Mitochondrial Function in Peripheral Blood Mononuclear Cells (PBMC) Is Enhanced, Together with Increased Reactive Oxygen Species, in Severe Asthmatic Patients in Exacerbation

 , ,
, ,
Abstract
:1. Introduction
2. Population and Methods
2.1. Patients and Study Design
2.2. Peripheral Blood Mononuclear Cells (PBMC) Isolation
2.3. Mitochondrial Respiration
2.4. Reactive Oxygen Species (ROS) Production
2.4.1. Measurement of Superoxide Anions in Blood
2.4.2. Mitochondrial ROS measurement
2.5. Mitochondrial Dioxygen Consumption of Healthy PBMC in Contact with Heterologous Plasma
2.6. Statistical Analysis
3. Results
3.1. Clinical Characteristics of the Subjects
3.2. Mitochondrial Respiratory Chain Complexes’ Activities Are Enhanced in Asthmatic Patients
3.2.1. Basal Consumption of Dioxygen (V0)
3.2.2. Mitochondrial Complexes I + III + IV + Vactivities
3.2.3. Mitochondrial Complexes I + II + III + IV + Vactivities
3.2.4. Inhibition of the Mitochondrial Complex I by Rotenone
3.2.5. Complex IV Activity
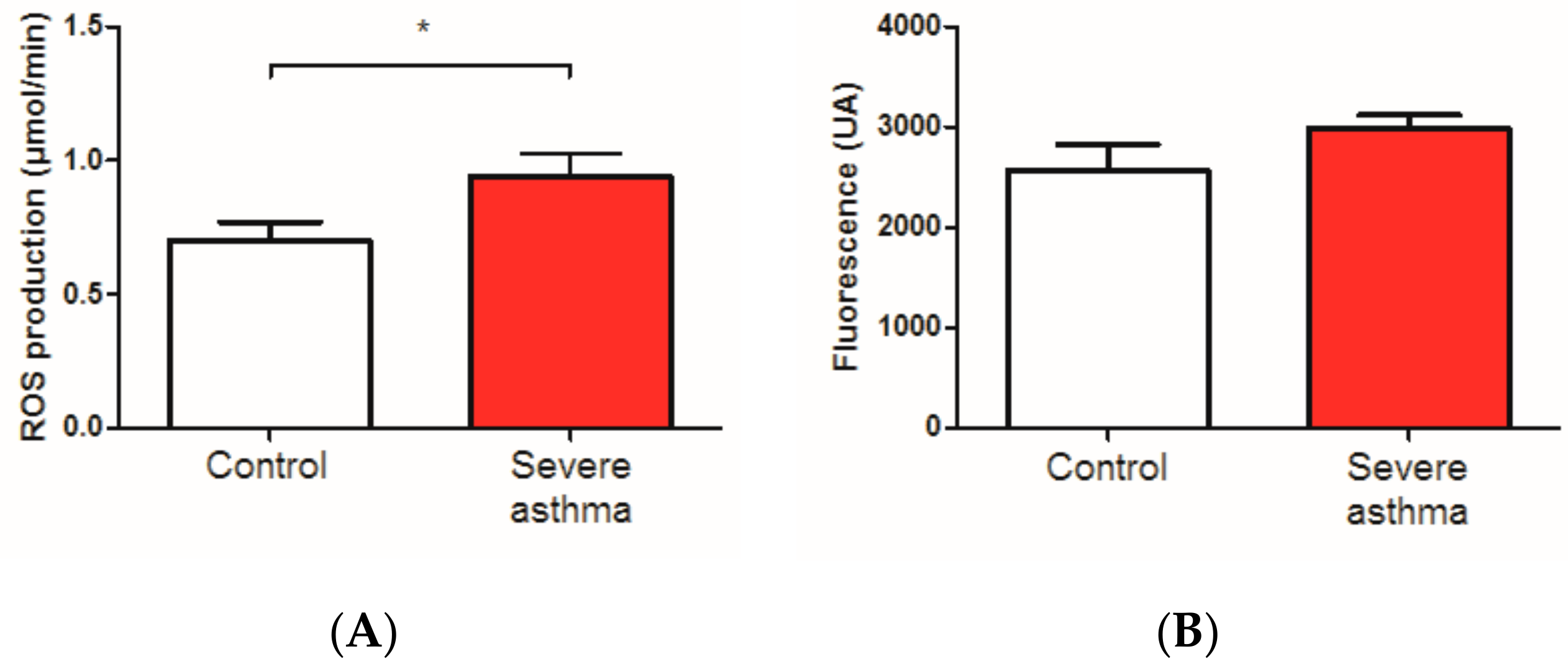
3.3. Reactive Oxygen Species Are Increased in the Blood of Asthmatic Patients
3.4. Effect of Heterologous Plasma of Healthy or Asthmatic Subjects on Mitochondrial Dioxygen Consumption and of Healthy PBMC
4. Discussion
Limitations of the Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADP | adenosine Di-phosphate |
| ATP | adenosine Tri-phosphate |
| AU | Arbitrary unit |
| FEV1 | Forced Expiratory Volume |
| GINA | global initiative for asthma |
| NADPH | Nicotinamide adénine dinucléotide phosphate |
| TMPD | ’N, N, N’N’-tétraméthyl-1,4-phénylènediamine |
References
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- GINA 2019. Available online: www.ginasthma.org (accessed on 17 June 2019).
- Reddy, P.H. Mitochondrial Dysfunction and Oxidative Stress in Asthma: Implications for Mitochondria-Targeted Antioxidant Therapeutics. Pharmaceuticals 2011, 4, 429–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera-Aguirre, L.; Bacsi, A.; Saavedra-Molina, A.; Kurosky, A.; Sur, S.; Boldogh, I. Mitochondrial dysfunction increases allergic airways inflammation. J. Immunol. 2009, 183, 5379–5387. [Google Scholar] [CrossRef] [PubMed]
- Mabalirajan, U.; Dinda, A.K.; Kumar, S.; Roshan, R.; Gupta, P.; Sharma, S.K.; Ghosh, B. Mitochondrial structural changes and dysfunction are associated with experimental allergic asthma. J. Immunol. 2008, 181, 3540–3548. [Google Scholar] [CrossRef]
- Boldogh, I.; Bacsi, A.; Choudhury, B.K.; Dharajiya, N.; Alam, R.; Hazra, T.K.; Mitra, S.; Goldblum, R.M.; Sur, S. ROS generated by pollen NADPH oxidase provide a signal that augments antigen-induced allergic airway inflammation. J. Clin. Investig. 2005, 115, 2169–2179. [Google Scholar] [CrossRef] [Green Version]
- Dharajiya, N.; Choudhury, B.K.; Bacsi, A.; Boldogh, I.; Alam, R.; Sur, S. Inhibiting pollen reduced nicotinamide adenine dinucleotide phosphate oxidase-induced signal by intrapulmonary administration of antioxidants blocks allergic airway inflammation. J. Allergy Clin. Immunol. 2007, 119, 646–653. [Google Scholar] [CrossRef]
- Bacsi, A.; Choudhury, B.K.; Dharajiya, N.; Sur, S.; Boldogh, I. Subpollen particles-carriers of allergenic proteins and oxidases. J. Allergy Clin. Immunol. 2006, 118, 844–850. [Google Scholar] [CrossRef]
- Chan, T.K.; Loh, X.Y.; Peh, H.Y.; Tan, W.N.F.; Tan, W.S.D.; Li, N.; Tay, I.J.J.; Wong, W.S.F.; Engelward, B.P. House dust mite-induced asthma causes oxidative damage and DNA double-strand breaks in the lungs. J. Allergy Clin. Immunol. 2016, 138, 84–96. [Google Scholar] [CrossRef]
- Antus, B. Oxidative Stress Markers in Sputum. Oxid. Med. Cell Longev. 2016. [Google Scholar] [CrossRef]
- Sahiner, U.M.; Birben, E.; Erzurum, S.; Sackesen, C.; Kalayci, O. Oxidative stress in asthma. World Allergy Organ. J. 2011, 4, 151–158. [Google Scholar] [CrossRef]
- Aldakheel, F.M.; Thomas, P.S.; Bourke, J.E.; Matheson, M.C.; Dharmage, S.C.; Lowe, A.J. Relationships between adult asthma and oxidative stress markers and pH in exhaled breath condensate: A systematic review. Allergy 2016, 71, 741–757. [Google Scholar] [CrossRef]
- Louhelainen, N.; Myllärniemi, M.; Rahman, I.; Kinnula, V.L. Airway biomarkers of the oxidant burden in asthma and chronic obstructive pulmonary disease- current and future perspectives. Int. J. Chron. Obstruct. Pulmon. Dis. 2008, 3, 585–603. [Google Scholar]
- Comhair, S.A.; Ricci, K.S.; Arroliga, M.; Lara, A.R.; Dweik, R.A.; Song, W.; Hazen, S.L.; Bleecker, E.R.; Busse, W.W.; Chung, K.F.; et al. Correlation of systemic superoxide dismutase deficiency to airflow obstruction in asthma. Am. J. Respir. Crit. Care Med. 2005, 172, 306–313. [Google Scholar] [CrossRef]
- Comhair, S.A.; Erzurum, S.C. Antioxidant responses to oxidant mediated lung diseases. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, L246–L255. [Google Scholar] [CrossRef]
- Iyer, D.; Mishra, N.; Agrawal, A. Mitochondrial Function in Allergic Disease. Curr. Allergy Asthma Rep. 2017, 17, 29. [Google Scholar] [CrossRef]
- Ijsselmuiden, A.J.; Musters, R.J.; de Ruiter, G.; van Heerebeek, L.; Alderse-Baas, F.; van Schilfgaarde, M.; Leyte, A.; Tangelder, G.J.; Laarman, G.J.; Paulus, W.J. Circulating white blood cells and platelets amplify oxidative stress in heart failure. Nat. Clin. Pract. Cardiovasc. Med. 2008, 5, 811–820. [Google Scholar] [CrossRef]
- Li, P.; Wang, B.; Sun, F.; Li, Y.; Li, Q.; Lang, H.; Zhao, Z.; Gao, P.; Zhao, Y.; Shang, Q.; et al. Mitochondrial respiratory dysfunctions of blood mononuclear cells link with cardiac disturbance in patients with early-stage heart failure. Sci. Rep. 2015. [Google Scholar] [CrossRef]
- Coluccia, R.; Raffa, S.; Ranieri, D.; Micaloni, A.; Valente, S.; Salerno, G.; Scrofani, C.; Testa, M.; Gallo, G.; Pagannone, E.; et al. Chronic heart failure is characterized by altered mitochondrial function and structure in circulating leucocytes. Oncotarget 2018, 9, 35028–35040. [Google Scholar] [CrossRef]
- Clere-Jehl, R.; Helms, J.; Kassem, M.; Le Borgne, P.; Delabranche, X.; Charles, A.L.; Geny, B.; Meziani, F.; Bilbault, P. Septic Shock Alters Mitochondrial Respiration of Lymphoid Cell-Lines and Human Peripheral Blood Mononuclear Cells: The Role of Plasma. Shock 2019, 51, 97–104. [Google Scholar] [CrossRef]
- Kaminski, M.M.; Sauer, S.W.; Klemke, C.D.; Suss, D.; Okun, J.G.; Krammer, P.H.; Gulow, K. Mitochondrial reactive oxygen species control T cell activation by regulating IL-2 and IL-4 expression: Mechanism of ciprofloxacin-mediated immunosuppression. J. Immunol. 2010, 184, 4827–4841. [Google Scholar] [CrossRef]
- Pattnaik, B.; Bodas, M.; Bhatraju, N.K.; Ahmad, T.; Pant, R.; Guleria, R.; Ghosh, B.; Agrawal, A. IL-4 promotes asymmetric dimethylarginine accumulation, oxo-nitrative stress, and hypoxic response-induced mitochondrial loss in airway epithelial cells. J. Allergy. Clin. Immunol. 2016, 138, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Barnig, C.; Charles, A.L.; Poirot, A.; Meyer, A.; Clere-Jehl, R.; de Blay, F.; Geny, B. Effect of nasal allergen challenge in allergic rhinitis on mitochondrial function of peripheral blood mononuclear cells. Ann. Allergy Asthma. Immunol. 2017, 118, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Marchandot, B.; Kibler, M.; Charles, A.L.; Trinh, A.; Petit Eisenmann, H.; Zeyons, F.; Von Hunolstein, J.J.; Reyde, A.; Matsushita, K.; Kindo, M.; et al. Does Transcatheter Aortic Valve Replacement Modulate the Kinetic of Superoxide Anion Generation? Antioxid. Redox. Signal 2019, 31, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Rajesh, M.; Yoshihiro, K.; Haskó, G.; Pacher, P. Simple quantitative detection of mitochondrial superoxide production in live cells. Biochem. Biophys. Res. Commun. 2007, 358, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.J.; Neufer, P.D. Type II skeletal myofibers possess unique properties that potentiate mitochondrial H(2)O(2) generation. Am. J. Physiol. Cell Physiol. 2006, 290, C844–C851. [Google Scholar] [CrossRef] [PubMed]
- Wolff, V.; Schlagowski, A.I.; Rouyer, O.; Charles, A.L.; Singh, F.; Auger, C.; Schini-Kerth, V.; Marescaux, C.; Raul, J.S.; Zoll, J.; et al. Tetrahydrocannabinol induces brain mitochondrial respiratory chain dysfunction and increases oxidative stress: A potential mechanism involved in cannabis-related stroke. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef]
- Sumbalova, Z.; Droescher, S.; Hiller, E.; Chang, S.C.; Garcia-Souza, L.F.; Calabria, E.; Volani, C.; Krumschnabel, G.; Gnaiger, E. O2k-Protocols: Isolation of peripheral blood mononuclear cells and platelets from human blood for HRR. Mitochondr. Physiol. Network. 2018, 21, 1–16. [Google Scholar]
- Bonato, M.; Tiné, M.; Bazzan, E.; Biondini, D.; Saetta, M.; Baraldo, S. Early Airway Pathological Changes in Children: New Insights into the Natural History of Wheezing. J. Clin. Med. 2019, 8, 1180. [Google Scholar] [CrossRef]
- Maestraggi, Q.; Lebas, B.; Clere-Jehl, R.; Ludes, P.O.; Chamaraux-Tran, T.N.; Schneider, F.; Diemunsch, P.; Geny, B.; Pottecher, J. Skeletal Muscle and Lymphocyte Mitochondrial Dysfunctions in Septic Shock Trigger ICU-Acquired Weakness and Sepsis-Induced Immunoparalysis. Biomed. Res. Int. 2017. [Google Scholar] [CrossRef]
- Rahman, I.; MacNee, W. Oxidative stress and regulation of glutathione in lung inflammation. Eur. Respir. J. 2000, 16, 534–554. [Google Scholar] [CrossRef]
- Dikalov, S. Cross talk between mitochondria and NAPH oxidases. Free Radic. Biol. Med. 2011, 51, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Bhatraju, N.K.; Agrawal, A. Mitochondrial Dysfunction Linking Obesity and Asthma. Ann. Am. Thorac. Soc. 2017, 14, S368–S373. [Google Scholar] [CrossRef] [PubMed]
- Létuvé, S.; Druilhe, A.; Grandsaigne, M.; Aubier, M.; Pretolani, M. Critical role of mitochondria, but not caspases, during glucocorticosteroid-induced human eosinophil apoptosis. Am. J. Respir. Cell Mol. Biol. 2002, 5, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Mabalirajan, U.; Ahmad, T.; Ghosh, B. Emerging interface between metabolic syndrome and asthma. Am. J. Respir. Cell Mol. Biol. 2011, 44, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Laverny, G.; Bernardi, L.; Charles, A.L.; Alsaleh, G.; Pottecher, J.; Sibilia, J.; Geny, B. Mitochondria: An Organelle of Bacterial Origin Controlling Inflammation. Front. Immunol. 2018, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Mildner, A.; Jung, S. Development and function of dendritic cell subsets. Immunity 2014, 40, 642–656. [Google Scholar] [CrossRef]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef]
- Lejay, A.; Meyer, A.; Schlagowski, A.I.; Charles, A.L.; Singh, F.; Bouitbir, J.; Pottecher, J.; Chakfé, N.; Zoll, J.; Geny, B. Mitochondria: Mitochondrial participation in ischemia-reperfusion injury in skeletal muscle. Int. J. Biochem. Cell Biol. 2014, 50, 101–105. [Google Scholar] [CrossRef]
- Leffler, J.; Read, J.F.; Jones, A.C.; Mok, D.; Hollams, E.M.; Laing, I.A.; Le Souef, P.N.; Sly, P.D.; Kusel, M.M.H.; de Klerk, N.H.; et al. Progressive increase of FcεRI expression across several PBMC subsets is associated with atopy and atopic asthma within school-aged children. Pediatr. Allergy Immunol. 2019, 30, 646–653. [Google Scholar] [CrossRef]
- Leffler, J.; Jones, A.C.; Hollams, E.M.; Prastanti, F.; Le Souëf, P.N.; Holt, P.G.; Bosco, A.; Laing, I.A.; Strickland, D.H. Basophil counts in PBMC populations during childhood acute wheeze/asthma are associated with future exacerbations. J. Allergy Clin. Immunol. 2018, 142, 1639–1641. [Google Scholar] [CrossRef]
- Price, D.B.; Rigazio, A.; Campbell, J.D.; Bleecker, E.R.; Corrigan, C.J.; Thomas, M.; Wenzel, S.E.; Wilson, A.M.; Small, M.B.; Gopalan, G.; et al. Blood eosinophil count and prospective annual asthma disease burden: A UK cohort study. Lancet Respir. Med. 2015, 3, 849–858. [Google Scholar] [CrossRef]
- Sidhu, S.S.; Yuan, S.; Innes, A.L.; Kerr, S.; Woodruff, P.G.; Hou, L.; Muller, S.J.; Fahy, J.V. Roles of epithelial cell-derived periostin in TGF-beta activation, collagen production, and collagen gel elasticity in asthma. Proc. Natl. Acad. Sci. USA 2010, 107, 14170–14175. [Google Scholar] [CrossRef] [PubMed]
- Takayama, G.; Arima, K.; Kanaji, T.; Toda, S.; Tanaka, H.; Shoji, S.; McKenzie, A.N.J.; Nagai, H.; Hotokebuchi, T.; Izuhara, K. Periostin: A novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and IL-13 signals. J. Allergy Clin. Immunol. 2006, 118, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Das, S.R.; Emin, M.T.; Wei, M.; Sun, L.; Westphalen, K.; Rowlands, D.J.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Mitochondrial transfer from bone-marrow-derived stromal cells to pulmonary alveoli protects against acute lung injury. Nat. Med. 2012, 18, 759–765. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group (n = 20) | Severe Asthmatic Patients (n = 20) | |
|---|---|---|
| Mean age (years) | 50.10 ± 3.77 | 50.95 ± 4.06 |
| Women | 14 | 15 |
| Male | 6 | 5 |
| Mean body mass index (BMI) (kg/m2) | 23.0 ± 0.72 | 28.01 ± 1.47 |
| Mean smoking habits (PA) | 1.70 ± 0.80 | 1.90 ± 0.69 |
| Atopy | 0 | 16 |
| Medical history: | ||
| Diabetes | 1 | 4 |
| Arterial hypertension | 3 | 8 |
| Venous thrombosis/pulmonary embolism | 0 | 1 |
| Other lung diseases | 0 | 2 |
| Neoplasia | 0 | 1 |
| Acute coronary syndrom with preserved LVEF | 2 | 1 |
| Blood eosinophil (Normal values: <600/mm3) | ||
| <300/mm3 | 9 | |
| 300–500/mm3 | 2 | |
| 500–1000/mm3 | 9 | |
| Blood neutrophils (Normal values: 1500–7500/mm3) | ||
| <7500/mm3 | 16 | |
| >7500/mm3 | 4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ederlé, C.; Charles, A.-L.; Khayath, N.; Poirot, A.; Meyer, A.; Clere-Jehl, R.; Andres, E.; De Blay, F.; Geny, B. Mitochondrial Function in Peripheral Blood Mononuclear Cells (PBMC) Is Enhanced, Together with Increased Reactive Oxygen Species, in Severe Asthmatic Patients in Exacerbation. J. Clin. Med. 2019, 8, 1613. https://doi.org/10.3390/jcm8101613
Ederlé C, Charles A-L, Khayath N, Poirot A, Meyer A, Clere-Jehl R, Andres E, De Blay F, Geny B. Mitochondrial Function in Peripheral Blood Mononuclear Cells (PBMC) Is Enhanced, Together with Increased Reactive Oxygen Species, in Severe Asthmatic Patients in Exacerbation. Journal of Clinical Medicine. 2019; 8(10):1613. https://doi.org/10.3390/jcm8101613
Chicago/Turabian StyleEderlé, Carole, Anne-Laure Charles, Naji Khayath, Anh Poirot, Alain Meyer, Raphaël Clere-Jehl, Emmanuel Andres, Frédéric De Blay, and Bernard Geny. 2019. "Mitochondrial Function in Peripheral Blood Mononuclear Cells (PBMC) Is Enhanced, Together with Increased Reactive Oxygen Species, in Severe Asthmatic Patients in Exacerbation" Journal of Clinical Medicine 8, no. 10: 1613. https://doi.org/10.3390/jcm8101613