Differential Expression of Circulating Plasma miRNA-370 and miRNA-10a from Patients with Hereditary Hemorrhagic Telangiectasia



Abstract
:1. Introduction
2. Material and Methods
2.1. Algorithms for miRNA-target Predictions
2.2. Patients
2.3. Extraction of miRNAs from Plasma Samples
2.4. Quantitative Reverse Transcriptase–Polymerase Chain Reaction (qRT-PCR) Analysis of miRNAs
2.5. Statistical Analysis
3. Results
3.1. Identification of miRNA-10a, miRNA-214, and miRNA-370, Potentially Related to HHT, Using in Silico and Literature Data
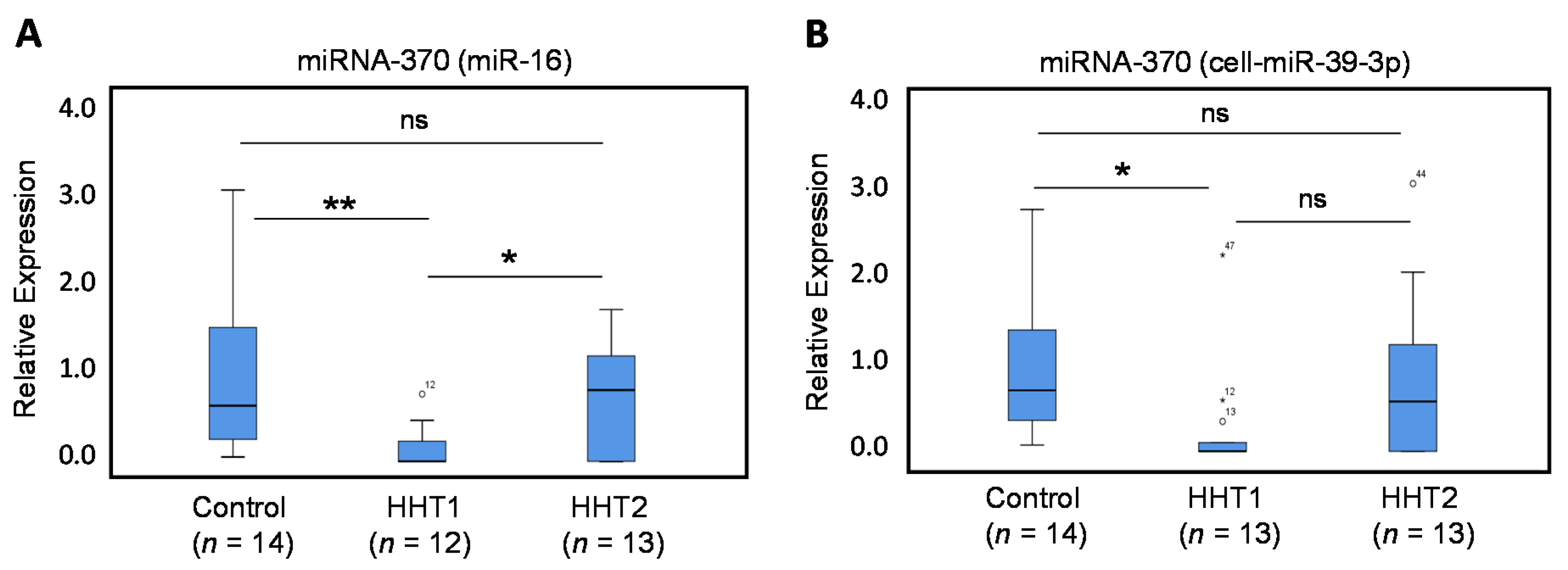
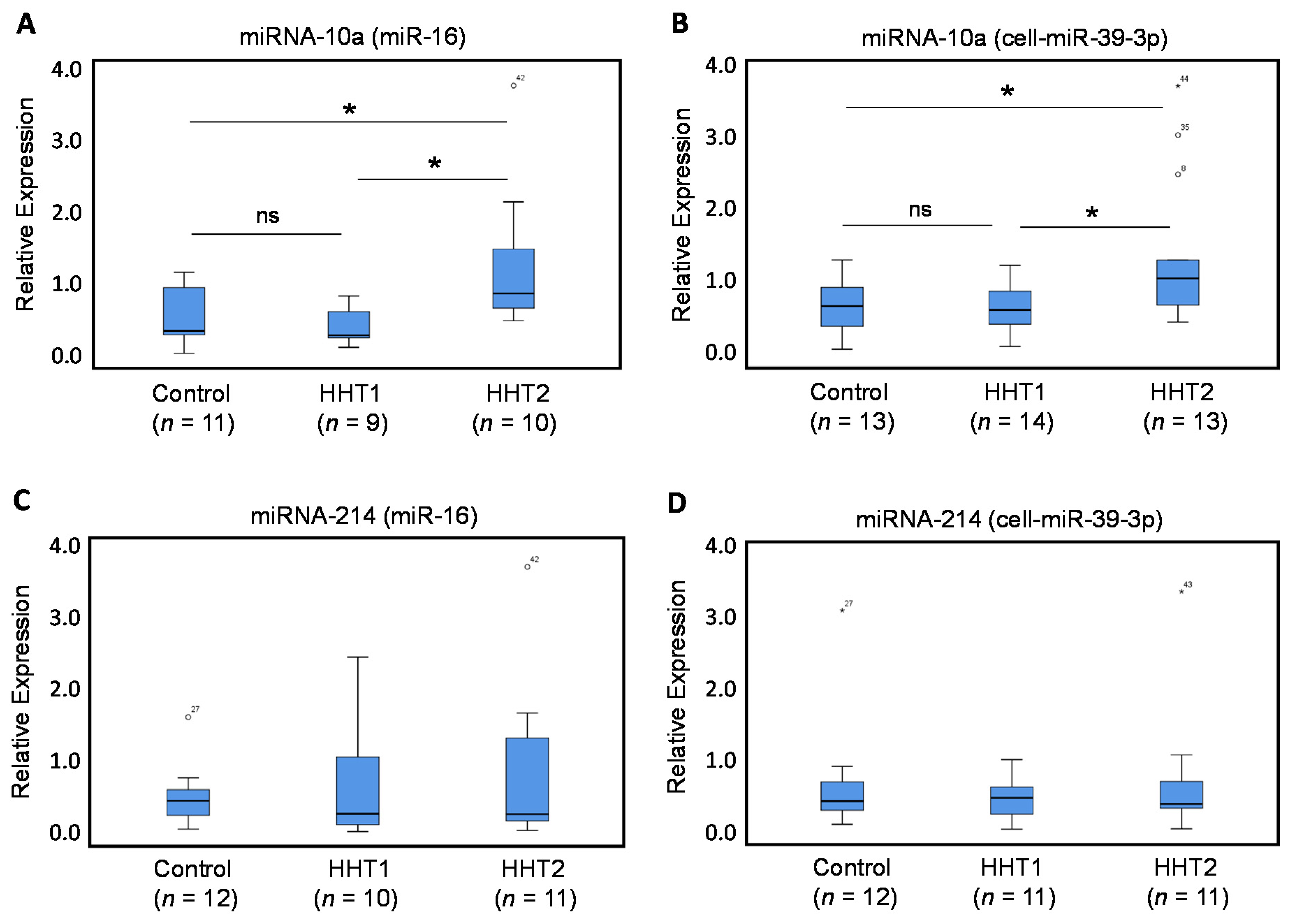
3.2. Circulating Levels of miRNA-370, miRNA-10a, and miRNA-214 in HHT1 and HHT2 Patient Plasma
3.2.1. Circulating Levels of miRNA-370 are Decreased in HHT1 Patient Plasma
3.2.2. Circulating Levels of miRNA-10a, but not of miRNA-214, are Dysregulated in HHT2 Patient Plasma
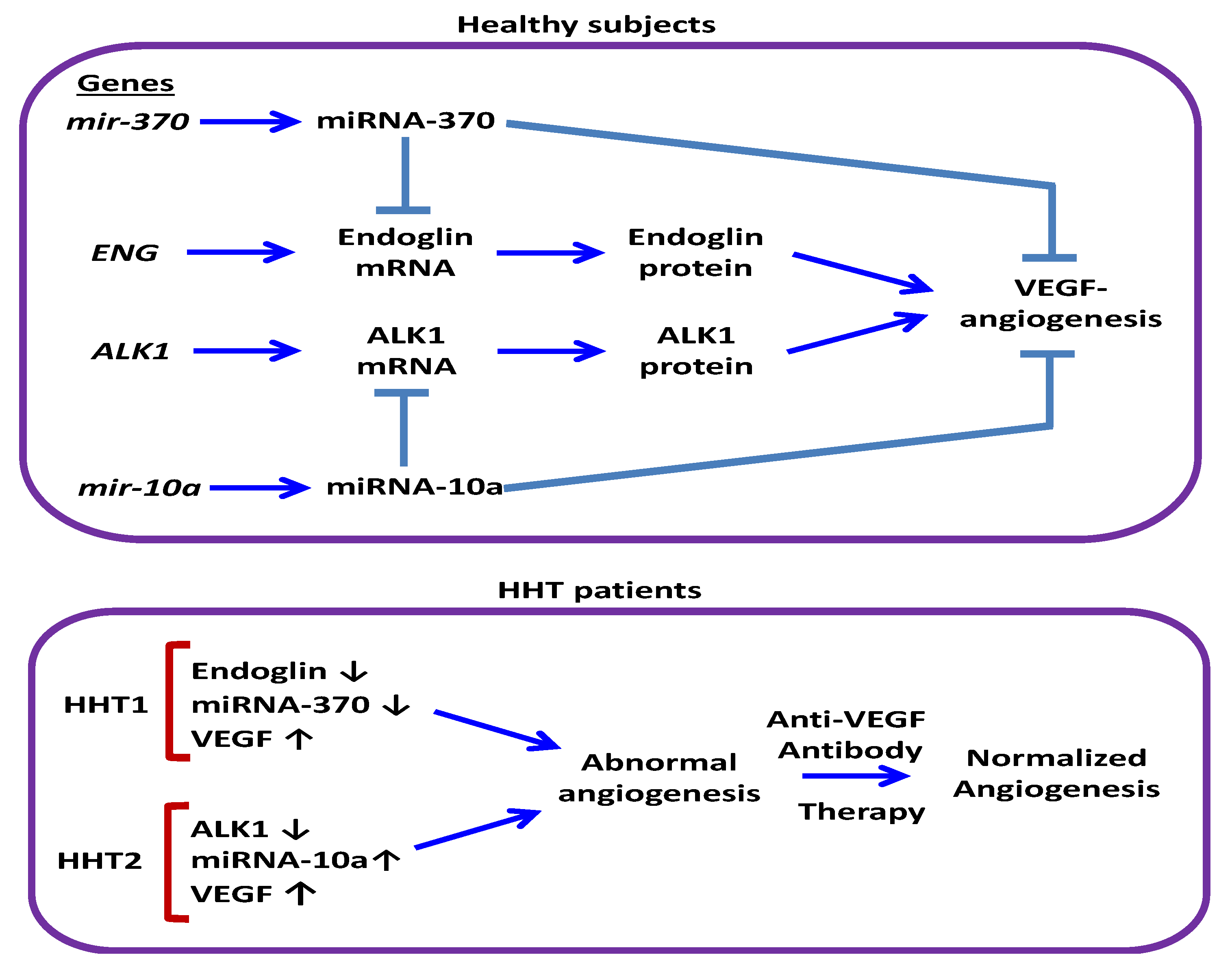
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shovlin, C.L. Hereditary haemorrhagic telangiectasia: Pathophysiology, diagnosis and treatment. Blood Rev. 2010, 24, 203–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, J.; Bayrak-Toydemir, P.; Pyeritz, R.E. Hereditary hemorrhagic telangiectasia: An overview of diagnosis, management, and pathogenesis. Genet. Med. 2011, 13, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shovlin, C.L.; Guttmacher, A.E.; Buscarini, E.; Faughnan, M.E.; Hyland, R.H.; Westermann, C.J.J.; Kjeldsen, A.D.; Plauchu, H. Diagnostic criteria for Hereditary Hemorrhagic Telangiectasia (Rendu- Osler-Weber Syndrome). Am. J. Med. Genet. 2000, 91, 66–67. [Google Scholar] [CrossRef]
- Shovlin, C.L.; Buscarini, E.; Kjeldsen, A.D.; Mager, H.J.; Sabba, C.; Droege, F.; Geisthoff, U.; Ugolini, S.; Dupuis-Girod, S. European Reference Network for Rare Vascular Diseases (VASCERN) Outcome Measures for Hereditary Haemorrhagic Telangiectasia (HHT). Orphanet. J. Rare. Dis. 2018, 13, 136. [Google Scholar] [CrossRef]
- Faughnan, M.E.; Palda, V.A.; Garcia-Tsao, G.; Geisthoff, U.W.; McDonald, J.; Proctor, D.D.; Spears, J.; Brown, D.H.; Buscarini, E.; Chesnutt, M.S.; et al. International guidelines for the diagnosis and management of hereditary haemorrhagic telangiectasia. J. Med. Genet. 2011, 48, 73–87. [Google Scholar] [CrossRef]
- McAllister, K.A.; Grogg, K.M.; Johnson, D.W.; Gallione, C.J.; Baldwin, M.A.; Jackson, C.E.; Helmbold, E.A.; Markel, D.S.; McKinnon, W.C.; Murrel, J.; et al. Endoglin, a TGF-β binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat. Genet. 1994, 8, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.W.; Berg, J.N.; Baldwin, M.A.; Gallione, C.J.; Marondel, I.; Yoon, S.J.; Stenzel, T.T.; Speer, M.; Pericak-Vance, M.A.; Diamond, A.; et al. Mutations in the activin receptor-like kinase 1 gene in hereditary haemorrhagic telangiectasia type. Nat. Genet. 1996, 13, 189–195. [Google Scholar] [CrossRef]
- Gallione, C.J.; Repetto, G.M.; Legius, E.; Rustgi, A.K.; Schelley, S.L.; Tejpar, S.; Mitchell, G.; Drouin, É.; Westermann, C.J.J.; Marchuk, D.A. A combined syndrome of juvenile polyposis and hereditary haemorrhagic telangiectasia associated with mutations in MADH4 (SMAD4). Lancet 2004, 363, 852–859. [Google Scholar] [CrossRef]
- Wooderchak-Donahue, W.L.; McDonald, J.; O’Fallon, B.; Upton, P.D.; Li, W.; Roman, B.L.; Young, S.; Plant, P.; Fülöp, G.T.; Langa, C.; et al. BMP9 mutations cause a vascular-anomaly syndrome with phenotypic overlap with hereditary hemorrhagic telangiectasia. Am. J. Hum. Genet. 2013, 93, 530–537. [Google Scholar] [CrossRef] [Green Version]
- McDonald, J.; Bayrak-Toydemir, P.; DeMille, D.; Wooderchak-Donahue, W.; Whitehead, K. Curaçao diagnostic criteria for hereditary hemorrhagic telangiectasia is highly predictive of a pathogenic variant in ENG or ACVRL1 (HHT1 and HHT2). Genet. Med. 2020. [Google Scholar] [CrossRef]
- Abdalla, S.A.; Letarte, M. Hereditary haemorrhagic telangiectasia: Current views on genetics and mechanisms of disease. J. Med. Genet. 2006, 43, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Llorente, L.; Gallardo-Vara, E.; Rossi, E.; Smadja, D.M.; Botella, L.M.; Bernabeu, C. Endoglin and alk1 as therapeutic targets for hereditary hemorrhagic telangiectasia. Expert Opin. Ther. Targets 2017, 21, 933–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, B.L.; Hinck, A.P. ALK1 signaling in development and disease: New paradigms. Cell. Mol. Life Sci. 2017, 74, 4539–4560. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.H.; Guo, J.; Morrell, N.W.; Li, W. Advances in the molecular regulation of endothelial BMP9 signalling complexes and implications for cardiovascular disease. Biochem. Soc. Trans. 2019, 47, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Park, S.O.; Wankhede, M.; Lee, Y.J.; Choi, E.-J.; Fliess, N.; Choe, S.-W.; Oh, S.-H.; Walter, G.; Raizada, M.; Sorg, B.; et al. Real-time imaging of de novo arteriovenous malformation in a mouse model of hereditary hemorrhagic telangiectasia. J. Clin. Investig. 2009, 119, 3487–3496. [Google Scholar] [CrossRef]
- Choi, E.J.; Chen, W.; Jun, K.; Arthur, H.M.; Young, W.L.; Su, H. Novel brain arteriovenous malformation mouse models for type 1 hereditary hemorrhagic telangiectasia. PLoS ONE 2014, 9, e88511. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Martin, E.M.; Nguyen, H.L.; Cunningham, T.A.; Choe, S.W.; Jiang, Z.; Arthur, H.M.; Lee, Y.J.; Oh, S.P. Common and distinctive pathogenetic features of arteriovenous malformations in hereditary hemorrhagic telangiectasia 1 and hereditary hemorrhagic telangiectasia 2 animal models—Brief report. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2232–2236. [Google Scholar] [CrossRef] [Green Version]
- Snellings, D.A.; Gallione, C.J.; Clark, D.S.; Vozoris, N.T.; Faughnan, M.E.; Marchuk, D.A. Somatic Mutations in Vascular Malformations of Hereditary Hemorrhagic Telangiectasia Result in Bi-allelic Loss of ENG or ACVRL1. Am. J. Hum. Genet. 2019, 105, 894–906. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Hata, A.; Lagna, G. Deregulation of Drosha in the pathogenesis of hereditary hemorrhagic telangiectasia. Curr. Opin. Hematol. 2019, 26, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Wooderchak-Donahue, W.L.; McDonald, J.; Ghatpande, P.; Baalbaki, M.; Sandoval, M.; Hart, D.; Clay, H.; Coughlin, S.; Lagna, G.; et al. Inactivating mutations in drosha mediate vascular abnormalities similar to hereditary hemorrhagic telangiectasia. Sci. Signal. 2018, 11, eaan6831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, H.; Suzuki, H.I. Systems and synthetic microRNA biology: From biogenesis to disease pathogenesis. Int. J. Mol. Sci. 2020, 21, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabruyn, S.P.; Hansen, S.; Ojeda-Fernández, M.L.; Bovy, N.; Zarrabeitia, R.; Recio-Poveda, L.; Bernabéu, C.; Martial, J.A.; Botella, L.M.; Struman, I. MiR-205 is downregulated in hereditary hemorrhagic telangiectasia and impairs TGF-beta signaling pathways in endothelial cells. Angiogenesis 2013, 16, 877–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Kandic, I.; Faughnan, M.E.; Kutryk, M.J. Elevated circulating microRNA-210 levels in patients with hereditary hemorrhagic telangiectasia and pulmonary arteriovenous malformations: A potential new biomarker. Biomarkers 2013, 18, 23–29. [Google Scholar] [CrossRef]
- Cannavicci, A.; Zhang, Q.; Dai, S.C.; Faughnan, M.E.; Kutryk, M.J.B. Decreased levels of miR-28-5p and miR-361-3p and increased levels of insulin-like growth factor 1 mRNA in mononuclear cells from patients with hereditary hemorrhagic telangiectasia. Can. J. Physiol. Pharmacol. 2019, 97, 562–569. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Hui, J.H.L.; Marco, A.; Ronshaugen, M. MicroRNA evolution by arm switching. EMBO Rep. 2011, 12, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, X.; Yuan, J.; Wu, J.; Deng, X.; Peng, J.; Wang, S.; Yang, C.; Ge, J.; Zou, Y. Evaluation of the performance of serum miRNAs as normalizers in microRNA studies focused on cardiovascular disease. J. Thorac. Dis. 2018, 10, 2599–2607. [Google Scholar] [CrossRef] [PubMed]
- Gareev, I.; Beylerli, O.; Yang, G.; Sun, J.; Pavlov, V.; Izmailov, A.; Shi, H.; Zhao, S. The current state of MiRNAs as biomarkers and therapeutic tools. Clin. Exp. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.P.; Chen, Y.G.; Lan, J.Y.; Shen, Z.J. MicroRNA-370 suppresses proliferation and promotes endometrioid ovarian cancer chemosensitivity to cDDP by negatively regulating ENG. Cancer Lett. 2014, 353, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, G.; Yu, C.; Shen, Z.; Xu, C.; Feng, Z.; Zhang, X.; Li, Y. A role of microRNA-370 in hepatic ischaemia-reperfusion injury by targeting transforming growth factor-β receptor II. Liver Int. 2015, 35, 1124–1132. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, X.; Hao, D. Upregulation of microRNA-370 facilitates the repair of amputated fingers through targeting forkhead box protein O1. Exp. Biol. Med. 2016, 241, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Song, G.; Yuan, J.; Qiao, S.; Xu, S.; Si, Z.; Yang, Y.; Xu, X.; Wang, A. Circular RNA circ_0003204 inhibits proliferation, migration and tube formation of endothelial cell in atherosclerosis via miR-370-3p/TGFβR2/phosph-SMAD3 axis. J. Biomed. Sci. 2020, 27, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Chen, L. MicroRNA-370 suppresses the retinal capillary endothelial cell growth by targeting KDR gene. Bratislava Med. J. 2017, 118, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.Z.; Chen, X.L.; Wu, W.; Hang, C.H. MicroRNA-370-3p inhibits human vascular smooth muscle cell proliferation via targeting KDR/AKT signaling pathway in cerebral aneurysm. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1080–1087. [Google Scholar]
- Gu, Y.; Becker, V.; Zhao, Y.; Menger, M.D.; Laschke, M.W. miR-370 inhibits the angiogenic activity of endothelial cells by targeting smoothened (SMO) and bone morphogenetic protein (BMP)-2. FASEB J. 2019, 33, 7213–7224. [Google Scholar] [CrossRef]
- Zhao, Y.; Ponnusamy, M.; Zhang, L.; Zhang, Y.; Liu, C.; Yu, W.; Wang, K.; Li, P. The role of miR-214 in cardiovascular diseases. Eur. J. Pharmacol. 2017, 816, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Shih, T.C.; Tien, Y.J.; Wen, C.J.; Yeh, T.S.; Yu, M.C.; Huang, C.H.; Lee, Y.S.; Yen, T.C.; Hsieh, S.Y. MicroRNA-214 downregulation contributes to tumor angiogenesis by inducing secretion of the hepatoma-derived growth factor in human hepatoma. J. Hepatol. 2012, 57, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Van Mil, A.; Grundmann, S.; Goumans, M.J.; Lei, Z.; Oerlemans, M.I.; Jaksani, S.; Doevendans, P.A.; Sluijter, J.P.G. MicroRNA-214 inhibits angiogenesis by targeting Quaking and reducing angiogenic growth factor release. Cardiovasc. Res. 2012, 93, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, Y.; Zhao, Q.; Wang, J.; He, X. MicroRNA-10a influences osteoblast differentiation and angiogenesis by regulating β-catenin expression. Cell. Physiol. Biochem. 2015, 37, 2194–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ling, C.C.; Li, L.; Qin, Y.; Qi, J.; Liu, X.; You, B.; Shi, Y.; Zhang, J.; Jiang, Q.; et al. MicroRNA-10a/10b represses a novel target gene mib1 to regulate angiogenesis. Cardiovasc. Res. 2016, 110, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Lin, T.E.; Lee, C.I.; Zhou, J.; Huang, Y.H.; Lee, P.L.; Shih, Y.T.; Chien, S.; Chiu, J.J. MicroRNA-10a is crucial for endothelial response to different flow patterns via interaction of retinoid acid receptors and histone deacetylases. Proc. Natl. Acad. Sci. USA 2017, 114, 2072–2077. [Google Scholar] [CrossRef] [Green Version]
- Navickas, R.; Gal, D.; Laucevičius, A.; Taparauskaite, A.; Zdanyte, M.; Holvoet, P. Identifying circulating microRNAs as biomarkers of cardiovascular disease: A systematic review. Cardiovasc. Res. 2016, 111, 322–337. [Google Scholar] [CrossRef]
- Botella, L.M.; Albiñana, V.; Ojeda-Fernandez, L.; Recio-Poveda, L.; Bernabéu, C. Research on potential biomarkers in hereditary hemorrhagic telangiectasia. Front. Genet. 2015, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- McDonald, J.; Wooderchak-Donahue, W.; VanSant Webb, C.; Whitehead, K.; Stevenson, D.A.; Bayrak-Toydemir, P. Hereditary hemorrhagic telangiectasia: Genetics and molecular diagnostics in a new era. Front. Genet. 2015, 6, 1. [Google Scholar] [CrossRef]
- Majumdar, S.; McWilliams, J.P. Approach to Pulmonary Arteriovenous Malformations: A Comprehensive Update. J. Clin. Med. 2020, 9, 1927. [Google Scholar] [CrossRef]
- Aronson, J.K.; Ferner, R.E. Biomarkers-A General Review. Curr. Protoc. Pharmacol. 2017, 76, 9.23.1–9.23.17. [Google Scholar] [CrossRef] [PubMed]
- Califf, R.M. Biomarker definitions and their applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour, S.; He, Y.; Nanda, A.; Ye, Q. MicroRNAs Involved in the Regulation of Angiogenesis in Bone Regeneration. Calcif. Tissue. Int. 2019, 105, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Choe, S.W.; Kim, Y.H.; Acharya, A.P.; Keselowsky, B.G.; Sorg, B.S.; Lee, Y.J.; Oh, S.P. VEGF neutralization can prevent and normalize arteriovenous malformations in an animal model for hereditary hemorrhagic telangiectasia 2. Angiogenesis 2014, 17, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Albiñana, V.; Cuesta, A.M.; de Rojas-P., I.; Gallardo-Vara, E.; Recio-Poveda, L.; Bernabéu, C.; Botella, L.M. Review of Pharmacological Strategies with Repurposed Drugs for Hereditary Hemorrhagic Telangiectasia Related Bleeding. J. Clin. Med. 2020, 9, 1766. [Google Scholar]
- Cirulli, A.; Liso, A.; D’Ovidio, F.; Mestice, A.; Pasculli, G.; Gallitelli, M.; Rizzi, R.; Specchia, G.; Sabbà, C. Vascular endothelial growth factor serum levels are elevated in patients with hereditary hemorrhagic telangiectasia. Acta Haematol. 2003, 110, 29–32. [Google Scholar] [CrossRef]
- Sadick, H.; Riedel, F.; Naim, R.; Goessler, U.; Hörmann, K.; Hafner, M.; Lux, A. Patients with hereditary hemorrhagic telangiectasia have increased plasma levels of vascular endothelial growth factor and transforming growth factor-β1 as well as high ALK1 tissue expression. Haematologica 2005, 90, 818–828. [Google Scholar]
- Buscarini, E.; Botella, L.M.; Geisthoff, U.; Kjeldsen, A.D.; Mager, H.J.; Pagella, F.; Suppressa, P.; Zarrabeitia, R.; Dupuis-Girod, S.; Shovlin, C.L.; et al. Safety of thalidomide and bevacizumab in patients with hereditary hemorrhagic telangiectasia. Orphanet J. Rare Dis. 2019, 14, 28. [Google Scholar] [CrossRef] [Green Version]
- Shao, E.S.; Lin, L.; Yao, Y.; Boström, K.I. Expression of vascular endothelial growth factor is coordinately regulated by the activin-like kinase receptors 1 and 5 in endothelial cells. Blood 2009, 114, 2197–2206. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Huang, J.J.; Golzio, C.; Gao, X.; Hector-Greene, M.; Katsanis, N.; Blobe, G.C. Endoglin interacts with VEGFR2 to promote angiogenesis. FASEB J. 2018, 32, 2934–2949. [Google Scholar] [CrossRef] [Green Version]
- López-Novoa, J.M.; Bernabeu, C. The physiological role of endoglin in the cardiovascular system. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H959–H974. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| HHT Type | Patient# | Genotype | Mutation |
|---|---|---|---|
| HHT1 | P#1.1 | ENG | 5′ UTR (gene promoter) c.-127 G>A |
| HHT1 | P#1.2 | ENG | 5′ UTR (gene promoter) c.-127 G>A |
| HHT1 | P#1.3 | ENG | 5′ UTR (gene promoter) c.-58 G>A |
| HHT1 | P#1.4 | ENG | 5′ UTR (gene promoter) c.-58 G>A |
| HHT1 | P#1.5 | ENG | 5′ UTR (gene promoter) c.-58 G>C |
| HHT1 | P#1.6 | ENG | Intron 1 c.68-2A>T |
| HHT1 | P#1.7 | ENG | Intron 1 c.68-2A>T |
| HHT1 | P#1.8 | ENG | Intron 1 c.68-2A>T |
| HHT1 | P#1.9 | ENG | Exon 4 c.392 C>T; p.Pro131Leu |
| HHT1 | P#1.10 | ENG | Exon 5 c.588 G>A; p.Trp196 * |
| HHT1 | P#1.11 | ENG | Exon 5 c.588 G>A; p.Trp196 * |
| HHT1 | P#1.12 | ENG | Exon 5 c.617delG; p.G206AfsX16 |
| HHT1 | P#1.13 | ENG | Exon 5 c.617delG; p.G206AfsX16 |
| HHT1 | P#1.14 | ENG | Exon 7 c.967_968del GT; p.V323fs * |
| HHT1 | P#1.15 | ENG | Exon 7 c.967_968delGT; p.V323fs * |
| HHT1 | P#1.16 | ENG | Exon 11 c.1434_1435 del AG p. R478fs * |
| HHT2 | P#2.1 | ALK1 | Exon 6 c.673_674delAG; p.S225fs |
| HHT2 | P#2.2 | ALK1 | Exon 6 c.673_674delAG; p.S225fs |
| HHT2 | P#2.3 | ALK1 | Exon 6 c.635 G>A; p.R212H |
| HHT2 | P#2.4 | ALK1 | Exon 7 c.889delC; H297fs * |
| HHT2 | P#2.5 | ALK1 | Exon 7 c.921-927dupATGCGGC; p.L310fs |
| HHT2 | P#2.6 | ALK1 | Exon 7 c. 926 G>A; p.G309A |
| HHT2 | P#2.7 | ALK1 | Exon 7 c.941 A>C; p.His314Pro |
| HHT2 | P#2.8 | ALK1 | Exon 7 c.988 G>T; p.D330Y |
| HHT2 | P#2.9 | ALK1 | Exon 7 c.1027 C>T; p.Q374X |
| HHT2 | P#2.10 | ALK1 | Exon 7 c.1027 C>T; p.Q374X |
| HHT2 | P#2.11 | ALK1 | Exon 7 c.1027 C>T; p.Q374X |
| HHT2 | P#2.12 | ALK1 | Exon 7 c.1030 C>T; p.C344R |
| HHT2 | P#2.13 | ALK1 | Exon 8 c.1120 C>T; p.R374W |
| HHT2 | P#2.14 | ALK1 | Exon 8 c.1120 C>T; p.Arg374Trp |
| HHT2 | P#2.15 | ALK1 | Exon 8 c.1232 G>A; p.Arg411Gln |
| HHT2 | P#2.16 | ALK1 | Exon 10 c.1435 C>T; p.Arg479X |
| miRNA | HHT1 | HHT2 | HHT (Pool) | Blood Sample | Reference |
|---|---|---|---|---|---|
| miRNA-370 | ↓ | ↔ | ND | Plasma | Present work |
| miRNA-10a | ↔ | ↑ | ND | Plasma | Present work |
| miRNA-28-5p | ND | ND | ↓ | PMNCs | Cannavicci et al., 2019 |
| miRNA-361-3p | ND | ND | ↓ | PMNCs | Cannavicci et al., 2019 |
| miRNA-210 | ND | ND | ↑ | Plasma | Zhang et al., 2013 |
| miRNA-210 | ND | ND | (with PAVMs) | Plasma | Zhang et al., 2013 |
| miRNA-27a | ↑ | ↑ | ↔ | Plasma | Tabruyn et al., 2013 |
| miRNA-205 | ↓ | ↓ | (without PAVMs) | Plasma | Tabruyn et al., 2013 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Llorente, L.; Albiñana, V.; Botella, L.M.; Bernabeu, C. Differential Expression of Circulating Plasma miRNA-370 and miRNA-10a from Patients with Hereditary Hemorrhagic Telangiectasia. J. Clin. Med. 2020, 9, 2855. https://doi.org/10.3390/jcm9092855
Ruiz-Llorente L, Albiñana V, Botella LM, Bernabeu C. Differential Expression of Circulating Plasma miRNA-370 and miRNA-10a from Patients with Hereditary Hemorrhagic Telangiectasia. Journal of Clinical Medicine. 2020; 9(9):2855. https://doi.org/10.3390/jcm9092855
Chicago/Turabian StyleRuiz-Llorente, Lidia, Virginia Albiñana, Luisa M. Botella, and Carmelo Bernabeu. 2020. "Differential Expression of Circulating Plasma miRNA-370 and miRNA-10a from Patients with Hereditary Hemorrhagic Telangiectasia" Journal of Clinical Medicine 9, no. 9: 2855. https://doi.org/10.3390/jcm9092855