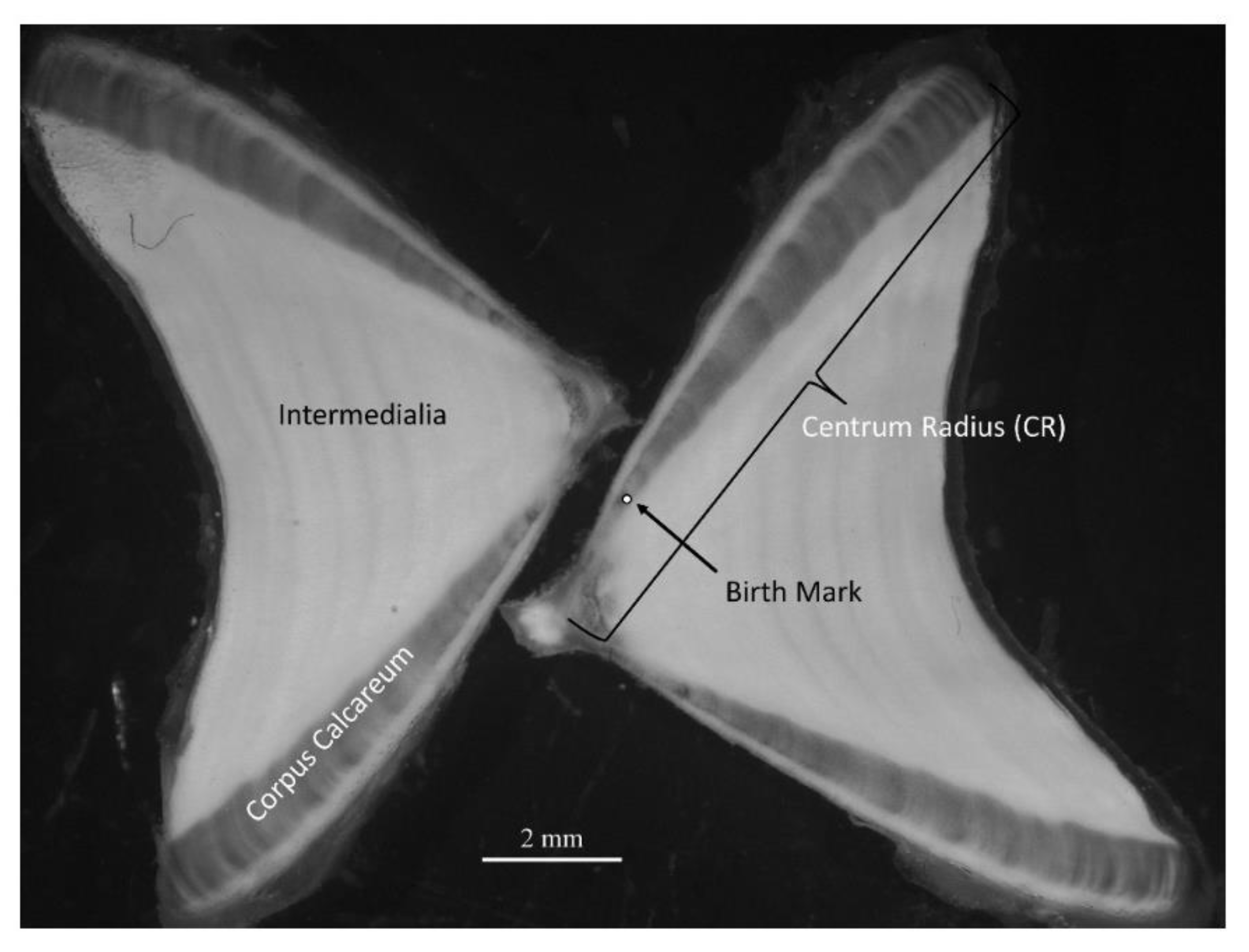
2.2. Age and Growth Curve Estimation
2.3. Validation of Ring Pattern on Vertebrae Using Tagging Data
2.4. Maturity and Reproduction
3. Results
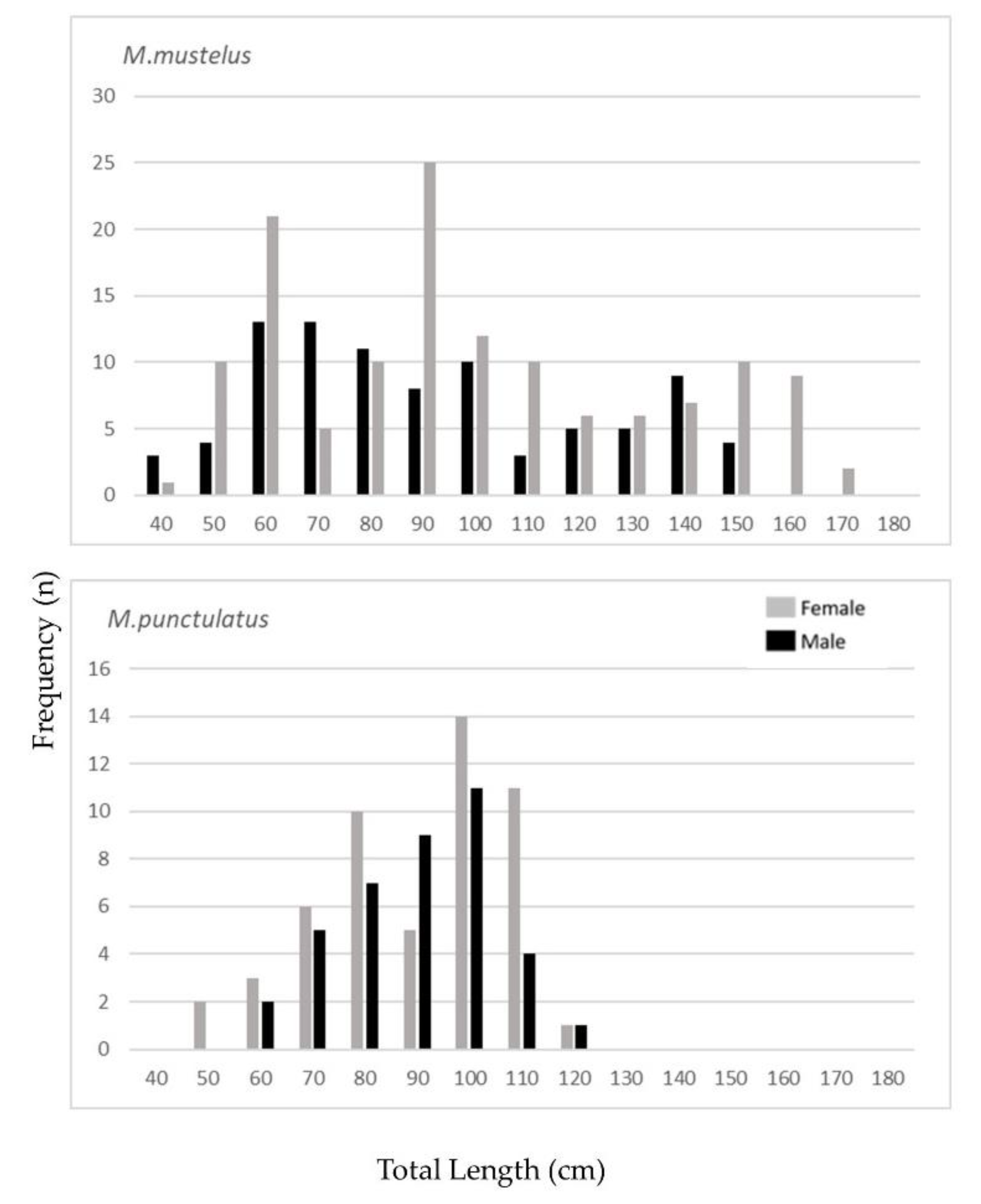
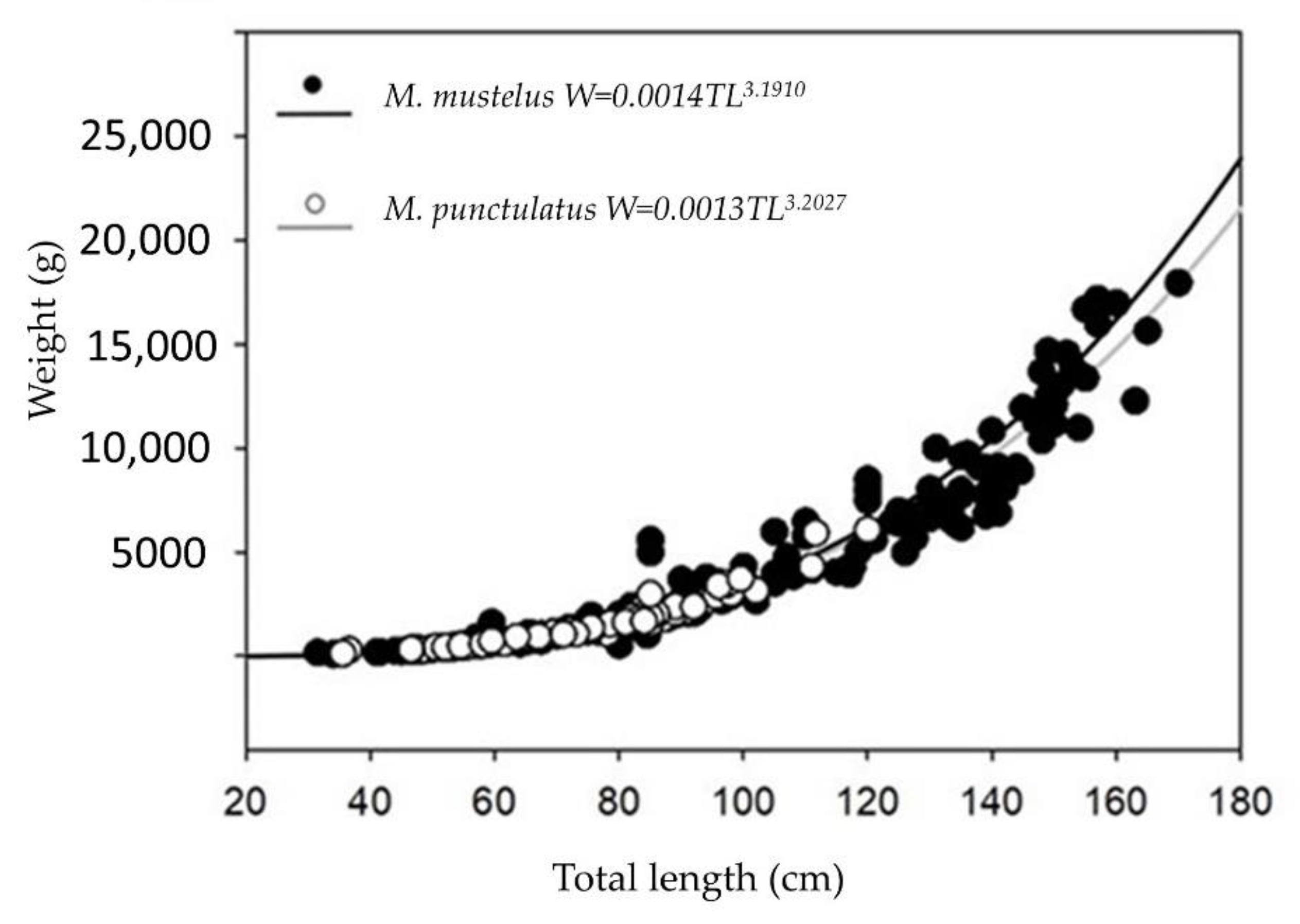
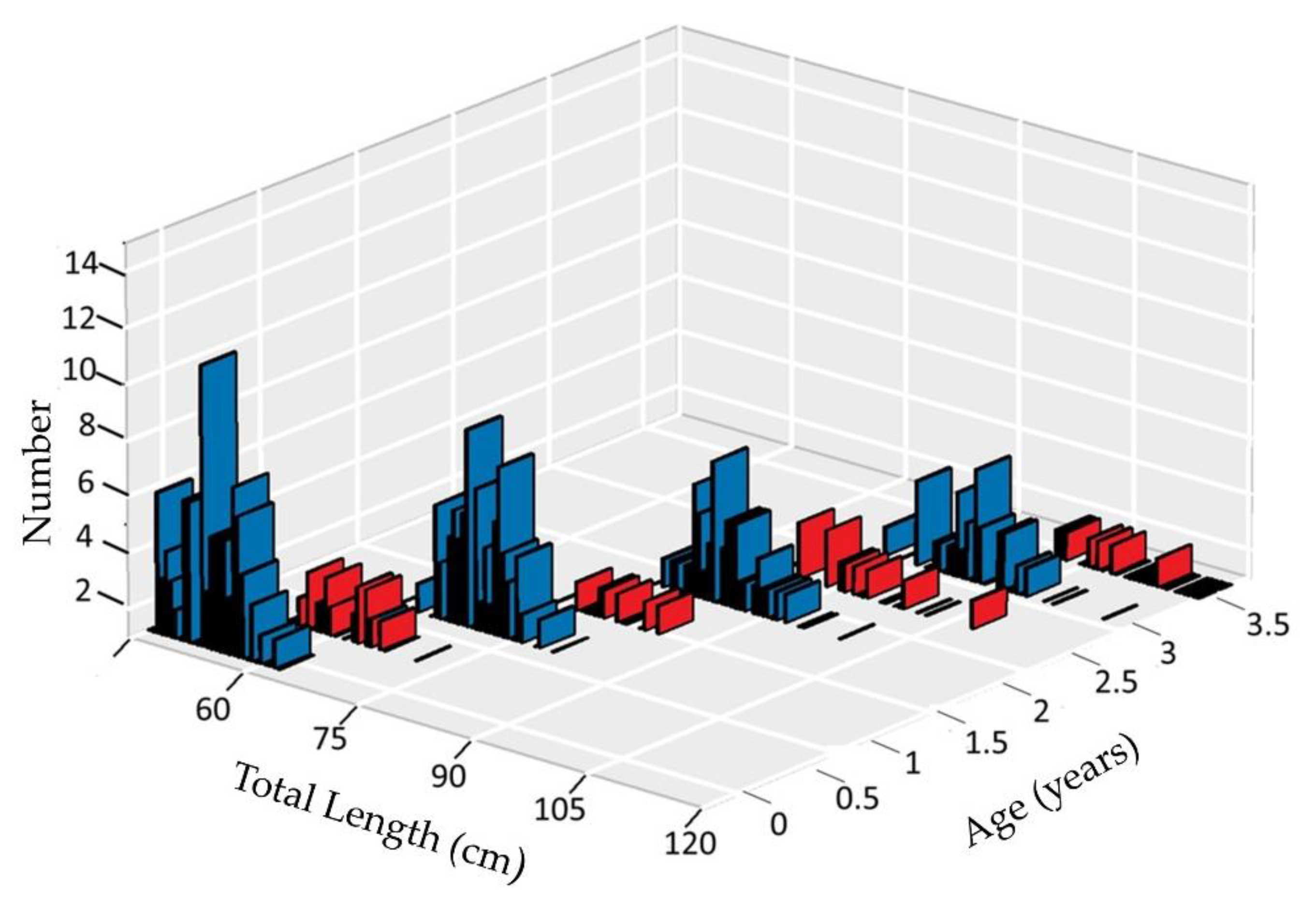
3.1. Smooth-Hounds’ Samples
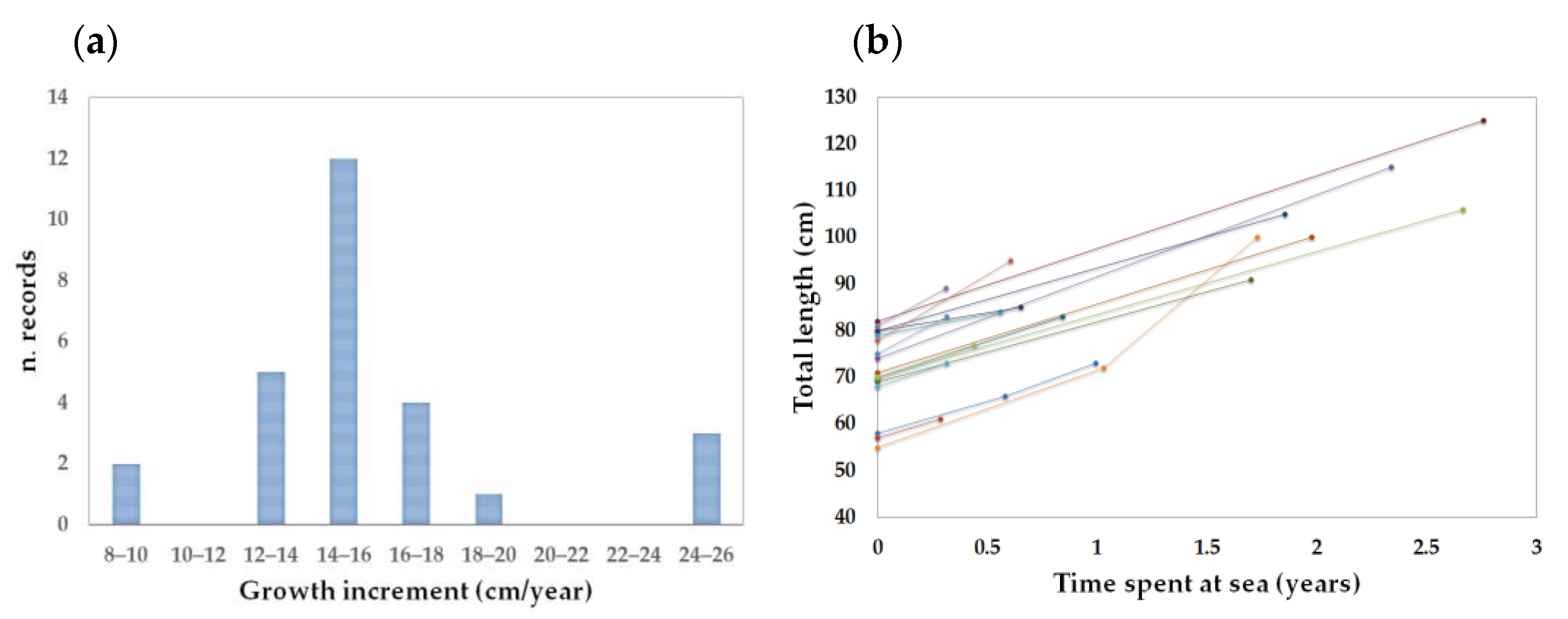
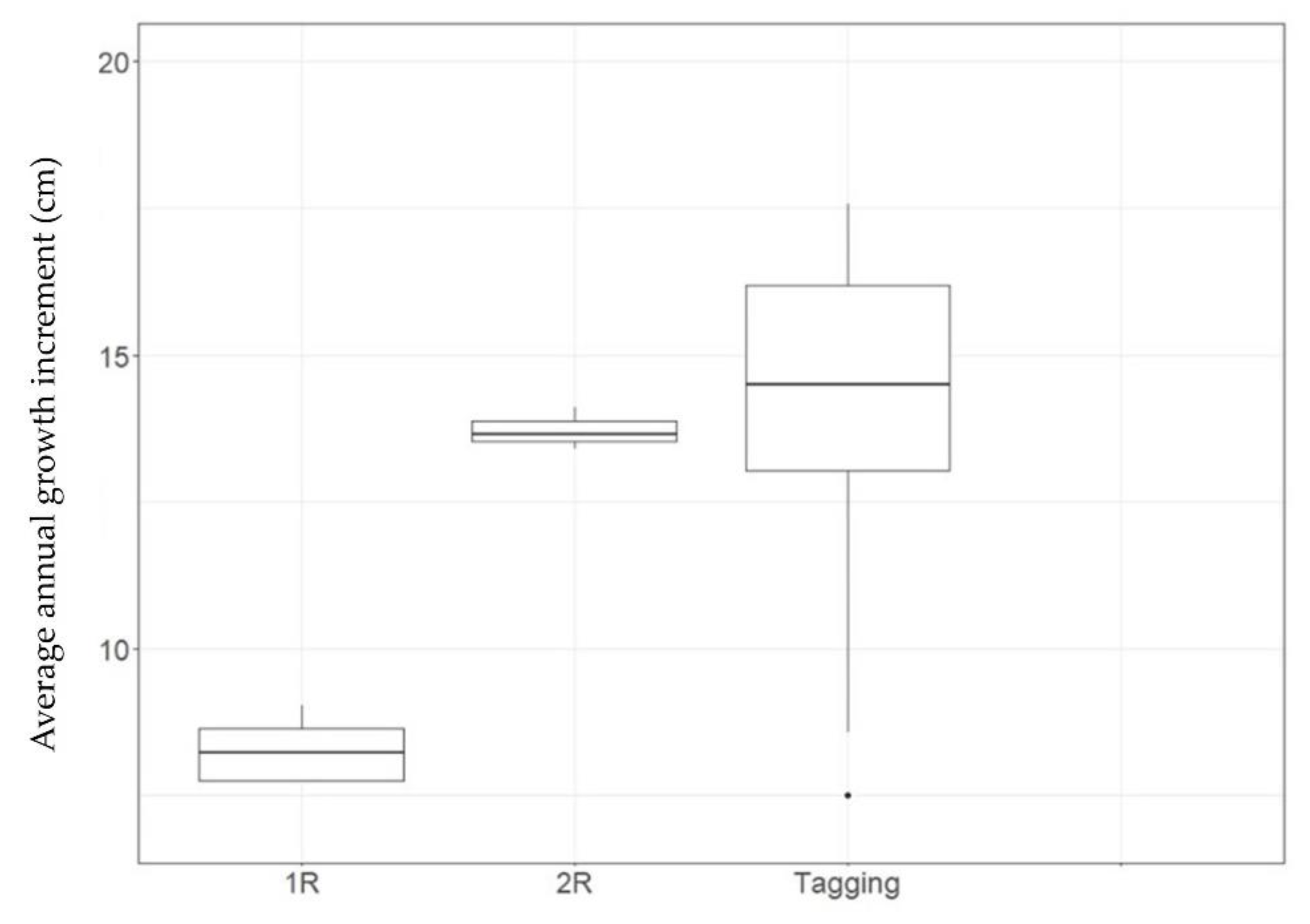
3.2. Tag-Recapture Data
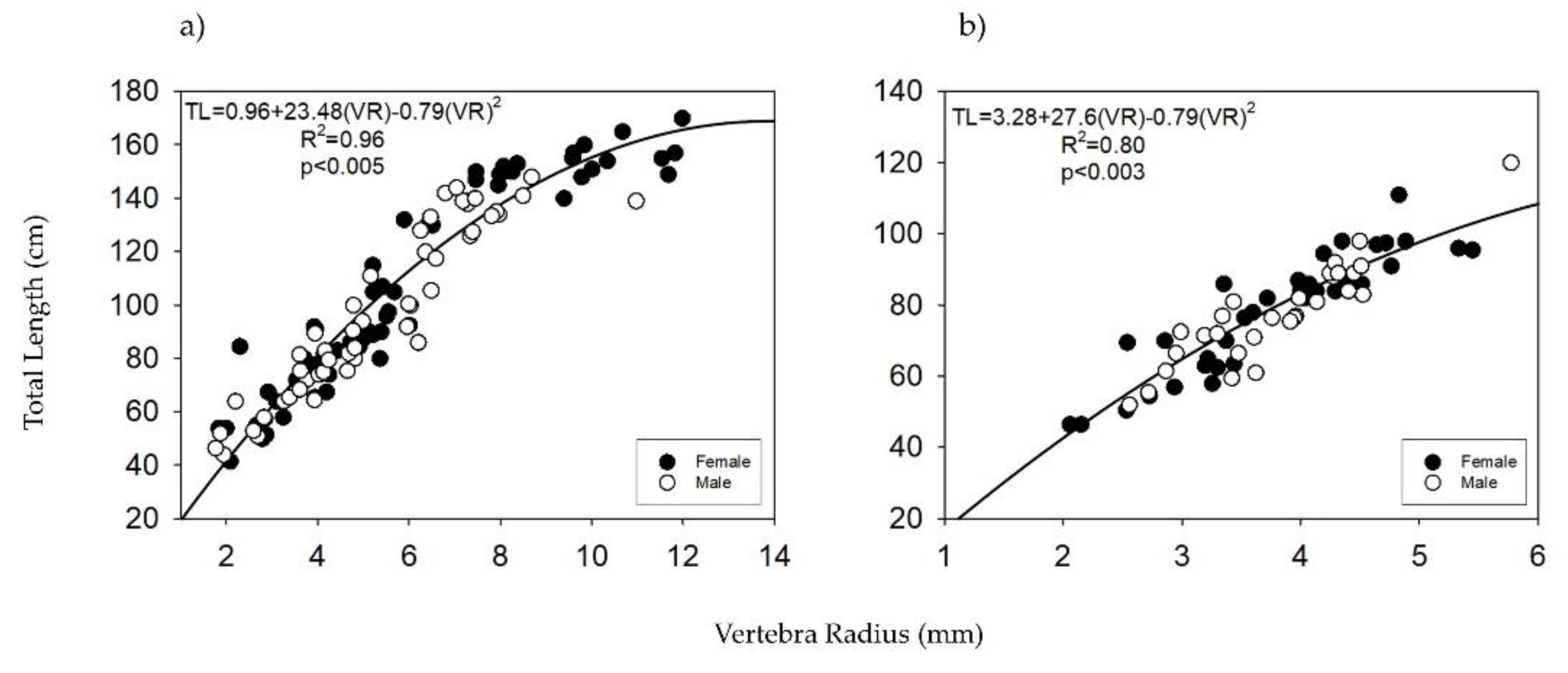
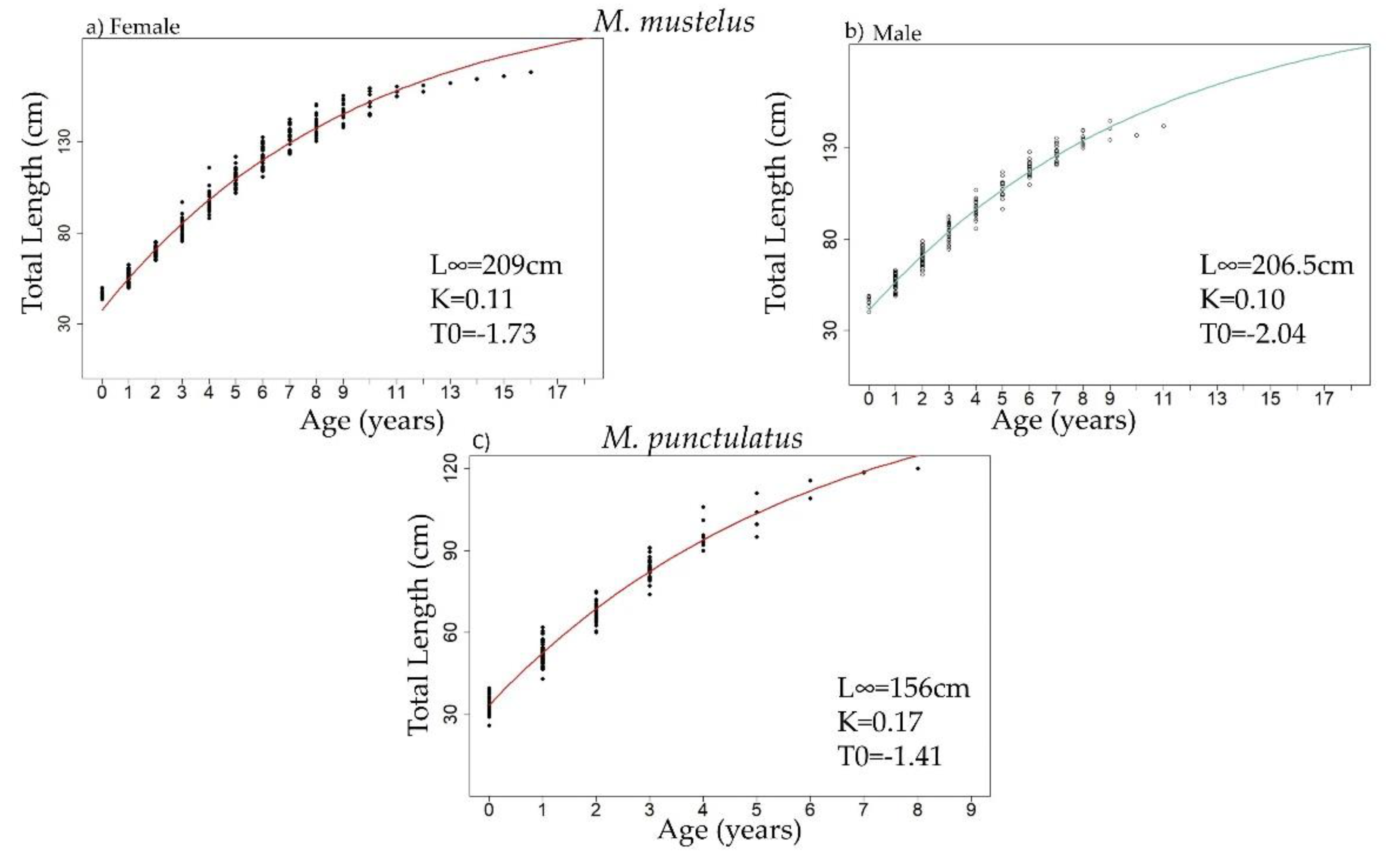
3.3. Age Estimation
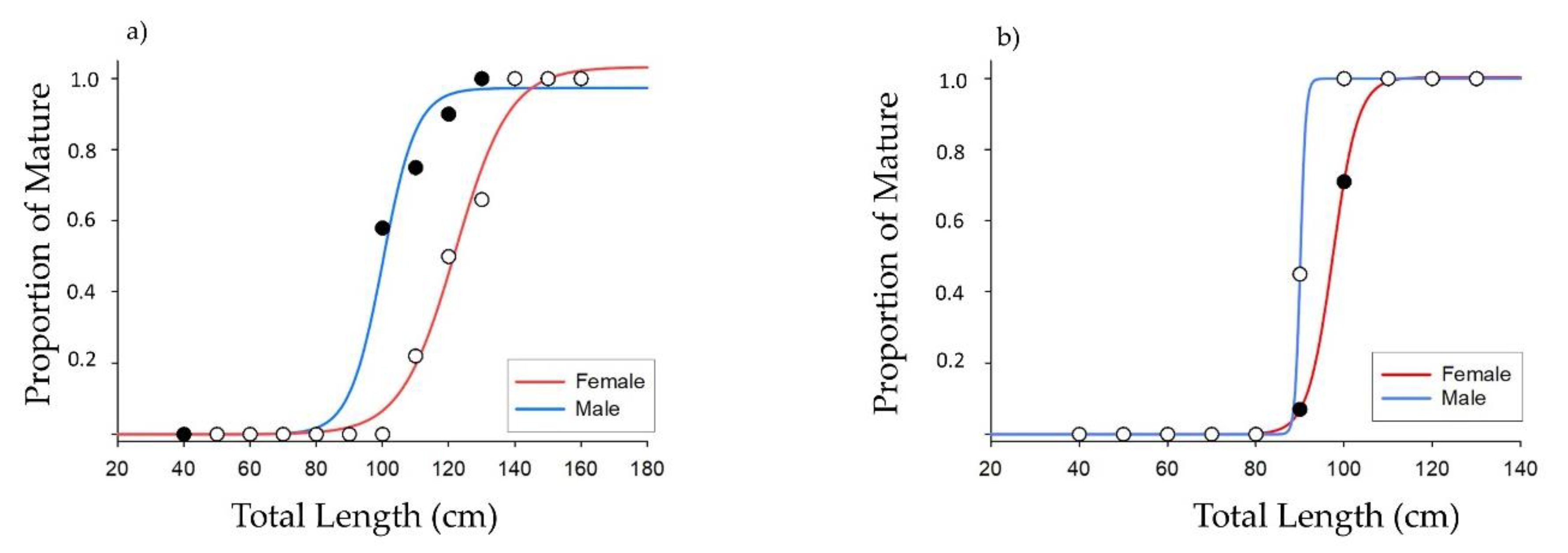
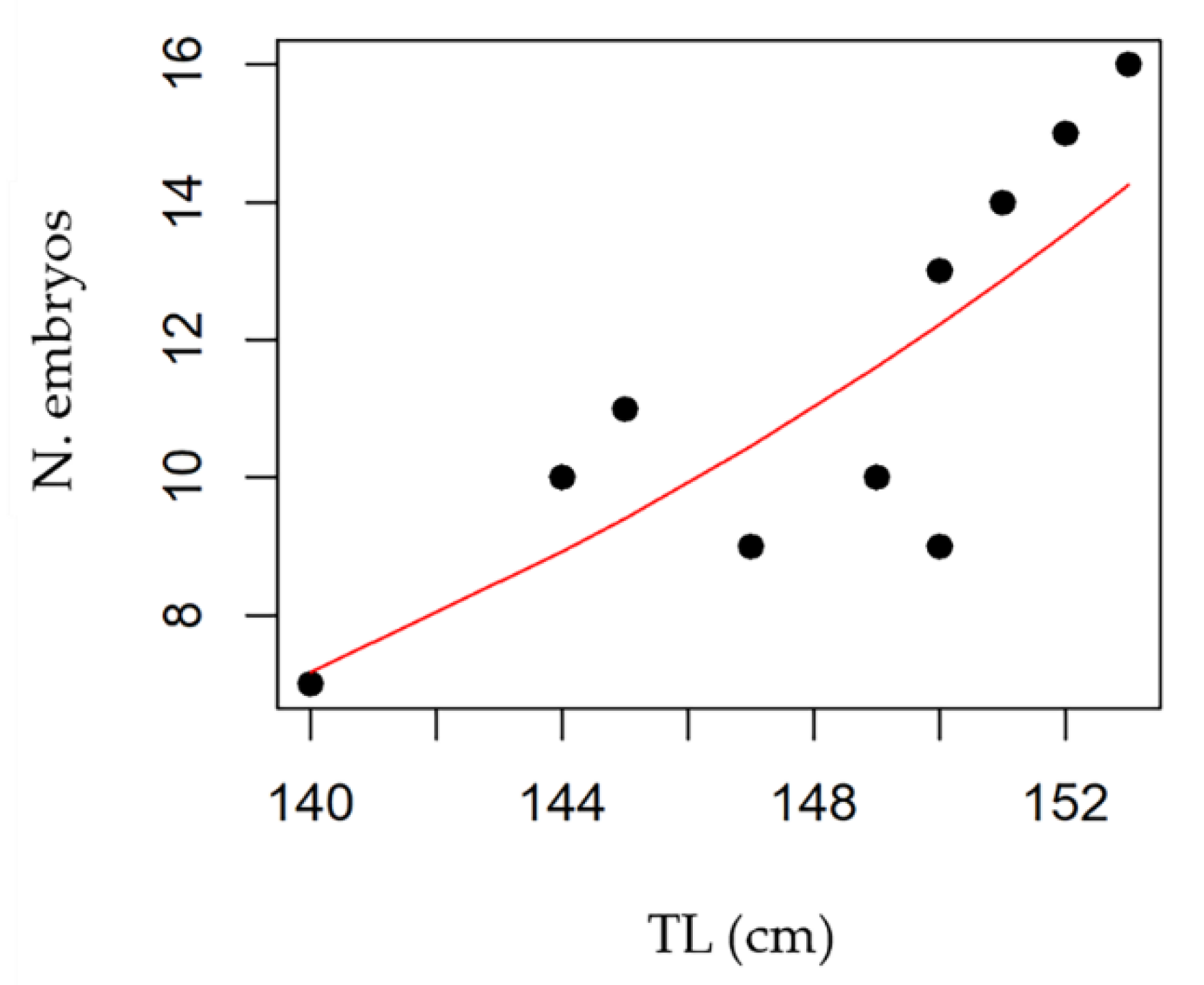
3.4. Maturity and Fecundity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCauley, D.J.; Pinsky, M.L.; Palumbi, S.R.; Estes, J.A.; Joyce, F.H.; Warner, R.R. Marine defaunation: Animal loss in the global ocean. Science 2015, 347, 1255641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, R.A.; Worm, B. Rapid worldwide depletion of predatory fish communities. Nature 2003, 423, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.K.; Myers, R.A.; Kehler, D.G.; Worm, B.; Harley, S.J.; Doherty, P.A. Collapse and conservation of shark populations in the Northwest Atlantic. Science 2003, 299, 389–392. [Google Scholar] [CrossRef]
- Baum, J.K.; Myers, R.A. Shifting baselines and the decline of pelagic sharks in the Gulf of Mexico. Ecol. Lett. 2004, 7, 135–145. [Google Scholar] [CrossRef]
- Shepherd, T.D.; Myers, R.A. Direct and indirect fishery effects on small coastal elasmobranchs in the northern Gulf of Mexico. Ecol. Lett. 2005, 8, 1095–1104. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787. [Google Scholar] [CrossRef]
- Maynou, F.; Sbrana, M.; Sartor, P.; Maravelias, C.; Kavadas, S.; Damalas, D.; Cartes, J.E.; Osio, G. Estimating Trends of Population Decline in Long-Lived Marine Species in the Mediterranean Sea Based on Fishers’ Perceptions. PLoS ONE 2011, 6, e21818. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, F.; Myers, R.A.; Serena, F.; Lotze, H.K. Loss of Large Predatory Sharks from the Mediterranean Sea. Conserv. Biol. 2008, 22, 952–964. [Google Scholar] [CrossRef]
- Colloca, F.; Enea, M.; Ragonese, S.; Di Lorenzo, M. A century of fishery data documenting the collapse of smooth-hounds (Mustelus spp.) in the Mediterranean Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 1145–1155. [Google Scholar] [CrossRef]
- Colloca, F.; Carrozzi, V.; Simonetti, A.; Di Lorenzo, M. Using Local Ecological Knowledge of Fishers to Reconstruct Abundance Trends of Elasmobranch Populations in the Strait of Sicily. Front. Mar. Sci. 2020, 7, 508. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Calò, A.; Di Franco, A.; Milisenda, G.; Aglieri, G.; Cattano, C.; Milazzo, M.; Guidetti, P. Small-scale fisheries catch more threatened elasmobranchs inside partially protected areas than in unprotected areas. Nat. Commun. 2022, 13, 4381. [Google Scholar] [CrossRef] [PubMed]
- Dulvy, N.K.; Allen, D.J.; Ralph, G.M.; Walls, R.H.L. The Conservation Status of Sharks, Rays, and Chimaeras in the Mediterranean Sea; IUCN: Malaga, Spain, 2016; p. 236. [Google Scholar] [CrossRef]
- Carpentieri, P.; Nastasi, A.; Sessa, M.; Srour, A. Incidental Catch of Vulnerable Species in Mediterranean and Black Sea fisheries—A Review; Studies and Reviews No. 100; General Fisheries Commission for the Mediterranean: Rome, Italy, 2021; p. 317. [Google Scholar] [CrossRef]
- Follesa, M.C.; Marongiu, M.F.; Zupa, W.; Bellodi, A.; Cau, A.; Cannas, R.; Colloca, F.; Djurovic, M.; Isajlovic, I.; Jadaud, A.; et al. Spatial variability of Chondrichthyes in the northern Mediterranean. Sci. Mar. 2019, 83, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Barbato, M.; Barría, C.; Bellodi, A.; Bonanomi, S.; Borme, D.; Ćetković, I.; Colloca, F.; Colmenero, A.I.; Crocetta, F.; De Carlo, F.; et al. The use of fishers’ Local Ecological Knowledge to reconstruct fish behavioural traits and fishers’ perception of conservation relevance of elasmobranchs in the Mediterranean Sea. Mediterr. Mar. Sci. 2021, 22, 603–622. [Google Scholar] [CrossRef]
- Farrell, E.D.; Clarke, M.W.; Mariani, S. A simple genetic identification method for Northeast Atlantic smoothhound sharks (Mustelus spp.). ICES J. Mar. Sci. 2009, 66, 561–565. [Google Scholar] [CrossRef]
- Marino, I.A.M.; Finotto, L.; Colloca, F.; Di Lorenzo, M.; Gristina, M.; Farrell, E.D.; Zane, L.; Mazzoldi, C. Resolving the ambiguities in the identification of two smooth-hound sharks (Mustelus mustelus and Mustelus punctulatus) using genetics and morphology. Mar. Biodivers. 2017, 48, 1551–1562. [Google Scholar] [CrossRef]
- Capapé, C.; Quignard, J.P. Contribution a la biologie des Triakidae des cotes tunisiennes. I. Mustelus mediterraneus Quignard et Capape, 1972, répartition géographique et bathymétrique, migrations et déplacements, reproduction, fécondite. Bull. Off. Nat. Peches Tunis. 1977, 1, 173–179. [Google Scholar]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A. Reproductive biology of the smooth-hound shark Mustelus mustelus (L.) in the Gulf of Gabès (south-central Mediterranean Sea). J. Fish Biol. 2008, 72, 1343–1354. [Google Scholar] [CrossRef]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A. Reproductive biology and diet of Mustelus punctulatus (Risso, 1826) (Chondrichthyes: Triakidae) from the Gulf of Gabès, central Mediterranean Sea. Sci. Mar. 2009, 73, 249–258. [Google Scholar] [CrossRef]
- Riginella, E.; Correale, V.; Marino, I.A.M.; Rasotto, M.B.; Vrbatovic, A.; Zane, L.; Mazzoldi, C. Contrasting life-history traits of two sympatric smooth-hound species: Implication for vulnerability. J. Fish Biol. 2020, 96, 853–857. [Google Scholar] [CrossRef]
- Costantini, M.; Bernardini, M.; Cordone, P.; Guilianini, P.G.; Orel, G. Observations on fishery, feeding habits and reproductive biology of Mustelus mustelus (Chondrichtyes, Triakidae) in Northern Adriatic Sea. Biol. Mar. Medit. 2000, 7, 427–432. [Google Scholar]
- Gračan, R.; Polak, T.; Lazar, B. Life history traits of the Blackspotted smooth-hound Mustelus punctulatus (Carcharhiniformes: Triakidae) in the Adriatic Sea. Nat. Croat. 2021, 30, 475–492. [Google Scholar] [CrossRef]
- Ozcan, E.; Basusta, N. Preliminary study on age, growth and reproduction of Mustelus mustelus (Elasmobranchii: Carcharhiniformes: Triakidae) inhabiting the Gulf of Iskenderun, north-eastern Mediterranean Sea. Acta Ichthyol. Piscat. 2018, 48, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Saïdi, B.; Enajjar, S.; Bradaï, M.N.; Bouaïn, A. Diet composition of smooth-hound shark, Mustelus mustelus (Linnaeus, 1758), in the Gulf of Gabès, southern Tunisia. J. Appl. Ichthyol. 2009, 25, 113–118. [Google Scholar] [CrossRef]
- Jardas, I.; Šantić, M.; Nerlović, V.; Pallaoro, A. Diet composition of blackspotted smooth-hound, Mustelus punctulatus (Risso, 1826), in the eastern Adriatic Sea. J. Appl. Ichthyol. 2007, 23, 279–281. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Vizzini, S.; Signa, G.; Andolina, C.; Palo, G.B.; Gristina, M.; Mazzoldi, C.; Colloca, F. Ontogenetic trophic segregation between two threatened smooth-hound sharks in the Central Mediterranean Sea. Sci. Rep. 2020, 10, 11011. [Google Scholar] [CrossRef]
- Kara, A.F.; Al Hajaji, M.; Ghmati, H.; Shakman, E.A. Growth & Reproduction of Mustelus mustelus (Chondrichthyes: Triakidae) in the south Mediterranean (Libyan coast). Libyan J. Ecol. Env. Sci. Tech. 2019, 1, 31–38. [Google Scholar]
- Goosen, A.J.J.; Smale, M.J. A preliminary study of age and growth of the smoothhound shark Mustelus mustelus (Triakidae). S. Afr. J. Mar. Sci. 1997, 18, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Smale, M.J.; Compagno, L.J.V. Life history and diet of two southern African smoothhound sharks, Mustelus mustelus (Linnaeus, 1758) and Mustelus palumbes Smith, 1957 (Pisces: Triakidae). Afr. J. Mar. Sci. 1997, 18, 229–248. [Google Scholar] [CrossRef]
- da Silva, C.; Attwood, C.G.; Wintner, S.P.; Wilke, C.G.; Winker, H.; Smale, M.J.; Kerwath, S.E. Life history of Mustelus mustelus in the Langebaan Lagoon marine protected area. Mar. Freshw. Res. 2021, 72, 1142–1159. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Sinerchia, M.; Colloca, F. The North sector of the Strait of Sicily: A priority area for conservation in the Mediterranean Sea. Hydrobiologia 2018, 821, 235–253. [Google Scholar] [CrossRef]
- Falsone, F.; Scannella, D.; Geraci, M.; Vitale, S.; Colloca, F.; Di Maio, F.; Milisenda, G.; Gancitano, V.; Bono, G.; Fiorentino, F. Identification and characterization of trammel net métiers: A case study from the southwestern Sicily (Central Mediterranean). Reg. Stud. Mar. Sci. 2020, 39, 101419. [Google Scholar] [CrossRef]
- Jorgensen, S.; Micheli, F.; White, T.; Van Houtan, K.; Alfaro-Shigueto, J.; Andrzejaczek, S.; Arnoldi, N.; Baum, J.; Block, B.; Britten, G.; et al. Emergent research and priorities for shark and ray conservation. Endanger. Species Res. 2022, 47, 171–203. [Google Scholar] [CrossRef]
- Ragonese, S.; Vitale, S.; Dimech, M.; Mazzola, S. Abundances of demersal sharks and chimaera from 1994-2009 scientific surveys in the Central Mediterranean Sea. PLoS ONE 2013, 8, e74865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauria, V.; Gristina, M.; Attrill, M.J.; Fiorentino, F.; Garofalo, G. Predictive habitat suitability models to aid conservation of elasmobranch diversity in the central Mediterranean Sea. Sci. Rep. 2015, 5, 13245. [Google Scholar] [CrossRef] [Green Version]
- Cailliet, G.M.; Smith, W.D.; Mollet, H.F.; Goldman, K.J. Age and growth studies of chondrichthyan fishes: The need for consistency in terminology, verification, validation, and growth function fitting. J. Appl. Phycol. 2006, 77, 211–228. [Google Scholar] [CrossRef]
- Goldman, K.J. Age and Growth of Elasmobranch Fishes. In Elasmobranch Fisheries Management Techniques; Musick, J.A., Bonfil, R., Eds.; APEC Fisheries Working Group: Singapore, 2004; pp. 97–132. ISBN 981-04-9682-6. [Google Scholar]
- Campana, S.E. Age Determination of Elasmobranchs with Special Reference to Mediterranean Species: A Technical Manual; FAO-GFCM: Rome, Italy, 2014; Studies and Review; Volume 94, ISBN 9789251083208. [Google Scholar]
- Cailliet, G.M.; Goldman, K.J. Age determination and validation in chondrichthyan fishes. In Biology of Sharks and Their Rela-Tives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 399–447. [Google Scholar]
- Campana, S.E. How Reliable are Growth Back-Calculations Based on Otoliths? Can. J. Fish. Aquat. Sci. 1990, 47, 2219–2227. [Google Scholar] [CrossRef]
- Ogle, D.; Wheeler, P.; Dinno, A. Package ‘FSA’. R Package Version 0.8.31. 2020. Available online: https://cran.r-project.org/web/packages/FSA/index.html (accessed on 10 July 2022).
- Nelson, G.A. Package ‘Fishmethods‘: Fishery Science Methods and Models. R Package Version 1.11-3. 2022. Available online: https://cran.r-project.org/web/packages/fishmethods/fishmethods.pdf (accessed on 15 February 2022).
- Conrath, C.L.; Musick, J.A. Reproductive biology of the smooth dogfish, Mustelus canis, in the Northwest Atlantic Ocean. Environ. Biol. Fishes 2002, 64, 367–377. [Google Scholar] [CrossRef]
- Hutchings, J. A Life Histories of Fish. In Handbook of Fish Biology and Fisheries; Hart, P.J.B., Reynolds, D., Eds.; Blackwell Science Ltd.: Oxford, UK, 2002; Volume 1, pp. 149–174. [Google Scholar]
- Moulton, P.; Walker, T.; Saddlier, S. Age and growth studies of Gummy Shark, Mustelus antarcticus Gunther, and School Shark, Galeorhinus galeus (Linnaeus), from Southern Australian Waters. Mar. Freshw. Res. 1992, 43, 1241–1267. [Google Scholar] [CrossRef]
- Conrath, C.L.; Gelsleichter, J.; Musick, J.A. Age and growth of the smooth dogfish (Mustelus canis) in the northwest Atlantic Ocean. Fish. Bull. 2002, 100, 674–682. [Google Scholar]
- Rigby, C.L.; White, W.T.; Smart, J.J.; Simpfendorfer, C.A. Life histories of two deep-water Australian endemic elasmobranchs: Argus skate Dipturus polyommata and eastern spotted gummy shark Mustelus walkeri. J. Fish Biol. 2016, 88, 1149–1174. [Google Scholar] [CrossRef] [PubMed]
- Farrell, E.D.; Mariani, S.; Clarke, M.W. Age and growth estimates for the starry smoothhound (Mustelus asterias) in the Northeast Atlantic Ocean. ICES J. Mar. Sci. 2010, 67, 931–939. [Google Scholar] [CrossRef]
- Yudin, K.G.; Cailliet, G.M. Age and growth of the gray smoothhound, Mustelus californicus, and the brown smoothhound, M. henlei, sharks from central California. Copeia 1990, 191–204. [Google Scholar] [CrossRef]
- Francis, M.; Francis, R. Growth rate estimates for New Zealand Rig (Mustelus lenticulatus). Mar. Freshw. Res. 1992, 43, 1157–1176. [Google Scholar] [CrossRef]
- Molina, J.M.; Blasina, G.E.; Cazorla, A.C.L. Age and growth of the highly exploited narrownose smooth-hound (Mustelus schmitti) (Pisces: Elasmobranchii). Fish. Bull. 2017, 115, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Mizue, K. Studies on sharks—XV. Age and growth of Japanese dogfish Mustelus manazo Bleeker in the East China Sea. Bull. Jap. Soc. Sci. Fish 1979, 45, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Cortés, E. Life history patterns and correlations in sharks. Rev. Fish. Sci. 2000, 8, 299–344. [Google Scholar] [CrossRef]
- Walker, T. Fishery simulation model for sharks applied to the Gummy Shark, Mustelus antarcticus Gunther, from Southern Australian waters. Mar. Freshw. Res. 1992, 43, 195–212. [Google Scholar] [CrossRef]
- Colautti, D.; Baigun, C.; Cazorla, A.L.; Llompart, F.; Molina, J.M.; Suquele, P.; Calvo, S. Population biology and fishery characteristics of the smooth-hound Mustelus schmitti in Anegada Bay, Argentina. Fish. Res. 2010, 106, 351–357. [Google Scholar] [CrossRef]
- Bitalo, D.N.; Maduna, S.N.; da Silva, C.; Roodt-Wilding, R.; der Merwe, A.E.B.-V. Differential gene flow patterns for two commercially exploited shark species, tope (Galeorhinus galeus) and common smoothhound (Mustelus mustelus) along the south–west coast of South Africa. Fish. Res. 2015, 172, 190–196. [Google Scholar] [CrossRef]
- Fisher, J.; Frank, K.; Leggett, W. Global variation in marine fish body size and its role in biodiversity–ecosystem functioning. Mar. Ecol. Prog. Ser. 2010, 405, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Taniuchi, T.; Shimizu, M. Geographic Variation in Growth of the Starspotted Dogfish Mustelus manazo from Five Localities in Japan and Taiwan. Fish. Sci. 1998, 64, 732–739. [Google Scholar] [CrossRef]
- Finotto, L.; Gristina, M.; Garofalo, G.; Riginella, E.; Mazzoldi, C. Contrasting life history and reproductive traits in two populations of Scyliorhinus canicula. Mar. Biol. 2015, 162, 1175–1186. [Google Scholar] [CrossRef]
- Klein, J.D.; Asbury, T.A.; da Silva, C.; Hull, K.L.; Dicken, M.L.; Gennari, E.; Maduna, S.N.; der Merwe, A.E.B. Site fidelity and shallow genetic structure in the common smooth-hound shark Mustelus mustelus confirmed by tag-recapture and genetic data. J. Fish Biol. 2021, 100, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Brevé, N.W.P.; Winter, H.V.; Wijmans, P.A.D.M.; Greenway, E.S.I.; Nagelkerke, L.A.J. Sex differentiation in seasonal distribution of the starry smooth-hound Mustelus asterias. J. Fish Biol. 2020, 97, 1870–1875. [Google Scholar] [CrossRef]
- Brevé, N.W.P.; Winter, H.V.; Van Overzee, H.M.J.; Farrell, E.D.; Walker, P.A. Seasonal migration of the starry smooth-hound shark Mustelus asterias as revealed from tag-recapture data of an angler-led tagging programme. J. Fish Biol. 2016, 89, 1158–1177. [Google Scholar] [CrossRef]
- Chapman, D.D.; Feldheim, K.A.; Papastamatiou, Y.P.; Hueter, R.E. There and Back Again: A Review of Residency and Return Migrations in Sharks, with Implications for Population Structure and Management. Annu. Rev. Mar. Sci. 2015, 7, 547–570. [Google Scholar] [CrossRef]
- Sandoval-Castillo, J.; Beheregaray, L. Metapopulation structure informs conservation management in a heavily exploited coastal shark (Mustelus henlei). Mar. Ecol. Prog. Ser. 2015, 533, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Hueter, R.E.; Mann, D.E.; Maruska, K.P.; Sisneros, J.A.; Demski, L.S. Sensory biology of elasmobranchs. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 325–368. [Google Scholar]
- Sacchi, J. Overview of Mitigation Measures to Reduce the Incidental Catch of Vulnerable Species in Fisheries; Studies and Reviews No. 100; General Fisheries Commission for the Mediterranean: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A. Demographic analysis of the dusky shark fishery in southwestern Australia. In Life in the Slow Lane: Ecology and Conservation of Long Lived Marine Animals; Musick, J.A., Ed.; American Fishery Society Symposium: Bethesda, MD, USA, 1999; pp. 149–160. [Google Scholar]
- Prince, J.D. Gauntlet Fisheries for Elasmobranchs—the Secret of Sustainable Shark Fisheries. J. Northwest Atl. Fish. Sci. 2005, 35, 407–416. [Google Scholar] [CrossRef]
- Stöhr, C.; Lundholm, C.; Crona, B.; Chabay, I. Stakeholder participation and sustainable fisheries: An integrative framework for assessing adaptive comanagement processes. Ecol. Soc. 2014, 19, 14. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mustelus mustelus | Mustelus punctulatus | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | ||||||||||||
| 1 | 22 | 20 | 29 | 25 | 30 | 31 | 32 | 1 | 22 | 24 | 21 | |
| L∞ F | 209 | / | / | / | / | 204.9 | / | 189.7 | / | / | 208.4 | / |
| kF | 0.11 | / | / | / | / | 0.06 | / | 0.11 | / | / | 0.04 | / |
| t0 F | −1.73 | / | / | / | / | −3.35 | / | −2.08 | / | / | −4.49 | / |
| L∞ M | 206 | / | / | / | / | 145.1 | / | 112 | / | / | 136.4 | / |
| kM | 0.10 | / | / | / | / | 0.12 | / | 0.41 | / | / | 0.10 | / |
| t0 M | −2.04 | / | / | / | / | −2.14 | / | −1.26 | / | / | −2.53 | / |
| L∞ F+M | / | / | / | 194.9 | 195.1 | / | / | / | 156 | / | / | / |
| kF+M | / | / | / | 0.03 | 0.06 | / | / | / | 0.17 | / | / | / |
| t0 F+M | / | / | / | −9.31 | −4.27 | / | / | / | −1.41 | / | / | / |
| Agemax F | 16 | / | / | 28 | 25 | 24 | / | 13 | 6 | / | 19 | / |
| Agemax M | 12 | / | / | 21 | 20 | 17 | / | 11 | 8 | / | 14 | / |
| Agemax F+M | / | / | / | / | / | / | / | / | / | |||
| A50F | 5 | / | / | / | 8 | 12–15 | / | 6.2 | 11 | / | 12.5 | / |
| A50M | 4 | / | / | / | 7 | 6–9 | / | 3.2 | 9 | / | 6.6 | / |
| Lmax F | 170 | 158 | 165 | 168 | 162.6 | 164 | 165 | 173.4 | 111 | 141 | 136.2 | 122 |
| Lmax M | 148 | 150 | 144.5 | 129.2 | 149 | 145 | 145 | 126.7 | 120 | 119 | 126.5 | 111 |
| L50F | 111.5 | 121.2 | 117.2 | 85 | 109 | / | 125 * | 119.4 | 92.5 | 109.9 | 100 | 95.6 |
| L50M | 92.5 | 108.1 | 97.1 | 75 | 92 | / | 95 * | 96.7 | 84.5 | 91.3 | 83.1 | 81.4 |
| Litter size | 8–16 | 3–18 | 4–18 | 8–24 | 10–15 | / | 2–23 | 2–26 | 11–18 | 7–35 | / | 12–27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boscolo Palo, G.; Di Lorenzo, M.; Gancitano, S.; Ragonese, S.; Mazzoldi, C.; Colloca, F. Sharks Do Not Always Grow Slowly: Tagging Data Reveal a Different Pattern of Growth, Longevity and Maturity for Threatened Smooth-Hounds in the Central Mediterranean Sea. J. Mar. Sci. Eng. 2022, 10, 1647. https://doi.org/10.3390/jmse10111647
Boscolo Palo G, Di Lorenzo M, Gancitano S, Ragonese S, Mazzoldi C, Colloca F. Sharks Do Not Always Grow Slowly: Tagging Data Reveal a Different Pattern of Growth, Longevity and Maturity for Threatened Smooth-Hounds in the Central Mediterranean Sea. Journal of Marine Science and Engineering. 2022; 10(11):1647. https://doi.org/10.3390/jmse10111647
Chicago/Turabian StyleBoscolo Palo, Gabriele, Manfredi Di Lorenzo, Salvatore Gancitano, Sergio Ragonese, Carlotta Mazzoldi, and Francesco Colloca. 2022. "Sharks Do Not Always Grow Slowly: Tagging Data Reveal a Different Pattern of Growth, Longevity and Maturity for Threatened Smooth-Hounds in the Central Mediterranean Sea" Journal of Marine Science and Engineering 10, no. 11: 1647. https://doi.org/10.3390/jmse10111647