Immunomodulatory Properties of Adipose-Derived Stem Cells Treated with 5-Azacytydine and Resveratrol on Peripheral Blood Mononuclear Cells and Macrophages in Metabolic Syndrome Animals

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Qualification of Horses
2.2. ASC Isolation and Culture
2.3. Extraction and Culture of Peripheral Blood Mononuclear Cells (PBMC)
2.4. Flow Cytometry Analysis
2.4.1. ASC-PBMC Co-Culture
2.4.2. ASC-RAW264.7 Co-Culture
2.5. Nitric Oxide (NO) and Superoxide Dismutase (SOD) Levels Analysis
2.6. Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.7. ELISA Tests
2.8. Statistical Analysis
3. Results
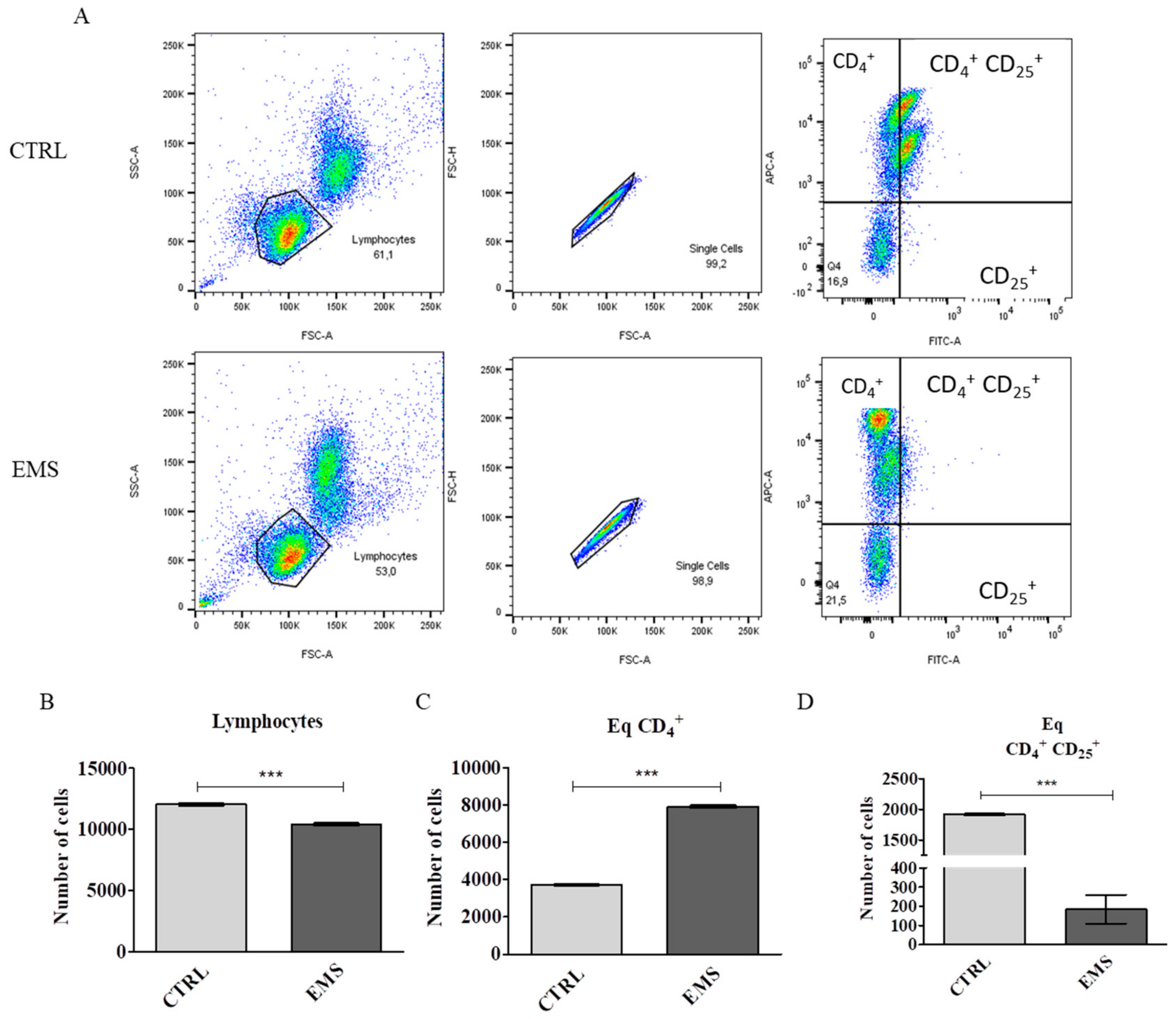
3.1. Flow Cytometry Analysis of PBMCs
3.2. Gene Expression After ASC-PBMC Co-Culture
3.3. Cytokines, NO, and SOD Amount after ASC-PBMC Co-Culture
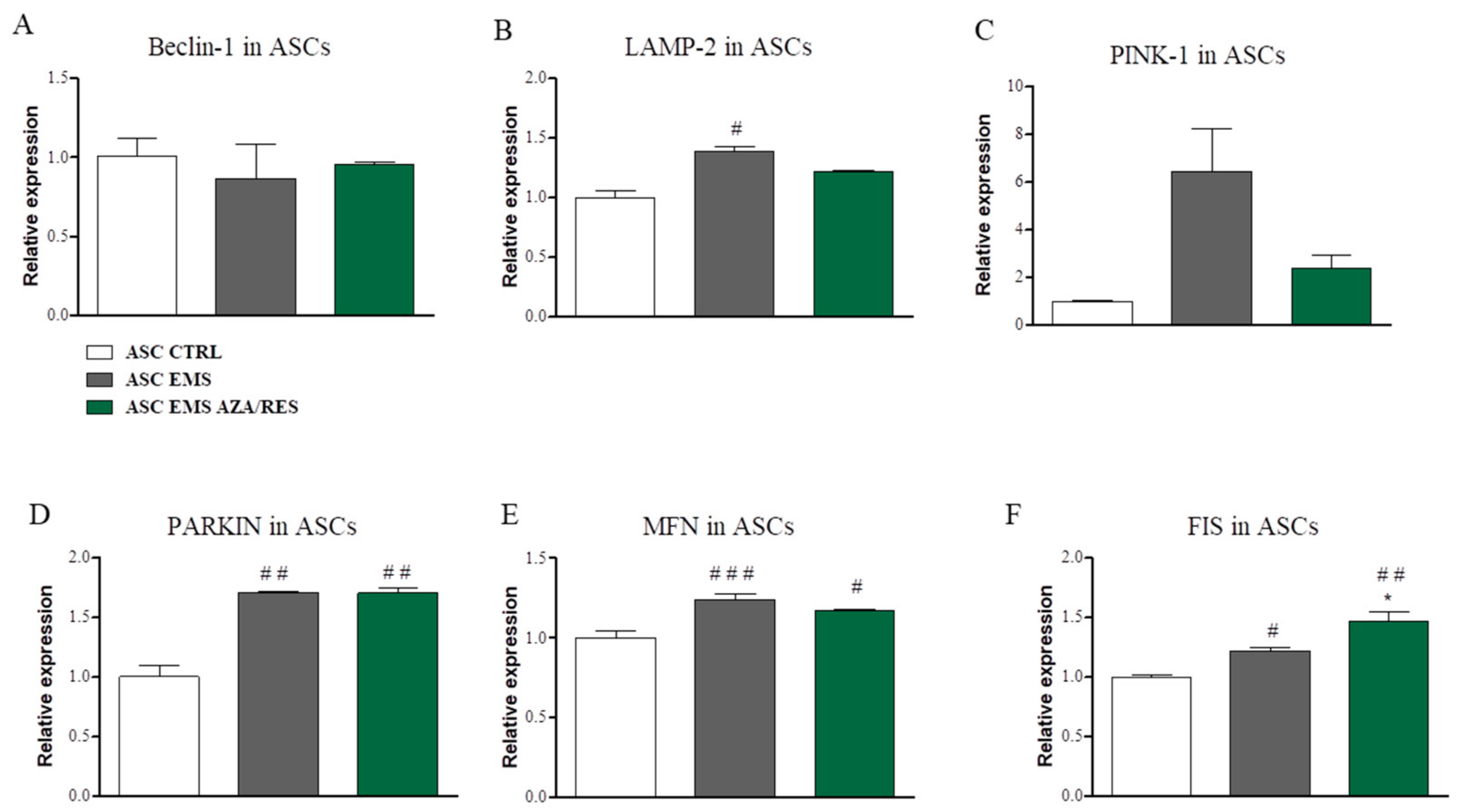
3.4. Evaluation of Autophagy-Related Genes after ASC-PBMC Co-Culture
3.5. Evaluation of Mitophagy-Related Genes after Co ASC-PBMC Co-Culture
3.6. Co-Culture of ASC with RAW264.7 Macrophages
3.7. Auto- and Mitophagy after ASC-RAW264.7 Co-Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yu, H.; Côté, P.; Shearer, H.M.; Wong, J.J.; Sutton, D.A.; Randhawa, K.A.; Varatharajan, S.; Southerst, D.; Mior, S.A.; Ameis, A.; et al. Effectiveness of passive physical modalities for shoulder pain: Systematic review by the Ontario protocol for traffic injury management collaboration. Phys. Ther. 2015, 95, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Castillo, D.T.; Lacefield, K.; C’de Baca, J.; Blankenship, A.; Qualls, C. Effectiveness of group-delivered cognitive therapy and treatment length in women veterans with PTSD. Behav. Sci. 2014, 4, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro-Spinetti, E.; de Mello, W.; Trindade, L.S.; Taub, D.D.; Taichman, R.S.; Balduino, A. Human bone marrow mesenchymal progenitors: Perspectives on an optimized in vitro manipulation. Front. Cell Dev. Biol. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.d.V.; Telles, P.D.d.S.; Nascimento, I.L.O. Immunological characteristics of mesenchymal stem cells. Rev. Bras. Hematol. Hemoter. 2013, 35, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.L.; Tang, K.C.; Patel, A.P.; Bonilla, L.M.; Pierobon, N.; Ponzio, N.M.; Rameshwar, P. Antigen-presenting property of mesenchymal stem cells occurs during a narrow window at low levels of interferon-γ. Blood 2006, 107, 4817–4824. [Google Scholar] [CrossRef] [PubMed]
- Puissant, B.; Barreau, C.; Bourin, P.; Clavel, C.; Corre, J.; Bousquet, C.; Taureau, C.; Cousin, B.; Abbal, M.; Laharrague, P.; et al. Immunomodulatory effect of human adipose tissue-derived adult stem cells: Comparison with bone marrow mesenchymal stem cells. Br. J. Haematol. 2005, 129, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.-F.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar] [CrossRef] [PubMed]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+CD25highFOXP3+ regulatory T cells. Stem Cells 2008, 26, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Kornicka, K.; Szlapka-Kosarzewska, J.; Weiss, C. Excessive endoplasmic reticulum stress correlates with impaired mitochondrial dynamics, mitophagy and apoptosis, in liver and adipose tissue, but not in Muscles in EMS horses. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, T.C.; Tipton, T.; McFarlane, D. Neutrophil and cytokine dysregulation in hyperinsulinemic obese horses. Vet. Immunol. Immunopathol. 2012, 145, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Burns, T.A.; Geor, R.J.; Mudge, M.C.; McCutcheon, L.J.; Hinchcliff, K.W.; Belknap, J.K. Proinflammatory cytokine and chemokine gene expression profiles in subcutaneous and visceral adipose tissue depots of insulin-resistant and insulin-sensitive light breed horses. J. Vet. Intern. Med. 2010, 24, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Vick, M.M.; Murphy, B.A.; Sessions, D.R.; Reedy, S.E.; Kennedy, E.L.; Horohov, D.W.; Cook, R.F.; Fitzgerald, B.P. Effects of systemic inflammation on insulin sensitivity in horses and inflammatory cytokine expression in adipose tissue. Am. J. Vet. Res. 2008, 69, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Kornicka, K.; Basinska, K.; Czyrek, A. Equine metabolic syndrome affects viability, senescence, and stress factors of equine adipose-derived mesenchymal stromal stem cells: New insight into EqASCs isolated from EMS horses in the context of their aging. Oxid. Med. Cell. Longev. 2016, 2016, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Kornicka, K.; Grzesiak, J.; Śmieszek, A.; Szłapka, J. Macroautophagy and Selective Mitophagy Ameliorate Chondrogenic Differentiation Potential in Adipose Stem Cells of Equine Metabolic Syndrome: New Findings in the Field of Progenitor Cells Differentiation. Available online: https://www.hindawi.com/journals/omcl/2016/3718468/ (accessed on 27 July 2017).
- Marycz, K.; Kornicka, K.; Marędziak, M.; Golonka, P.; Nicpoń, J. Equine metabolic syndrome impairs adipose stem cells osteogenic differentiation by predominance of autophagy over selective mitophagy. J. Cell. Mol. Med. 2016, 20, 2384–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornicka, K.; Marycz, K.; Marędziak, M.; Tomaszewski, K.A.; Nicpoń, J. The effects of the DNA methyltranfserases inhibitor 5-Azacitidine on ageing, oxidative stress and DNA methylation of adipose derived stem cells. J. Cell. Mol. Med. 2017, 21, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Nawrocka, D.; Lis-Bartos, A.; Marędziak, M.; Marycz, K. Polyurethane–polylactide-based material doped with resveratrol decreases senescence and oxidative stress of adipose-derived mesenchymal stromal stem cell (ASCs). RSC Adv. 2017, 7, 24070–24084. [Google Scholar] [CrossRef] [Green Version]
- Kuballa, P.; Nolte, W.M.; Castoreno, A.B.; Xavier, R.J. Autophagy and the immune system. Annu. Rev. Immunol. 2012, 30, 611–646. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Weiss, C.; Śmieszek, A.; Kornicka, K. Evaluation of oxidative stress and mitophagy during adipogenic differentiation of adipose-derived stem cells isolated from equine metabolic syndrome (EMS) horses. Stem Cells Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Van der Weerd, K.; Dik, W.A.; Schrijver, B.; Schweitzer, D.H.; Langerak, A.W.; Drexhage, H.A.; Kiewiet, R.M.; van Aken, M.O.; van Huisstede, A.; van Dongen, J.J.M.; et al. Morbidly obese human subjects have increased peripheral blood CD4+ T cells with skewing toward a Treg- and Th2-dominated phenotype. Diabetes 2012, 61, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, K.; Yan, X.; Shinmura, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yamamoto, T.; Anzai, A.; Isobe, S.; Yoshida, N.; et al. Obesity accelerates T cell senescence in murine visceral adipose tissue. J. Clin. Investig. 2016, 126, 4626–4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basinska, K.; Marycz, K.; Śmieszek, A.; Nicpoń, J. The production and distribution of IL-6 and TNF-α in subcutaneous adipose tissue and their correlation with serum concentrations in Welsh ponies with equine metabolic syndrome. J. Vet. Sci. 2015, 16, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Rudensky, A.Y. Foxp3 in control of the regulatory T cell lineage. Nat. Immunol. 2007, 8, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan-Bogdan, M.; McDonnell, M.E.; Shin, H.; Rehman, Q.; Hasturk, H.; Apovian, C.M.; Nikolajczyk, B.S. Elevated proinflammatory cytokine production by a skewed T cell compartment requires monocytes and promotes inflammation in type 2 diabetes. J. Immunol. 2011, 186, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.-M.; Brandhorst, G.; Czepluch, F.; Lankeit, M.; Eberle, C.; Herzberg, S.; Faustin, V.; Riggert, J.; Oellerich, M.; Hasenfuss, G.; et al. Circulating regulatory T cells are reduced in obesity and may identify subjects at increased metabolic and cardiovascular risk. Obesity 2013, 21, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A.; Lee, J.; Goldfine, A.B.; Benoist, C.; Shoelson, S.; et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundin, M.; D’arcy, P.; Johansson, C.C.; Barrett, A.J.; Lönnies, H.; Sundberg, B.; Nava, S.; Kiessling, R.; Mougiakakos, D.; Le Blanc, K. Multipotent mesenchymal stromal cells express FoxP3: A marker for the immunosuppressive capacity? J. Immunother. 2011, 34, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Leto Barone, A.A.; Khalifian, S.; Lee, W.P.A.; Brandacher, G. Immunomodulatory effects of adipose-derived stem cells: Fact or fiction? Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Abdi, R.; Fiorina, P.; Adra, C.N.; Atkinson, M.; Sayegh, M.H. Immunomodulation by mesenchymal stem cells: A potential therapeutic strategy for type 1 diabetes. Diabetes 2008, 57, 1759–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubayashi, S.; Akasu, F.; Kasuga, Y.; Snow, K.; Keystone, E.; Volpé, R. In vitro production of interferon-gamma by peripheral blood from patients with Graves’ disease, Hashimoto’s thyroiditis and rheumatoid arthritis. Clin. Exp. Immunol. 1990, 82, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xiao, C.; Wang, P.; Xu, W.; Zhang, A.; Li, Q.; Xu, X. The alteration of Th1/Th2/Th17/Treg paradigm in patients with type 2 diabetes mellitus: Relationship with diabetic nephropathy. Hum. Immunol. 2014, 75, 289–296. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Ryan, J.M.; Tobin, L.; Murphy, M.J.; Barry, F.P.; Mahon, B.P. Cell contact, prostaglandin E(2) and transforming growth factor beta 1 play non-redundant roles in human mesenchymal stem cell induction of CD4+CD25(High) forkhead box P3+ regulatory T cells. Clin. Exp. Immunol. 2009, 156, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Bao, P.; Liu, G.; Wei, Y. Association between IL-6 and related risk factors of metabolic syndrome and cardiovascular disease in young rats. Int. J. Clin. Exp. Med. 2015, 8, 13491–13499. [Google Scholar] [PubMed]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh, S.; Mazloom, H.; Sadeghi, A.; Emamgholipour, S.; Golestani, A.; Noorbakhsh, F.; Khoshniatnikoo, M.; Meshkani, R. Evidence for the link between defective autophagy and inflammation in peripheral blood mononuclear cells of type 2 diabetic patients. J. Physiol. Biochem. 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hirota, Y.; Kang, D.; Kanki, T. The physiological role of mitophagy: New insights into phosphorylation events. Int. J. Cell Biol. 2012, 2012, 354914. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-X.; Yin, X.-M. Mitophagy: Mechanisms, pathophysiological roles, and analysis. Biol. Chem. 2012, 393, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.; Deen, N.; Zamani, S.; Hasnat, M.A. Mitophagy and the release of inflammatory cytokines. Mitochondrion 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, S.-P.; Fu, J.-S.; Bai, L.; Guo, L. Resveratrol augments therapeutic efficiency of mouse bone marrow mesenchymal stem cell-based therapy in experimental autoimmune encephalomyelitis. Int. J. Dev. Neurosci. 2016, 49, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-H.; Ma, Q.-Y.; Wang, L.-C.; Sha, H.-C.; Wu, S.-L.; Zhang, M. Effect of resveratrol on peritoneal macrophages in rats with severe acute pancreatitis. Inflamm. Res. 2005, 54, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Elmali, N.; Baysal, O.; Harma, A.; Esenkaya, I.; Mizrak, B. Effects of resveratrol in inflammatory arthritis. Inflammation 2007, 30, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Cerón, J.J.; Tomás-Barberán, F.; Dolara, P.; Espín, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, J.; Li, X.; Zhou, Q.; Bai, J.; Shi, Y.; Le, G. Resveratrol prevents suppression of regulatory T-cell production, oxidative stress, and inflammation of mice prone or resistant to high-fat diet-induced obesity. Nutr. Res. 2013, 33, 971–981. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence 5′-3′ | Amplicon Length (bp) | Accession no. |
|---|---|---|---|---|
| INFNG | F: | CACCAGCAAGCTGGAAGACT | 163 | NM_001081949.1 |
| R: | CCGGCCTCGAAATGGATTCT | |||
| FOXP3 | F: | AGATGCTGGCCGAGGTCAAC | 149 | XM_023633195.1 |
| R: | TGCGGAACTCGAACTCATCC | |||
| IL-10 | F: | ATAAGAGCAAGGCAGTGGAGC | 77 | NM_001082490.1 |
| R: | ACTCATGGCTTTGTAGACACC | |||
| TGF beta | F: | ATTCCTGGCGCTACCTCAGT | 197 | NM_001081849.1 |
| R: | GCTGGAACTGAACCCGTTGAT | |||
| LC3 | F: | TTCTGAGACACAGTCGGAGC | 128 | XM_001493613.6 |
| R: | CTTTGTTCGAAGGTGTGGCG | |||
| Beclin–1 | F: | GATGCGTTATGCCCAGATGC | 233 | XM_014833759.1 |
| R: | AACGGCAGCTCCTCTGAAAT | |||
| LAMP-2 | F: | GCACCCCTGGGAAGTTCTTA | 147 | XM_014831347.1 |
| R: | ATCCAGCGAACACTCTTGGG | |||
| PINK-1 | F: | GCACAATGAGCCAGGAGCTA | 298 | XM_014737247.1 |
| R: | GGGGTATTCACGCGAAGGTA | |||
| PARKIN | F: | TCCCAGTGGAGGTCGATTCT | 218 | XM_014858374.1 |
| R: | CCCTCCAGGTGTGTTCGTTT | |||
| MFN | F: | AAGTGGCATTTTTCGGCAGG | 217 | XM_001495170.5 |
| R: | TCCATATGAAGGGCATGGGC | |||
| FIS | F: | GGTGCGAAGCAAGTACAACG | 118 | XM_001504462.4 |
| R: | GTTGCCCACAGCCAGATAGA | |||
| GAPDH | F: | GATGCCCCAATGTTTGTGA | 250 | XM_014866500.1 |
| R: | AAGCAGGGATGATGTTCTGG | |||
| TNF-α | F: | ACAGAAAGCATGATCCGCGA | 295 | NM_013693.3 |
| R: | CTTGGTGGTTTGCTACGACG | |||
| IL-1β | F: | TGCCACCTTTTGACAGTGATG | 138 | NM_008361.4 |
| R: | TGATGTGCTGCTGCGAGATT | |||
| IL-6 | F: | GAGGATACCACTCCCAACAGACC | 141 | NM_001314054.1 |
| R: | AAGTGCATCATCGTTGTTCATACA | |||
| GAPDH | F: | TGCACCACCAACTGCTTAG | 177 | XM_017321385.1 |
| R: | GGATGCAGGGATGATGTTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kornicka, K.; Śmieszek, A.; Węgrzyn, A.S.; Röcken, M.; Marycz, K. Immunomodulatory Properties of Adipose-Derived Stem Cells Treated with 5-Azacytydine and Resveratrol on Peripheral Blood Mononuclear Cells and Macrophages in Metabolic Syndrome Animals. J. Clin. Med. 2018, 7, 383. https://doi.org/10.3390/jcm7110383
Kornicka K, Śmieszek A, Węgrzyn AS, Röcken M, Marycz K. Immunomodulatory Properties of Adipose-Derived Stem Cells Treated with 5-Azacytydine and Resveratrol on Peripheral Blood Mononuclear Cells and Macrophages in Metabolic Syndrome Animals. Journal of Clinical Medicine. 2018; 7(11):383. https://doi.org/10.3390/jcm7110383
Chicago/Turabian StyleKornicka, Katarzyna, Agnieszka Śmieszek, Agnieszka Sławomira Węgrzyn, Michael Röcken, and Krzysztof Marycz. 2018. "Immunomodulatory Properties of Adipose-Derived Stem Cells Treated with 5-Azacytydine and Resveratrol on Peripheral Blood Mononuclear Cells and Macrophages in Metabolic Syndrome Animals" Journal of Clinical Medicine 7, no. 11: 383. https://doi.org/10.3390/jcm7110383