
The Isolation of Vibrio crassostreae and V. cyclitrophicus in Lesser-Spotted Dogfish (Scyliorhinus canicula) Juveniles Reared in a Public Aquarium

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Outbreak Description and Conditions of Sampling Procedure
2.2. Necropsy
2.3. Parasitological Examination
2.4. Virological Examination
2.5. Bacteriological and Biochemical Analyses
2.6. Biomolecular Analysis for Bacterial Identification
2.7. Phylogenetic Analysis
2.8. Statistical Analysis
3. Results
3.1. Necropsy, Parasitological, and Virological Examinations
3.2. Bacteriological Analysis
3.3. Biomolecular Analysis for Bacterial Identification
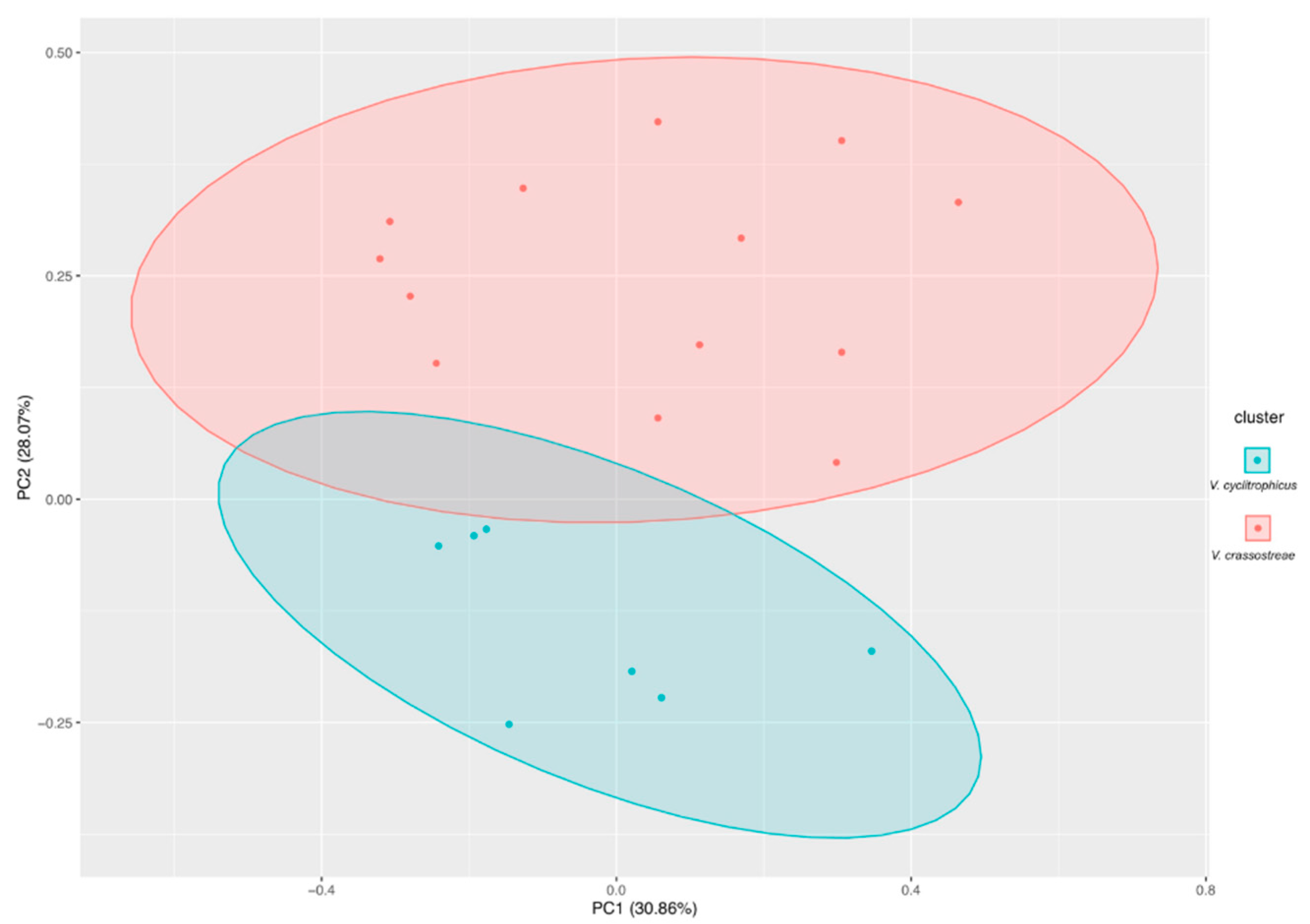
3.4. Phylogenetic Analysis
3.5. Biochemical Features of Isolates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reen, F.J.; Almagro-Moreno, S.; Ussery, D.; Boyd, E.F. The genomic code inferring Vibrionaceae niche specialization. Nat. Rev. Microbiol. 2006, 9, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.N. Fitness factors in vibrios: A mini-review. Microb. Ecol. 2013, 65, 826–851. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R.; Soira, W.M. The ecology of Vibrio Cholera. In Cholera; Barua, D., Greenough, W.B., Eds.; Springer: Boston, MA, USA, 1992; pp. 1–36. ISBN 978-1-4757-9690-2. [Google Scholar]
- Sorum, H.; Myhr, E.; Zwicker, B.M.; Lillehaug, A. Comparison by plasmid profiling of Vibrio salmonicida strains isolated from diseased fish from different north European and Canadian coastal areas of the Atlantic Ocean. Can. J. Fish Aquat. Sci. 1993, 50, 247–250. [Google Scholar] [CrossRef]
- Diggles, B.K.; Carson, J.; Hine, P.M.; Hickman, R.W.; Tait, M.J. Vibrio species associated with mortalities in hatchery-reared turbot (Colistium nudipinnis) and brill (C. guntheri) in New Zealand. Aquaculture 2000, 183, 1–12. [Google Scholar] [CrossRef]
- Goarant, C.; Herlin, J.; Brizard, R.; Marteau, A.L.; Martin, C.; Martin, B. Toxic factors of Vibrio strains pathogenic to shrimp. Dis. Aquat. Org. 2000, 40, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.G.; Nakamura, A.; Mori, K. Inhibitory effects of ovoglobulins on bacillary necrosis in larvae of the Pacific oyster, Crassostrea gigas. J. Invertebr. Pathol. 2000, 75, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Paillard, C.; Le Roux, F.; Borrego, J.J. Bacterial disease in marine bivalves, a review of recent studies: Trends and evolution. Aquat. Living Resour. 2004, 17, 477–498. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.C.; Lin, J.Y.; Hsiao, P.T.; Lee, K.K. Isolation and characterization of pathogenic Vibrio alginolyticus from diseased cobia Rachycentron canadum. J. Basic Microbiol. 2004, 44, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.H.; Sun, J.J.; Liu, L.; Zhao, H.H.; Wang, H.F.; Liang, L.Q.; Liu, L.B.; Li, G.F. Characterization of two phenotypes of Photobacterium damselae subsp. damselae isolated from diseased juvenile Trachinotus ovatus reared in cage mariculture. J. World Aquacult. Soc. 2009, 40, 281–289. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Y.W.; Mo, Z.Q.; Luo, X.C.; Sun, H.Y.; Liu, P.; Li, A.X.; Zhou, S.M.; Dan, X.M. Outbreak of a novel disease associated with Vibrio mimicus infection in fresh water cultured yellow catfish, Pelteobagrus fulvidraco. Aquaculture 2014, 432, 119–124. [Google Scholar] [CrossRef]
- Rajan, P.R.; Lopez, C.; Lin, J.H.; Yang, H. Vibrio alginolyticus infection in cobia (Rachycentron canadum) cultured in Taiwan. Bull. Eur. Ass. Fish Pathol. 2001, 21, 228–234. [Google Scholar]
- Ransangan, J.; Mustafa, S. Identification of Vibrio harveyi isolated from diseased Asian seabass (Lates calcarifer) by use of 16S ribosomal DNA sequencing. J. Aquat. Anim. Health 2009, 21, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Austin, D. Bacterial Fish Pathogens: Diseases of Farmed and Wild Fish, 6th ed.; Springer International Publishing: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Akayli, T.; Timur, G.; Albayrak, G.; Aydemir, B. Identification and genotyping of Vibrio ordalii: A comparison of different methods. Isr. J. Aquacult-Bamid 2010, 62, 9–18. [Google Scholar] [CrossRef]
- Dong, H.T.; Taengphu, S.; Sangsuriya, P.; Charoensapsri, W.; Phiwsaiya, K.; Sornwatana, T.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Recovery of Vibrio harveyi from scale drop and muscle necrosis disease in farmed barramundi, Lates calcarifer in Vietnam. Aquaculture 2017, 473, 89–96. [Google Scholar] [CrossRef]
- Labella, A.; Vida, M.; Alonso, M.C.; Infante, C.; Cardenas, S.; Lopez–Romalde, S.; Manchado, M.; Borrego, J.J. First isolation of Photobacterium damselae ssp. damselae from cultured redbanded seabream, Pagrus auriga Valenciennes, in Spain. J. Fish Dis. 2006, 2, 175–179. [Google Scholar] [CrossRef]
- Nurliyana, M.; Amal, M.N.A.; Zamri-Saad, M.; Ina-Salwany, M.Y. Possible transmission routes of Vibrio spp. in tropical cage-cultured marine fishes. Lett. Appl. Microbiol. 2019, 68, 485–496. [Google Scholar] [CrossRef]
- Kim, M.N.; Bang, H.J. Detection of marine pathogenic bacterial Vibrio species by multiplex polymerase chain reaction (PCR). J. Environ. Biol. 2008, 29, 543–546. [Google Scholar]
- Sandlund, N.; Rødseth, O.M.; Knappskog, D.H.; Fiksdal, I.U.; Bergh, Ø. Comparative susceptibility of turbot, halibut, and cod yolk-sac larvae to challenge with Vibrio spp. Dis. Aquat. Org. 2010, 89, 29–37. [Google Scholar] [CrossRef]
- Haenen, O.L.M.; Van Zanten, E.; Jansen, R.; Roozenburg, I.; Engelsma, M.Y.; Dijkstra, A.; Möller, A.V.M. Vibrio vulnificus outbreaks in Dutch eel farms since 1996: Strain diversity and impact. Dis. Aquat. Org. 2014, 108, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Bellos, G.; Angelidis, P.; Miliou, H. Effect of temperature and seasonality principal epizootiological risk factor on vibriosis and photobacteriosis outbreaks for european sea bass in greece (1998–2013). J. Aquac. Res. Dev. 2015, 6, 10–4172. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.H.; Wu, K.; Chu, T.W.; Wu, T.M. Dietary supplementation of probiotic, Bacillus subtilis E20, enhances the growth performance and disease resistance against Vibrio alginolyticus in parrot fish (Oplegnathus fasciatus). Aquac. Int. 2018, 26, 63–74. [Google Scholar] [CrossRef]
- Grimes, D.J.; Colwell, R.R.; Stemmler, J.; Hada, H.; Maneval, D.; Hetrick, F.M.; Stoskopf, M. Vibrio species as agents of elasmobranch disease. Helgol. Meeresunters. 1984, 37, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Heng, S.P.; Letchumanan, V.; Deng, C.Y.; Ab Mutalib, N.S.; Khan, T.M.; Chuah, L.H.; Chan, K.G.; Goh, B.H.; Pusparajah, P.; Lee, L.H. Vibrio vulnificus: An environmental and clinical burden. Front. Microbiol. 2017, 8, 997. [Google Scholar] [CrossRef] [Green Version]
- Libralato, S.; Christensen, V.; Pauly, D. A method for identifying keystone species in food web models. Ecol. Modell. 2006, 195, 153–171. [Google Scholar] [CrossRef]
- Baum, J.K.; Worm, B. Cascading top-down effects of changing oceanic predator abundances. J. Anim. Ecol. 2009, 78, 699–714. [Google Scholar] [CrossRef]
- Ferretti, F.; Myers, R.A.; Serena, F.; Lotze, H.K. Loss of large predatory sharks from the Mediterranean Sea. Biol. Conserv. 2008, 22, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Voultsiadou, E. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, M.; Navarro, J.; Palomera, I. Ecological role, fishing impact, and management options for the recovery of a Mediterranean endemic skate by means of food web models. Biol. Conserv. 2013, 157, 108–120. [Google Scholar] [CrossRef]
- Malak, D.A. Overview of the Conservation Status of the Marine Fishes of the Mediterranean Sea; IUCN: Gland, Switzerland, 2011. [Google Scholar]
- Lyle, J.M. Food and feeding habits of the lesser spotted dogfish, Scyliorhinus canicula (L.) in Isle of Man waters. J. Fish. Biol. 1983, 23, 139–148. [Google Scholar] [CrossRef]
- Olaso, I.; Velasco, F.; Pérez, N. Importance of discarded blue whiting (Micromessistius poutassou) in the diet of lesser spotted dogfish (Scyliorhinus canicula) in the Cantabrian Sea. ICES J. Mar. Sci. 1998, 55, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.A.; Huveneers, C.; Peddemors, V.; Boomer, A.; Harcourt, R.G. Born to be free? Assessing the viability of releasing captive-bred wobbegongs to restock depleted populations. Front. Mar. Sci. 2015, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Terrell, S.P. An Introduction to Viral, Bacterial and Fungal Diseases of Elasmobranchs. In Elasmobranch Husbandry Manual: Captive Care of Sharks, Rays and Their Relatives; Smith, M., Warmolts, D., Thoney, D., Hueter, R., Eds.; Ohio Biological Survey Inc.: Columbus, OH, USA, 2004; p. 589. [Google Scholar]
- Bakopoulos, V.; Tsepa, E.; Diakou, A.; Kokkoris, G.; Kolygas, M.; Athanassopoulou, F. Parasites of Scyliorhinus canicula (Linnaeus, 1758) in the north-eastern Aegean Sea. J. Mar. Biolog. Assoc. 2018, 98, 2133–2143. [Google Scholar] [CrossRef]
- Gratzek, J.B.; Gilbert, J.P.; Lohr, A.L.; Shotts, E.B., Jr.; Brown, J. Ultraviolet light control of Ichthyophthirius multifiliis Fouquet in a closed fish culture recirculation system. J. Fish Dis. 1983, 6, 145–153. [Google Scholar] [CrossRef]
- Pratesi, R. L’interazione radiazione ottica-biomateria: Meccanismi d’azione, danni biologici e cautele d’uso. In Quaderni di Tecniche di Protezione Ambientale a Cura di Adriano Zavatti; Pitagora Editrice: Bologna, Italy, 1993. [Google Scholar]
- Panzarin, V.; Patarnello, P.; Mori, A.; Rampazzo, E.; Cappellozza, E.; Bovo, G.; Cattoli, G. Development and validation of a real-time TaqMan PCR assay for the detection of betanodavirus in clinical specimens. Arch. Virol. 2010, 155, 1193–1203. [Google Scholar] [CrossRef]
- Pastorino, P.; Colussi, S.; Pizzul, E.; Varello, K.; Menconi, V.; Mugetti, D.; Tomasoni, M.; Esposito, G.; Bertoli, M.; Bozzetta, E.; et al. The unusual isolation of carnobacteria in eyes of healthy salmonids in high-mountain lakes. Sci. Rep. 2021, 11, 2314. [Google Scholar] [CrossRef] [PubMed]
- Ki, J.S.; Zhang, R.; Zhang, W.; Huang, Y.L.; Qian, P.Y. Analysis of RNA polymerase beta subunit (rpoB) gene sequences for the discriminative power of marine Vibrio species. Microb. Ecol. 2009, 58, 679–691. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for Inferring Very Large Phylogenies by Using the Neighbor-Joining Method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Rychert, J.; Creely, D.; Mayo-Smith, L.M.; Calderwood, S.B.; Ivers, L.C.; Ryan, E.T.; Boncy, J.; Qadri, F.; Ahmed, D.; Ferraro, M.J.; et al. Evaluation of matrix-assisted laser desorption ionization-time of flight mass spectrometry for identification of Vibrio cholerae. J. Clin. Microbiol. 2015, 53, 329–331. [Google Scholar] [CrossRef] [Green Version]
- Tarr, C.L.; Patel, J.S.; Puhr, N.D.; Sowers, E.G.; Bopp, C.A.; Strockbine, N.A. Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. J. Clin. Microbiol. 2007, 45, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.; Huq, A.; Colwell, R.R. Analysis of 16S–23S rRNA intergenic spacer regions of Vibrio cholerae and Vibrio mimicus. Appl. Environ. Microbiol. 1999, 65, 2202–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, F.L.; Gevers, D.; Thompson, C.C.; Dawyndt, P.; Naser, S.; Hoste, B.; Munn, C.B.; Swings, J. Phylogeny and molecular identification of vibrios on the basis of multilocus sequence analysis. Appl. Environ. Microbiol. 2005, 71, 5107–5115. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.C.; Thompson, F.L.; Vicente, A.C.P.; Swings, J. Phylogenetic analysis of vibrios and related species by means of atpA gene sequences. Int. J. Syst. Evol. Microbiol. 2007, 57, 2480–2484. [Google Scholar] [CrossRef] [PubMed]
- Sohn, H.; Kim, J.; Jin, C.; Lee, J. Identification of Vibrio species isolated from cultured olive flounder (Paralichthys olivaceus) in Jeju Island, South Korea. Fish. Aquat. Sci. 2019, 22, 14. [Google Scholar] [CrossRef]
- Nuttall, R.; Sharma, G.; Moisander, P.H. Draft Genome Sequence of Vibrio cyclitrophicus NCT10V, Cultivated from the Microbiome of a Marine Copepod. Microbiol. Resour. Announc. 2019, 8, e01208-19. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.F.; Chen, Y.W.; Xu, J.K.; Ding, W.Y.; Shao, A.Q.; Zhu, Y.T.; Yang, J.L. Temperature elevation and Vibrio cyclitrophicus infection reduce the diversity of haemolymph microbiome of the mussel Mytilus coruscus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Hedlund, B.P.; Staley, J.T. Vibrio cyclotrophicus sp. nov., a polycyclic aromatic hydrocarbon (PAH)-degrading marine bacterium. Int. J. Syst. Evol. Microbiol. 2001, 51, 61–66. [Google Scholar] [CrossRef]
- Faury, N.; Saulnier, D.; Thompson, F.L.; Gay, M.; Swings, J.; Roux, F.L. Vibrio crassostreae sp. nov., isolated from the haemolymph of oysters (Crassostrea gigas). Int. J. Syst. Evol. Microbiol. 2004, 54, 2137–2140. [Google Scholar] [CrossRef] [Green Version]
- Petton, B.; Boudry, P.; Alunno-Bruscia, M.; Pernet, F. Factors influencing disease-induced mortality of Pacific oysters Crassostrea gigas. Aquac. Environ. Interact. 2015, 6, 205–222. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.W. Tractable models for testing theories about natural strategies: Foraging behaviour and habitat selection of free-ranging sharks. J. Fish Biol. 2003, 63, 53–73. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | Isolates No. | Matrix | Medium |
|---|---|---|---|
| 1 | A | Brain | TCBS |
| B | Brain | TSA2 | |
| 2 | Blood | BA | |
| 3 | A | Kidney | TSA2 |
| B | Brain | TSA2 | |
| 4 | A | Blood | BA |
| B | Brain | TCBS | |
| C | Brain | TCBS | |
| 6 | Kidney | TSA2 | |
| 8 | A | Kidney | TCBS |
| B | Kidney | TCBS | |
| 9 | A | Blood | TSA2 |
| B | Blood | TSA2 | |
| C | Brain | TCBS | |
| 11 | Brain | TCBS | |
| 13 | A | Kidney | TSA2 |
| B | Blood | TSA2 | |
| C | Brain | TSA2 | |
| 14 | A | Blood | TSA2 |
| B | Blood | TSA2 | |
| 19 | A | Kidney | TSA2 |
| B | Brain | BA | |
| 20 | A | Kidney | TSA2 |
| B | Blood | TSA2 |
| Isolates No. | Genetic ID | ID (%) | Reference for Identity | GenBank Accession |
|---|---|---|---|---|
| 1A | V. crassostreae | 98.71 | CP016228 | OM158222 |
| 1B | V. crassostreae | 98.85 | CP016228 | OM158223 |
| 2 | V. crassostreae | 98.69 | CP016228 | OM158216 |
| 3A | V. cyclitrophicus | 99.86 | CP039700 | OM158211 |
| 3B | V. cyclitrophicus | 100 | CP039700 | not submitted * |
| 4A | V. cyclitrophicus | 99.85 | CP039700 | OM158217 |
| 4B | V. crassostreae | 98.66 | CP016228 | OM158224 |
| 4C | V. cyclitrophicus | 99.86 | CP039700 | OM158225 |
| 6 | V. crassostreae | 98.8 | CP016228 | OM158212 |
| 8A | V. crassostreae | 98.81 | CP016228 | OM158213 |
| 8B | V. crassostreae | 98.82 | CP016228 | OM158214 |
| 9A | V. crassostreae | 98.71 | CP016228 | OM158218 |
| 9B | V. cyclitrophicus | 100 | CP039700 | not submitted * |
| 9C | V. crassostreae | 98.71 | CP016228 | OM158226 |
| 11 | V. crassostreae | 98.71 | CP016228 | OM158227 |
| 13A | V. cyclitrophicus | 100 | CP039700 | not submitted * |
| 13B | V. cyclitrophicus | 100 | CP039700 | not submitted * |
| 13C | V. cyclitrophicus | 99.86 | CP039700 | OM158228 |
| 14A | V. crassostreae | 99.2 | CP016228 | OM158219 |
| 14B | V. crassostreae | 98.72 | CP016228 | OM158220 |
| 19A | V. cyclitrophicus | 100 | CP039700 | not submitted * |
| 19B | V. crassostreae | 98.72 | CP016228 | OM158229 |
| 20A | V. cyclitrophicus | 99.86 | CP039700 | OM158215 |
| 20B | V. cyclitrophicus | 99.86 | CP039700 | OM158221 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomasoni, M.; Esposito, G.; Mugetti, D.; Pastorino, P.; Stoppani, N.; Menconi, V.; Gagliardi, F.; Corrias, I.; Pira, A.; Acutis, P.L.; et al. The Isolation of Vibrio crassostreae and V. cyclitrophicus in Lesser-Spotted Dogfish (Scyliorhinus canicula) Juveniles Reared in a Public Aquarium. J. Mar. Sci. Eng. 2022, 10, 114. https://doi.org/10.3390/jmse10010114
Tomasoni M, Esposito G, Mugetti D, Pastorino P, Stoppani N, Menconi V, Gagliardi F, Corrias I, Pira A, Acutis PL, et al. The Isolation of Vibrio crassostreae and V. cyclitrophicus in Lesser-Spotted Dogfish (Scyliorhinus canicula) Juveniles Reared in a Public Aquarium. Journal of Marine Science and Engineering. 2022; 10(1):114. https://doi.org/10.3390/jmse10010114
Chicago/Turabian StyleTomasoni, Mattia, Giuseppe Esposito, Davide Mugetti, Paolo Pastorino, Nadia Stoppani, Vasco Menconi, Flavio Gagliardi, Ilaria Corrias, Angela Pira, Pier Luigi Acutis, and et al. 2022. "The Isolation of Vibrio crassostreae and V. cyclitrophicus in Lesser-Spotted Dogfish (Scyliorhinus canicula) Juveniles Reared in a Public Aquarium" Journal of Marine Science and Engineering 10, no. 1: 114. https://doi.org/10.3390/jmse10010114