Amoebae in Chronic, Polymicrobial Endodontic Infections Are Associated with Altered Microbial Communities of Increased Virulence

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Correlation and Statistical Analysis
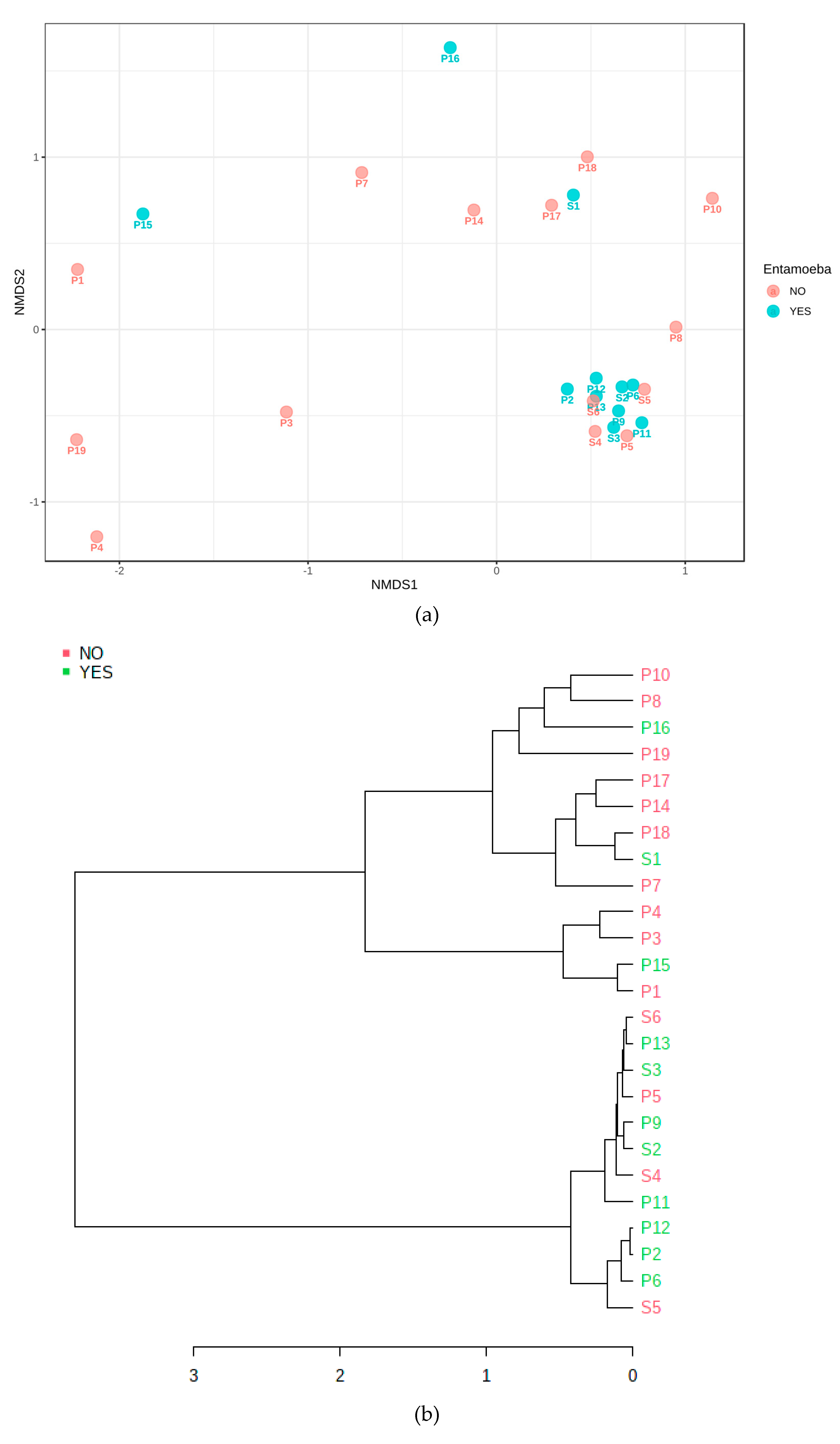
2.2.1. Community Clustering
2.2.2. Statistical Methods
3. Results
3.1. Patient Cohort
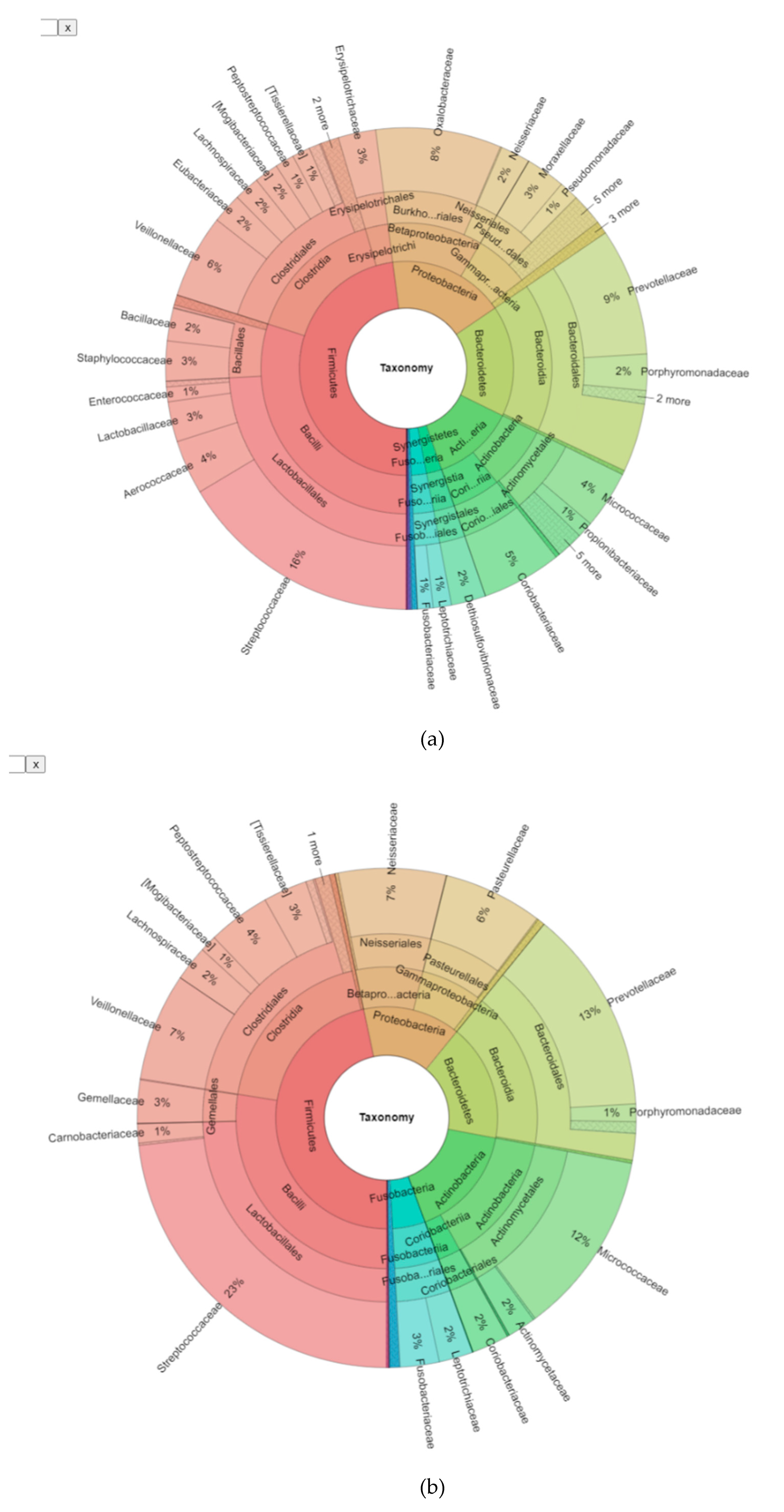
3.2. Bioinformatic Analysis of the Bacterial Microbiome in Endodontic Lesions
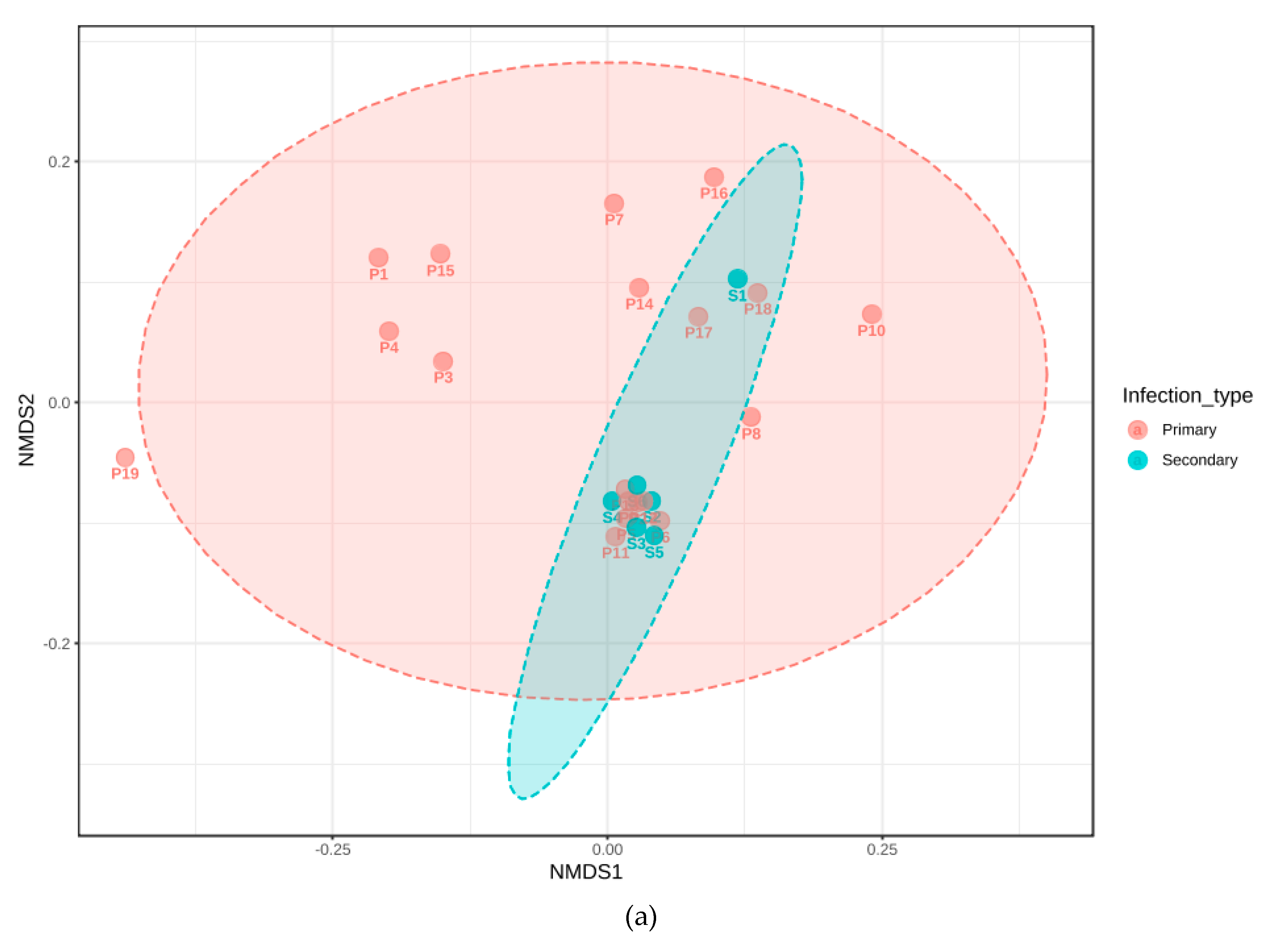
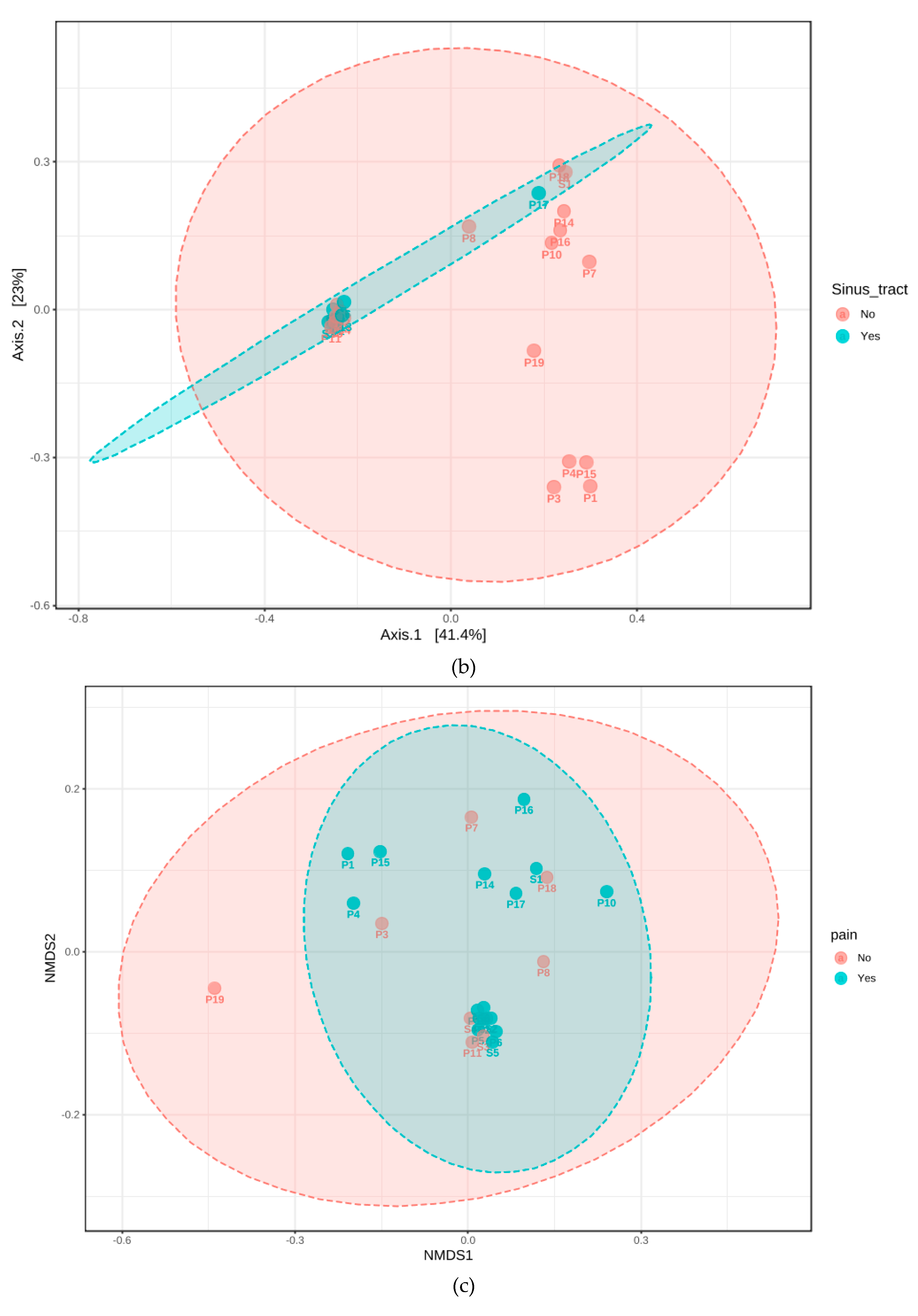
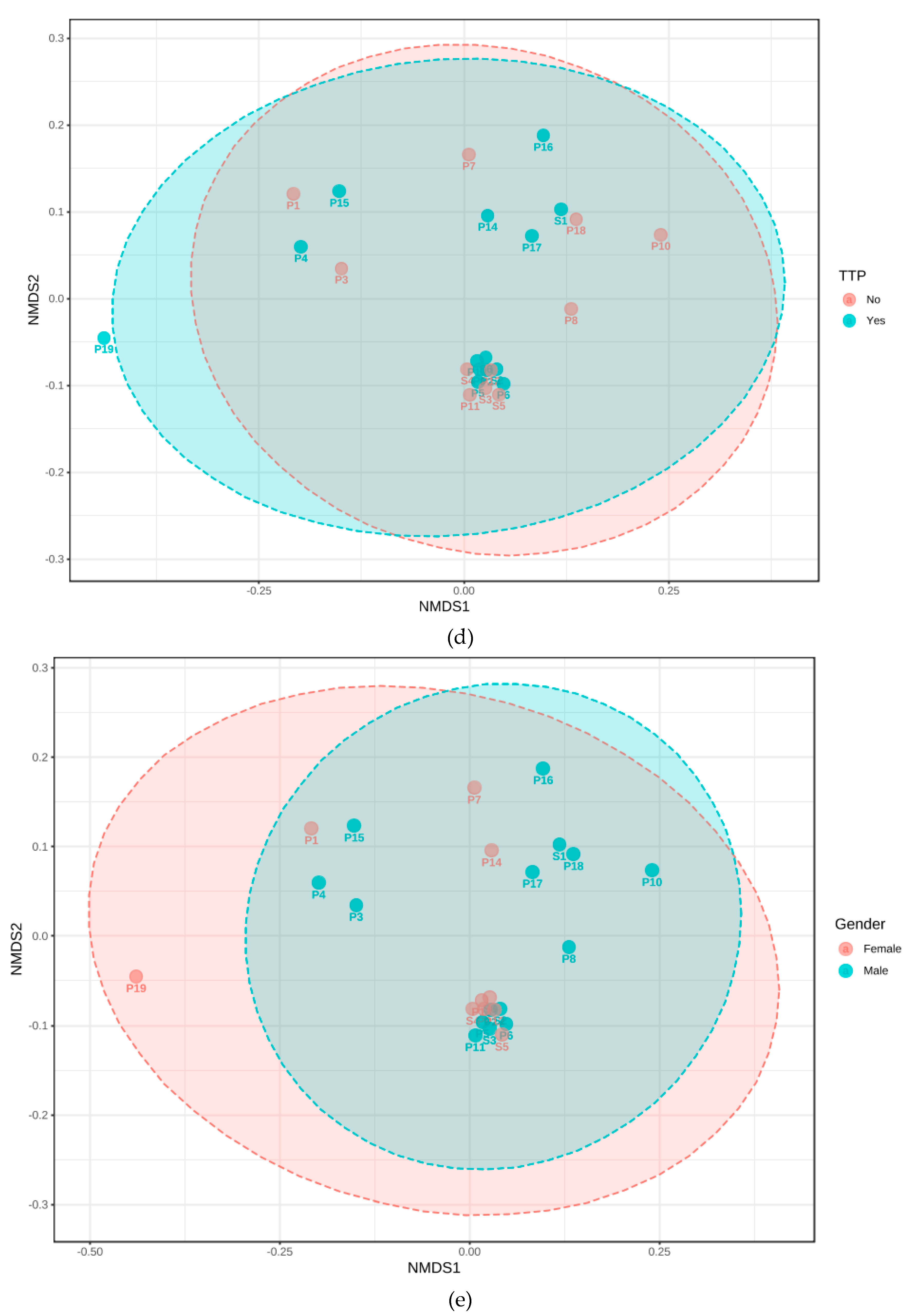
3.3. Phylogenetic Grouping of Endodontic Infections
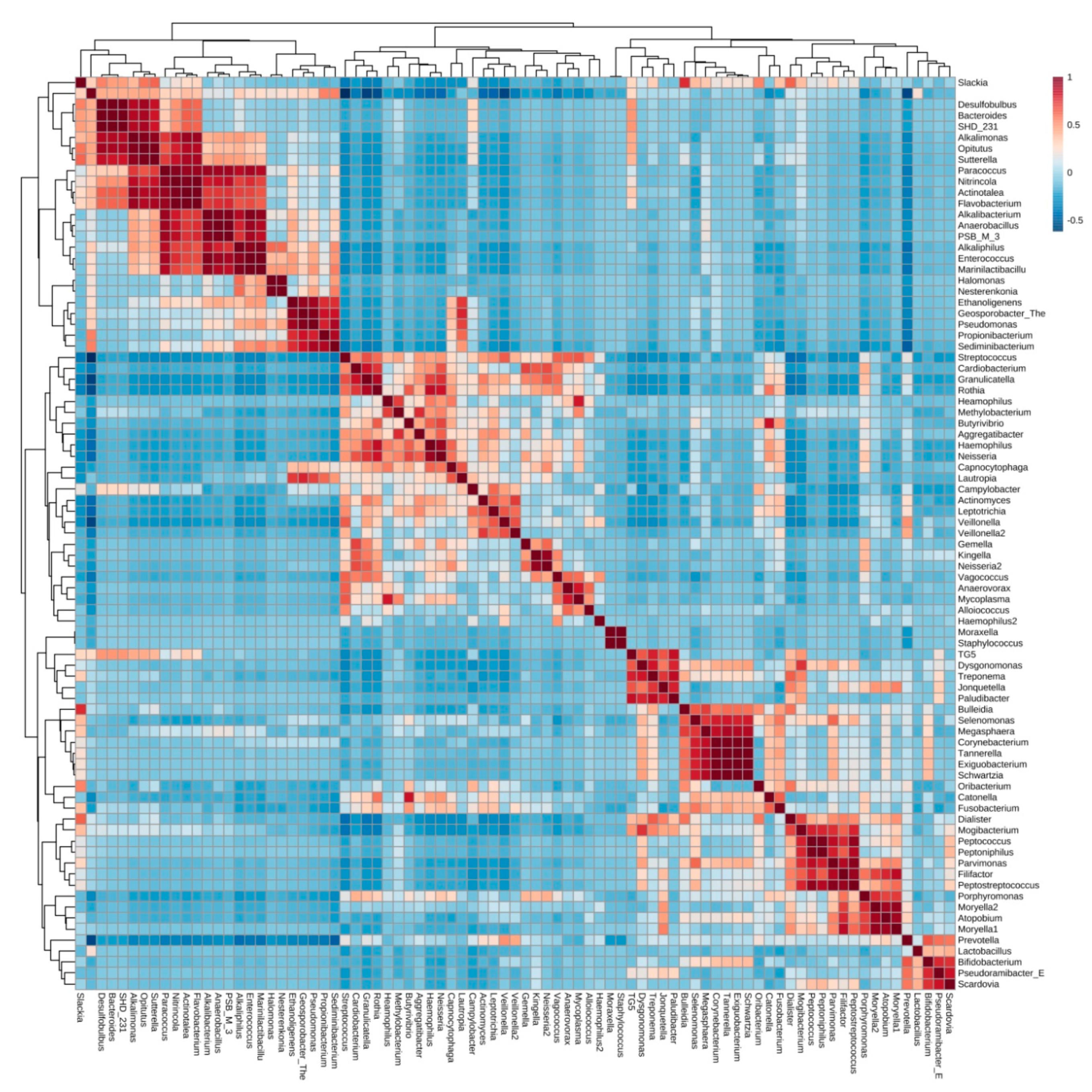
3.4. Cluster Analysis of Bacteria Present in Endodontic Lesions
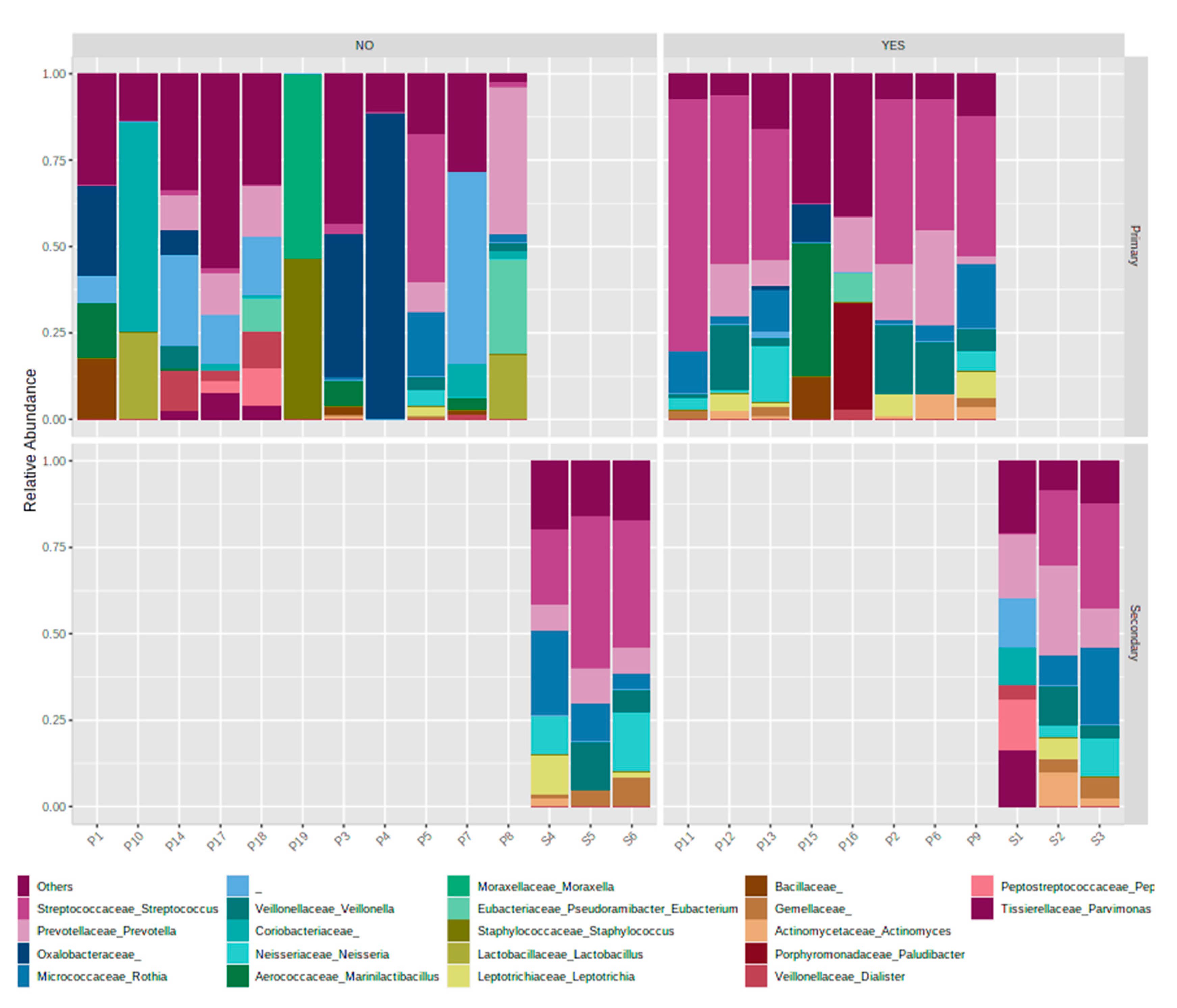
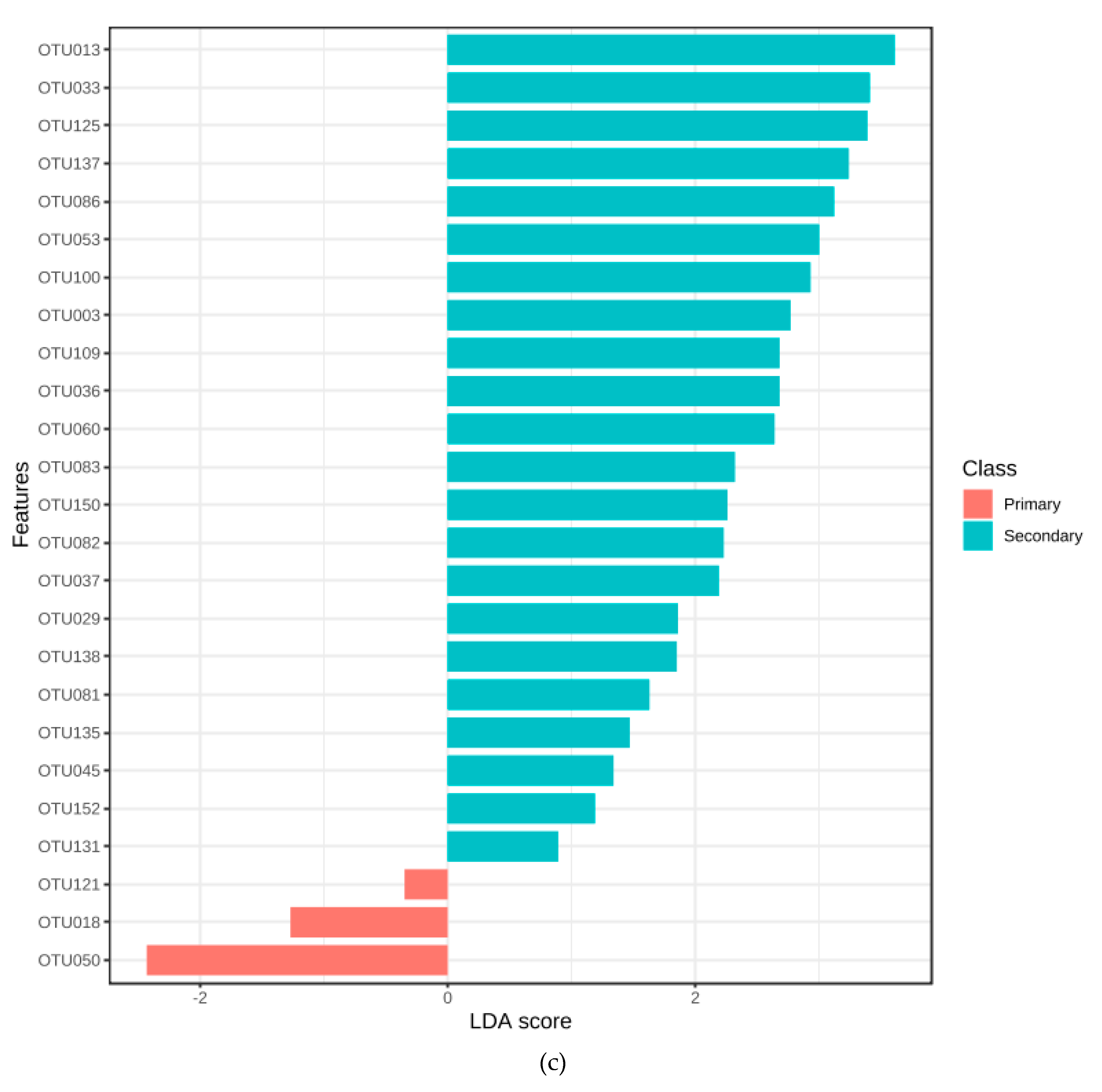
3.5. Association with Primary or Secondary Infections
3.6. Quantitative Polymerase Chain Reaction (qPCR)
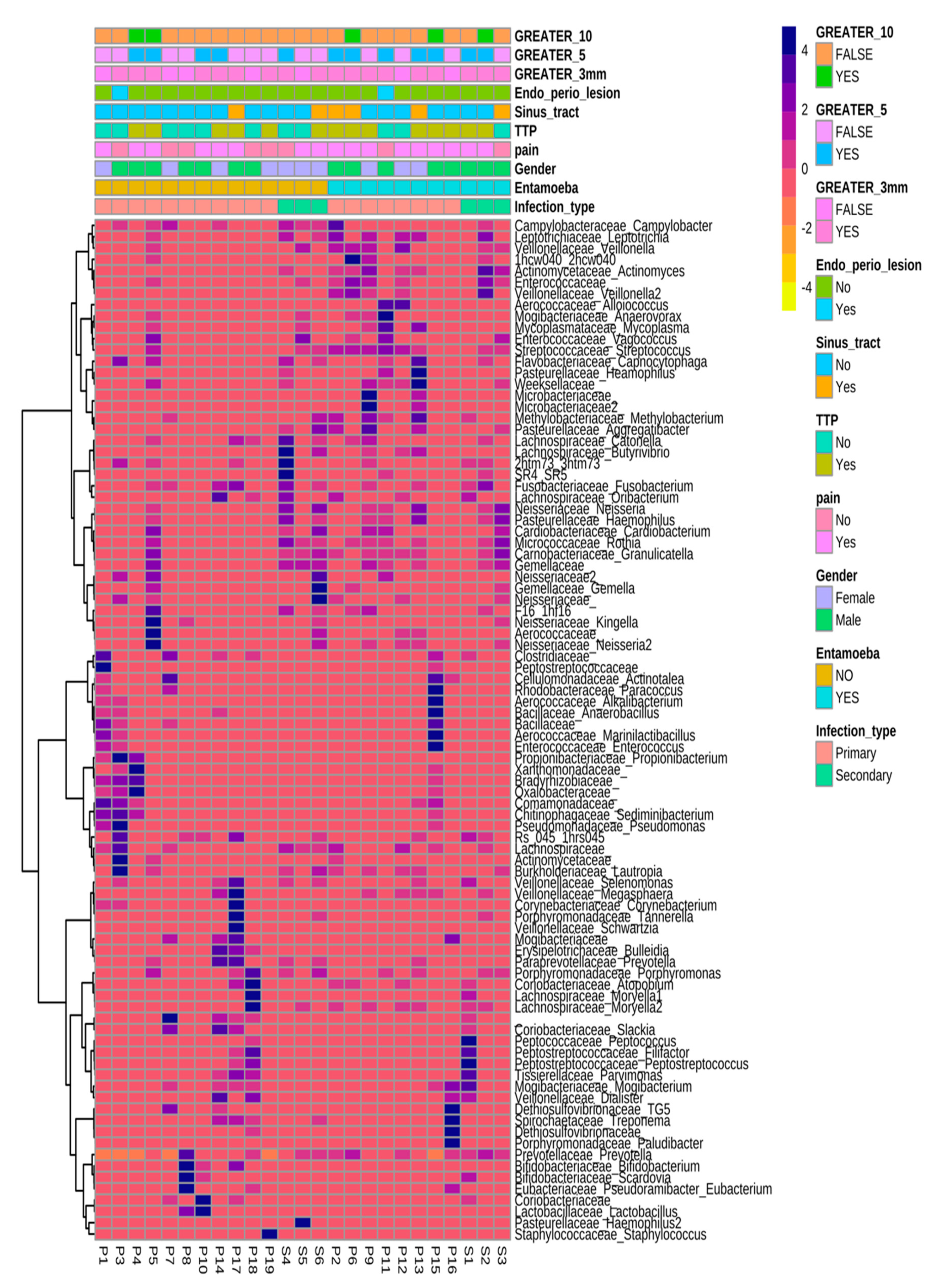
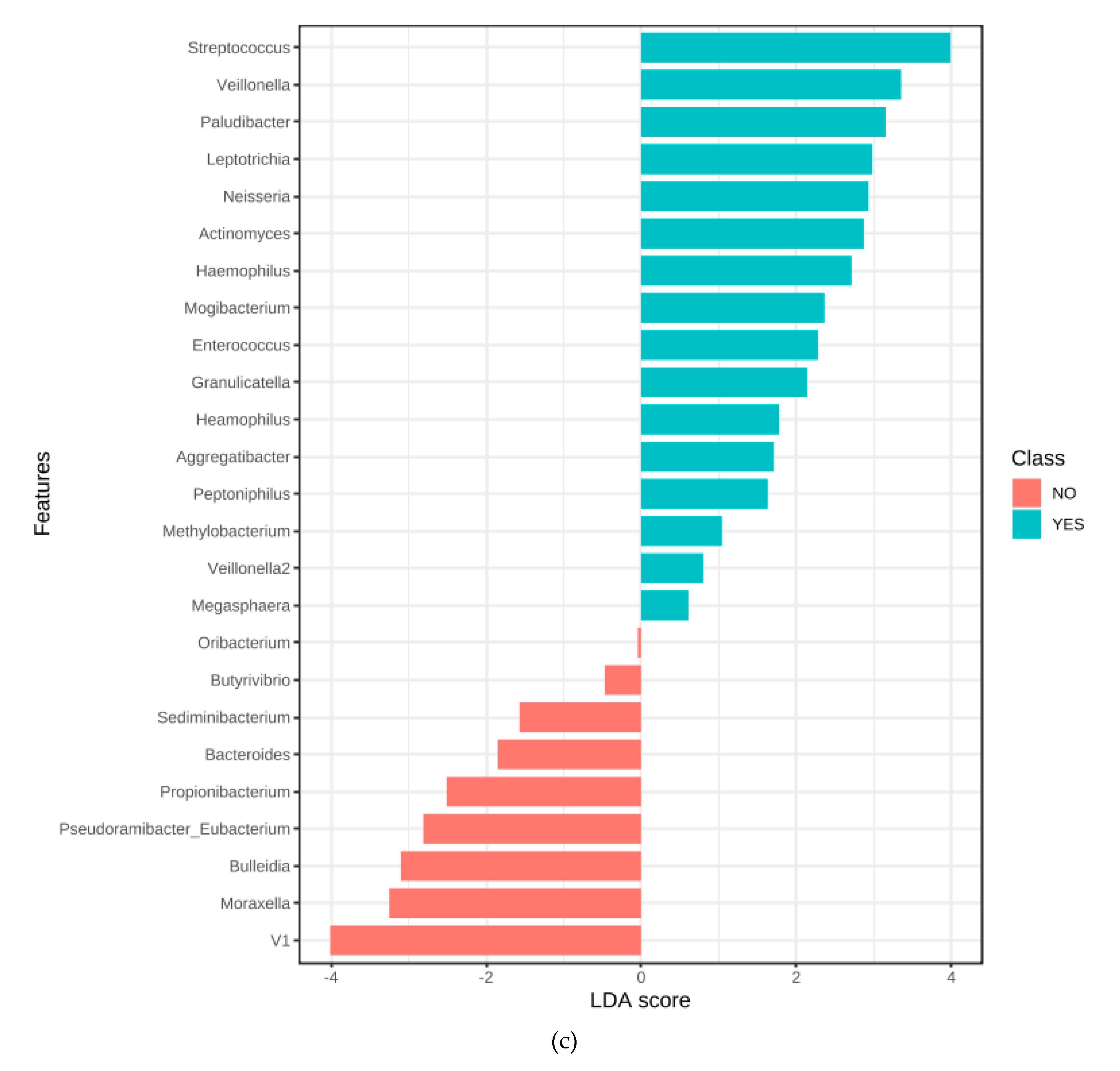
3.7. Microbiome Associated with Entamoeba Status
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siqueira, J.F., Jr.; Rocas, I.N. Diversity of endodontic microbiota revisited. J. Dent. Res. 2009, 88, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Paster, B.J.; Olsen, I.; Aas, J.A.; Dewhirst, F.E. The breadth of bacterial diversity in the human periodontal pocket and other oral sites. Periodontology 2000 2006, 42, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Al-Ahmad, A.; Elamin, F.; Jonas, D.; Mirghani, Y.; Schilhabel, M.; Karygianni, L.; Hellwig, E.; Rehman, A. Comparison of the bacterial composition and structure in symptomatic and asymptomatic endodontic infections associated with root-filled teeth using pyrosequencing. PLoS ONE 2013, 8, e84960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogden, K.A.; Guthmiller, J.M.; Taylor, C.E. Human polymicrobial infections. Lancet 2005, 365, 253–255. [Google Scholar] [CrossRef]
- Vonaesch, P.; Anderson, M.; Sansonetti, P.J. Pathogens, microbiome and the host: Emergence of the ecological Koch’s postulates. FEMS Microbiol. Rev. 2018, 42, 273–292. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, K.; Bertaux, J.; Krome, K.; Hartmann, A.; Scheu, S.; Bonkowski, M. Soil amoebae rapidly change bacterial community composition in the rhizosphere of Arabidopsis thaliana. ISME J. 2009, 3, 675–684. [Google Scholar] [CrossRef]
- Pernthaler, J. Predation on prokaryotes in the water column and its ecological implications. Nat. Rev. Microbiol. 2005, 3, 537–546. [Google Scholar] [CrossRef]
- Jurgens, K.; Matz, C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek 2002, 81, 413–434. [Google Scholar] [CrossRef]
- Bonifait, L.; Charette, S.J.; Filion, G.; Gottschalk, M.; Grenier, D. Amoeba host model for evaluation of Streptococcus suis virulence. Appl. Envrion. Microbiol. 2011, 77, 6271–6273. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.N. The selection of bacterial food by soil amoebae and the toxic effects of bacterial pigments and other products on soil protozoa. Br. J. Exp. Pathol. 1945, 316–325. [Google Scholar]
- Singh, B.N. A method of estimating the numbers of soil protozoa, especially amoebae, based on their differential feeding on bacteria. Ann. Appl. Biol. 1946, 33, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Evstigneeva, A.; Raoult, D.; Karpachevskiy, L.; La Scola, B. Amoeba co-culture of soil specimens recovered 33 different bacteria, including four new species and Streptococcus pneumoniae. Microbiology 2009, 155, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquet, V.E.; Charette, S.J. Amoeba-resisting bacteria found in multilamellar bodies secreted by Dictyostelium discoideum: Social amoebae can also package bacteria. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, M.T.; Rosenthal, B.M.; Haxo, A.F.; Berk, S.G. Enhanced survival of Salmonella enterica in vesicles released by a soilborne Tetrahymena species. Appl Environ. Microbiol. 2005, 71, 1562–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, M.; Amard, V.; Bar-Pinatel, C.; Charpentier, F.; Chatard, J.M.; Desmuyck, Y.; Ihler, S.; Rochet, J.P.; de La Tribouille, V.R.; Saladin, L.; et al. Detection of the amoeba Entamoeba gingivalis in periodontal pockets. Parasite 2014, 21, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, M.; Fresno, M.; Girones, N.; Guillen, N.; Santi-Rocca, J. Reassessing the role of Entamoeba gingivalis in Periodontitis. Front. Cell. Infect. Microbiol. 2018, 8, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, K.S.; Solga, M.D.; Mackey-Lawrence, N.M.; Somlata; Bhattacharya, A.; Petri, W.A., Jr. Trogocytosis by Entamoeba histolytica contributes to cell killing and tissue invasion. Nature 2014, 508, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Munson, M.A.; Pitt-Ford, T.; Chong, B.; Weightman, A.; Wade, W.G. Molecular and cultural analysis of the microflora associated with endodontic infections. J. Dent. Res. 2002, 81, 761–766. [Google Scholar] [CrossRef]
- Zhang, C.; Hou, B.X.; Zhao, H.Y.; Sun, Z. Microbial diversity in failed endodontic root-filled teeth. Chin. Med. J. 2012, 125, 1163–1168. [Google Scholar]
- Mortensen, H.; Winther, J.E.; Birn, H. Periapical granulomas and cysts. An investigation of 1,600 cases. Scand. J. Dent. Res. 1970, 78, 241–250. [Google Scholar] [PubMed]
- Sunde, P.T.; Olsen, I.; Gobel, U.B.; Theegarten, D.; Winter, S.; Debelian, G.J.; Tronstad, L.; Moter, A. Fluorescence in situ shybridisation (FISH) for direct svisualisation of bacteria in periapical lesions of asymptomatic root-filled teeth. Microbiology 2003, 149, 1095–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehnder, M.; Rechenberg, D.K.; Thurnheer, T.; Luthi-Schaller, H.; Belibasakis, G.N. FISHing for gutta-percha-adhered biofilms in purulent post-treatment apical periodontitis. Mol. Oral. Microbiol. 2017, 32, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Debelian, G.J.; Olsen, I.; Tronstad, L. Systemic diseases caused by oral microorganisms. Endod. Dent. Traumatol. 1994, 10, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.B.; Dayer, M.; Prendergast, B.D.; Sandoe, J.; Westaby, S.; Thornhill, M.; British Heart Valve Society. Beyond the antibiotic prophylaxis of infective endocarditis: The problem of dental surveillance. Heart 2013, 99, 363–364. [Google Scholar] [CrossRef]
- Greub, G.; Raoult, D. Microorganisms resistant to free-living amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef] [Green Version]
- Al-Quadan, T.; Price, C.T.; Abu Kwaik, Y. Exploitation of evolutionarily conserved amoeba and mammalian processes by Legionella. Trends Microbiol. 2012, 20, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Coticchia, J.M.; Getnick, G.S.; Yun, R.D.; Arnold, J.E. Age-, Site-, and Time-Specific Differences in Pediatric Deep Neck Abscesses. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Brook, I. Microbiology of abscesses of the head and neck in children. Ann. Otol. Rhinol. Laryngol. 1987, 96, 429–433. [Google Scholar] [CrossRef]
- Stein, D.S.; Nelson, K.E. Endocarditis due to nutritionally deficient streptococci: Therapeutic dilemma. Rev. Infect. Dis. 1987, 9, 908–916. [Google Scholar] [CrossRef]
- Rocas, I.N.; Siqueira, J.F., Jr. Culture-independent detection of Eikenella corrodens and Veillonella parvula in primary endodontic infections. J. Endod. 2006, 32, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, N.; Noiri, Y.; Narimatsu, M.; Ebisu, S. Identification and slocalisation of extraradicular biofilm-forming bacteria associated with refractory endodontic pathogens. Appl. Environ. Microbiol. 2005, 71, 8738–8743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, E.L.; Martinho, F.C.; Leite, F.R.; Nascimento, G.G.; Gomes, B.P. Macrophage cell activation with acute apical abscess contents determined by interleukin-1 Beta and tumor necrosis factor alpha production. J. Endod. 2014, 40, 1752–1757. [Google Scholar] [CrossRef] [PubMed]
- Bor, B.; Bedree, J.K.; Shi, W.; McLean, J.S.; He, X. Saccharibacteria (TM7) in the Human Oral Microbiome. J. Dent. Res. 2019, 98, 500–509. [Google Scholar] [CrossRef]
- Tan, P.T.; Chang, L.Y.; Huang, Y.C.; Chiu, C.H.; Wang, C.R.; Lin, T.Y. Deep neck infections in children. J. Microbiol. Immunol. Infect. 2001, 34, 287–292. [Google Scholar]
- Molmeret, M.; Horn, M.; Wagner, M.; Santic, M.; Abu Kwaik, Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol. 2005, 71, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Adiba, S.; Nizak, C.; van Baalen, M.; Denamur, E.; Depaulis, F. From grazing resistance to pathogenesis: The coincidental evolution of virulence factors. PLoS ONE 2010, 5, e11882. [Google Scholar] [CrossRef]
- Bhaijee, F.; Bell, D. Entamoeba gingivalis in Acute Osteomyelitis of the Mandible. Case. Rep. Med. 2011, 2011, 357301. [Google Scholar] [CrossRef] [Green Version]
- Trim, R.D.; Skinner, M.A.; Farone, M.B.; Dubois, J.D.; Newsome, A.L. Use of PCR to detect Entamoeba gingivalis in diseased gingival pockets and demonstrate its absence in healthy gingival sites. Parasitol. Res. 2011, 109, 857–864. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection type | Primary/Secondary % | 76:2 |
| Gender | Gender Ratio M:F % | 52:48 |
| Age | Age (Years) | 42.1 (SD 14.2) |
| Symptoms | Pain (Y/N) % | 83:17 |
| Signs | Tenderness to percussion (Y/N) % | 56:44 |
| Apical radiolucency extent | Largest extent of radiolucency (mm) | average 6.7 mm (0–25.4 mm) |
| Distribution of large radiolucencies | Largest extent of radiolucency diameter > 5 mm (Y/N) % | 52:48 |
| Fistula | Sinus tract present (Y/N) % | 20:80 |
| Genera (In Order of Abundance) | Species (In Order of Abundance) |
|---|---|
| Streptococcus, Prevotella, Rothia, Veillonella, Neisseria, Marinilactobacillus, Lactobacillus, Staphylococcus, Moraxella, Solobacterium (formerly Bulleidia), Pseudoramibacter/Eubacterium, Peptostreptococcus, Fusobacterium, Leptotrichia, Parvimonas, Dialister, Haemophilus, Paludibacter, Atopobium, Pseudomonas, Actinomyces, Porphyromonas, Enterococcus, Corynebacterium, TG5 and Granulicatella | Rothia mucilaginosa, Prevotella melaninogenica, Veillonella dispar, Neisseria subflava, Marinilactobacillus psychrotolerans, Staphylococcus aureus, Haemophilus parainfluenzae, Prevotella nigrescens, Prevotella tannerae, Prevotella pallens, Propionibacterium acnes, Lactobacillus zeae, Rothia dentocariosa, Rothia aeria, Lactobacillus reuteri, Streptococcus anginosus, Porphyromonas endodontalis, Aggregatibacter segnis, Corynebacterium durum, Prevotella nanceiensis, Treponema socranskii, Haemophilus parahaemolyticus, Alkaliphilus transvaalensis, Veillonella parvula, Haemophilus influenzae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koller, G.; Foschi, F.; Mitchell, P.; Witherden, E.; Bruce, K.; Mannocci, F. Amoebae in Chronic, Polymicrobial Endodontic Infections Are Associated with Altered Microbial Communities of Increased Virulence. J. Clin. Med. 2020, 9, 3700. https://doi.org/10.3390/jcm9113700
Koller G, Foschi F, Mitchell P, Witherden E, Bruce K, Mannocci F. Amoebae in Chronic, Polymicrobial Endodontic Infections Are Associated with Altered Microbial Communities of Increased Virulence. Journal of Clinical Medicine. 2020; 9(11):3700. https://doi.org/10.3390/jcm9113700
Chicago/Turabian StyleKoller, Garrit, Federico Foschi, Philip Mitchell, Elizabeth Witherden, Kenneth Bruce, and Francesco Mannocci. 2020. "Amoebae in Chronic, Polymicrobial Endodontic Infections Are Associated with Altered Microbial Communities of Increased Virulence" Journal of Clinical Medicine 9, no. 11: 3700. https://doi.org/10.3390/jcm9113700