The Anti-Inflammatory Effects of Fermented Herbal Roots of Asparagus cochinchinensis in an Ovalbumin-Induced Asthma Model

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of BAW
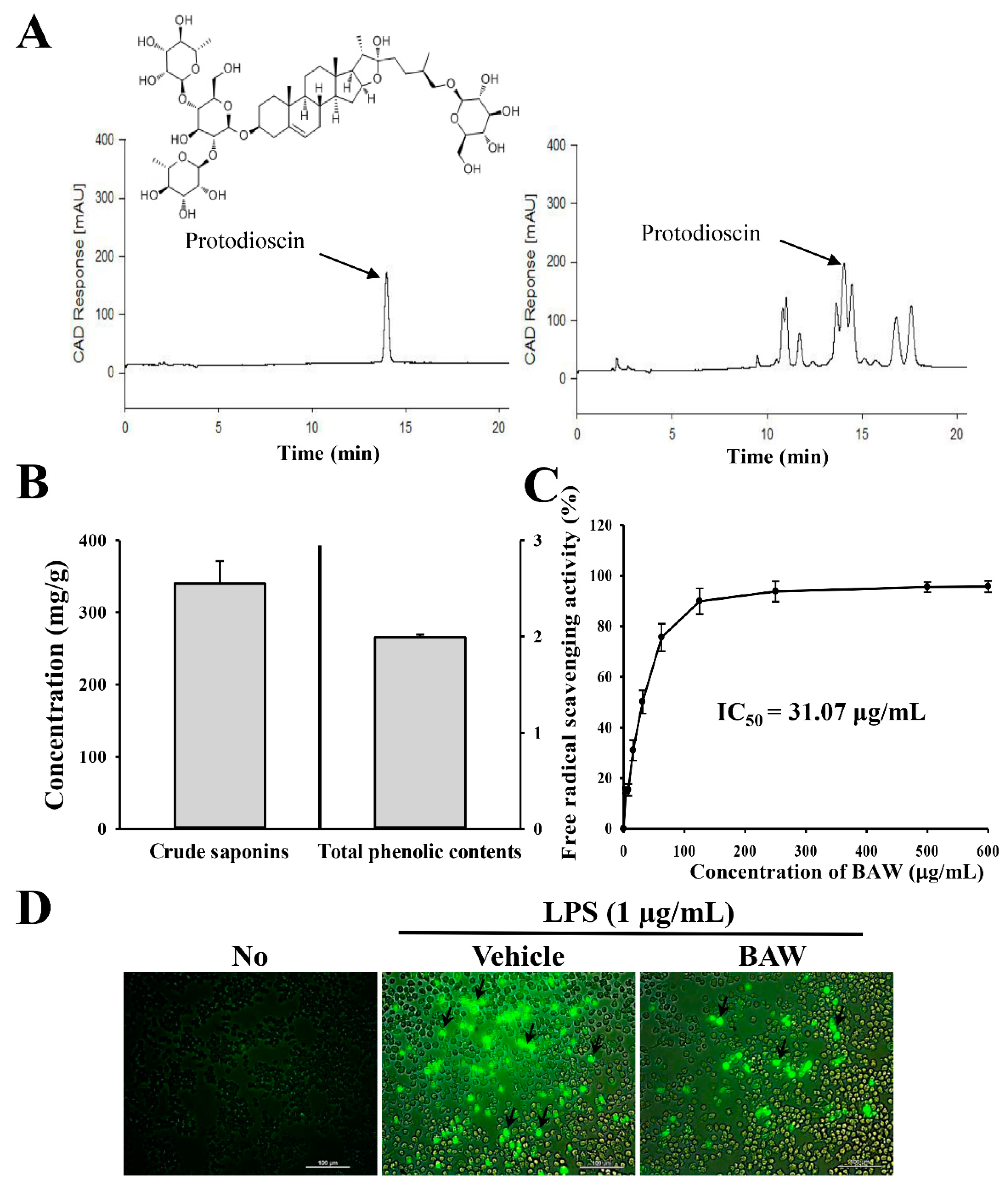
2.2. Measurement of Bioactive Compounds in BAW
2.3. High Performance Liquid Chromatography (HPLC) Analysis
2.4. Scavenging Activity of Free Radical
2.5. Analysis of Intracellular Reactive Oxigen Species (ROS) Level
2.6. Design of Animal Experiment
2.7. Enumeration of Total Cells in Bronchoalveolar Lavage Fluid (BALF)
2.8. Enzyme-Linked Immunosorbent Assay (ELISA) for IL-4 in BALF and Serum
2.9. Detection of OVA-Specific IgE Concentration
2.10. Histopathological Analysis
2.11. Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis for Cytokine Gene Expression
2.12. AChE Activity Analysis
2.13. Western Blot Analysis
2.14. Statistical Analysis
3. Results
3.1. Bioactive Components and Antioxidant Activity of BAW
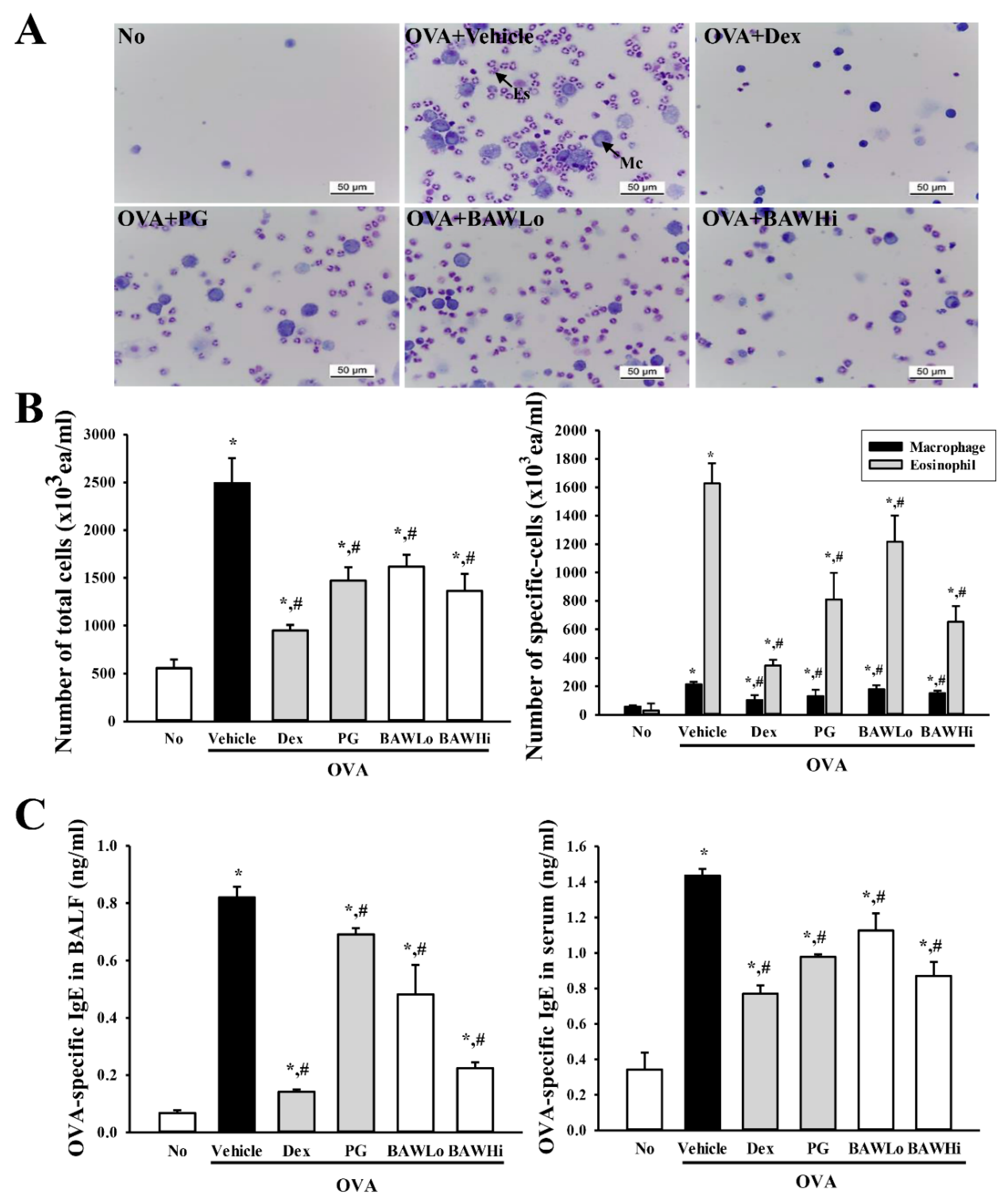
3.2. BAW Suppress the Influx of Leukocytes in BALF of OVA-Induced Asthma Model
3.3. BAW Effect on IgE Concentration in Serum and BALF of OVA-Induced Asthma Model
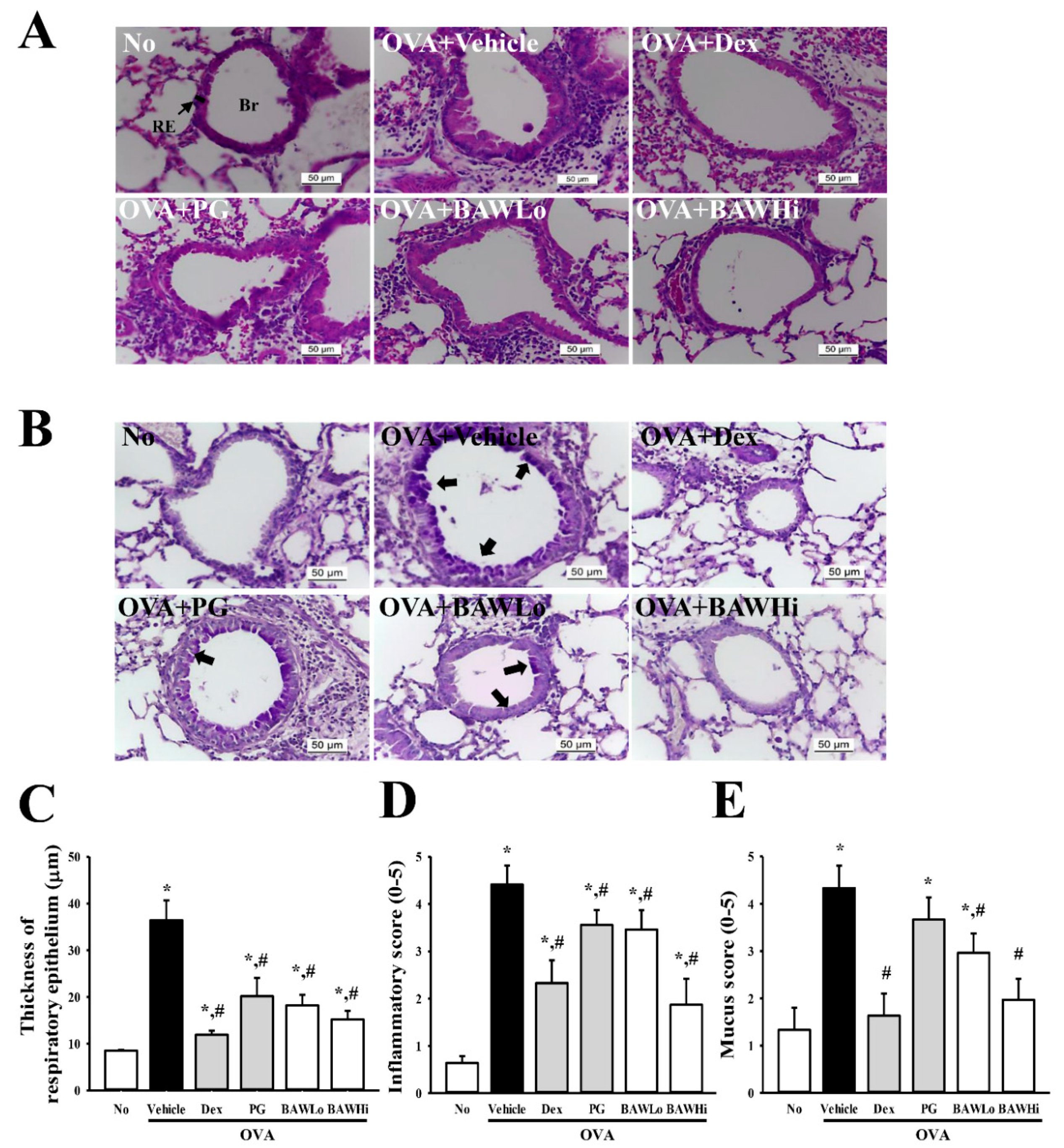
3.4. Alterations in Inflammatory Cell Infiltration and Epithelial Damage of OVA-Induced Asthma Model Treated with BAW
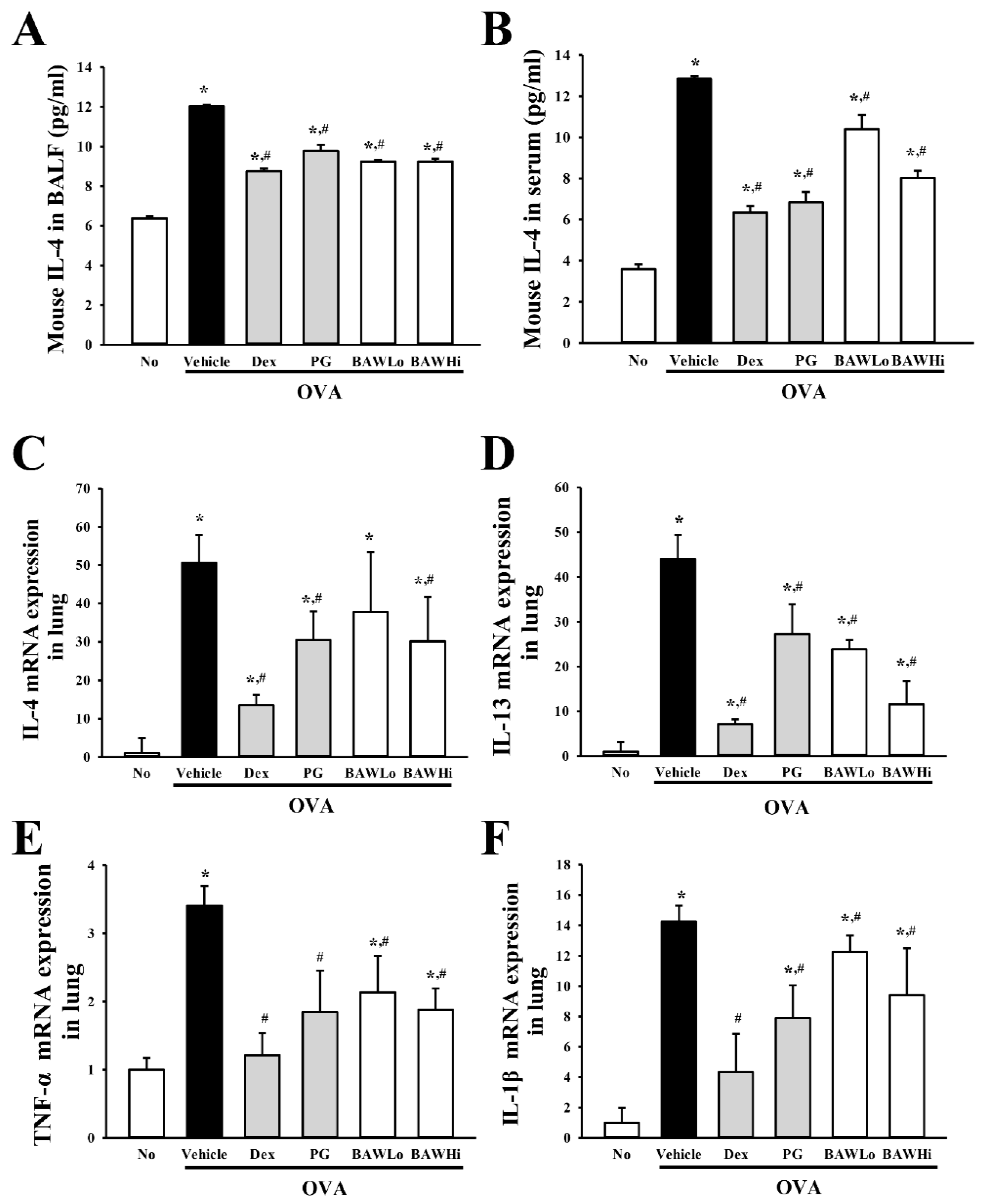
3.5. Alteration in the Expression of Key Cytokines in OVA-Induced Asthma Model Treated with BAW
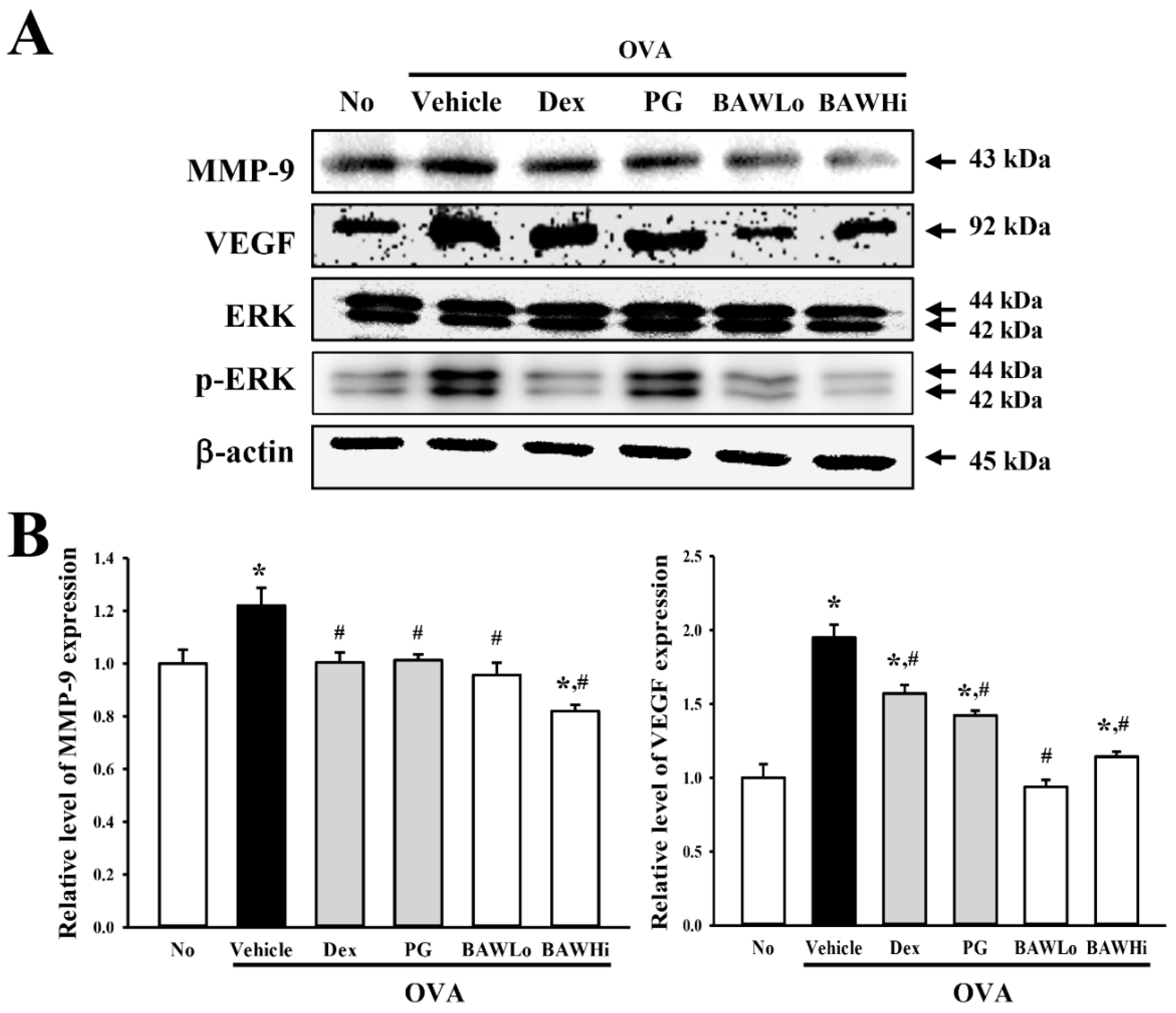
3.6. Alteration in Airway Remodeling of OVA-Induced Asthma Model Treated with BAW
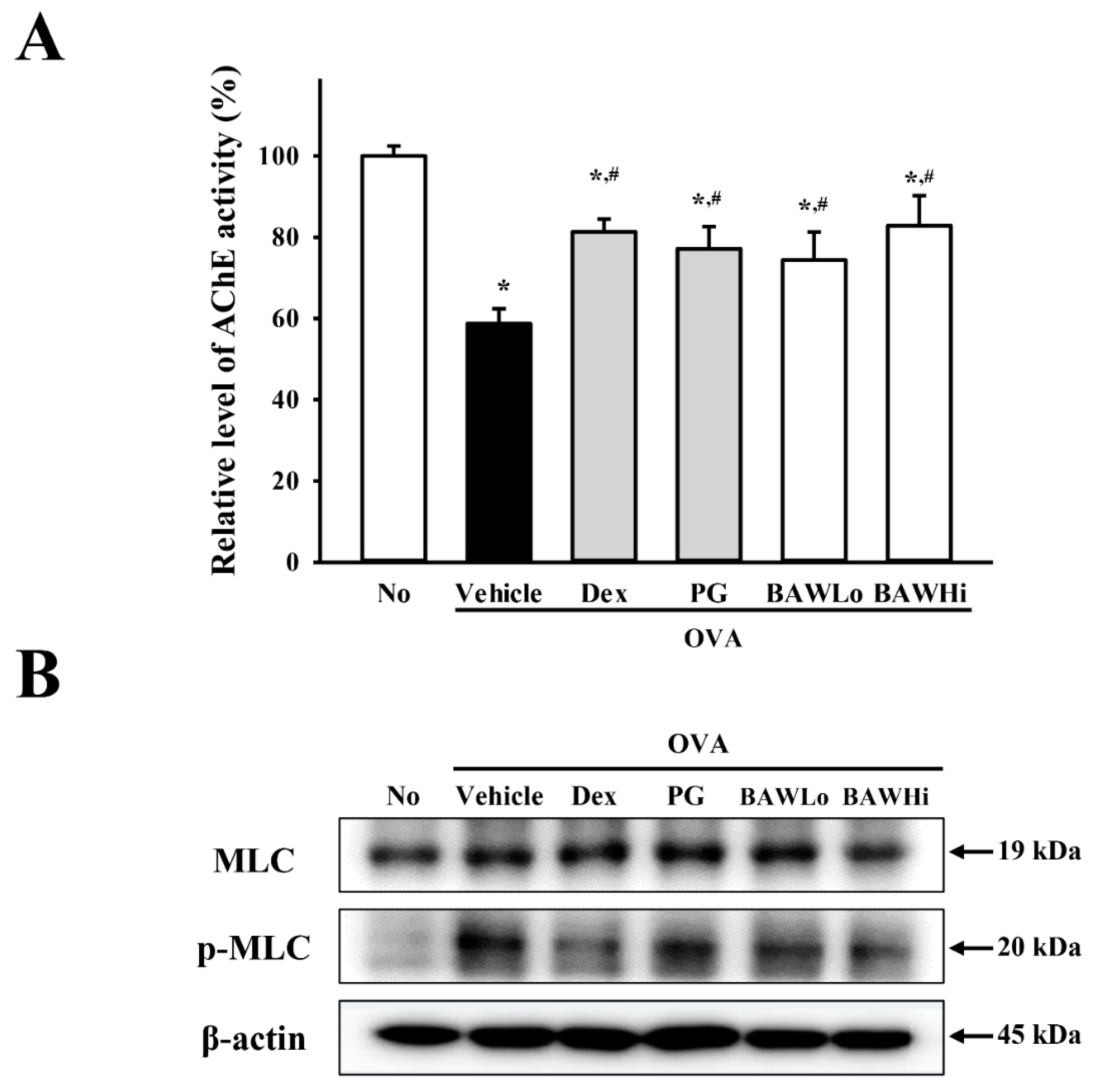
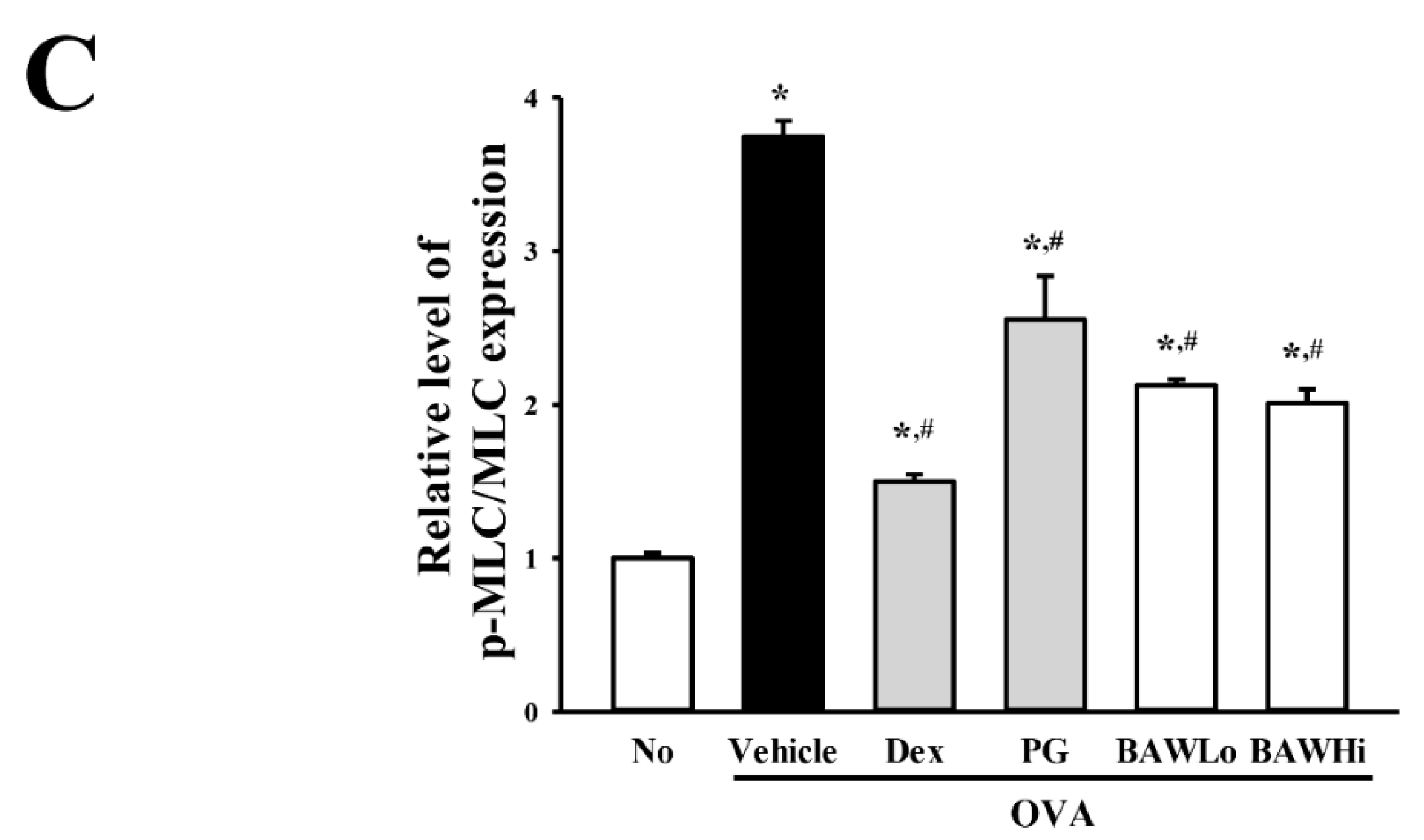
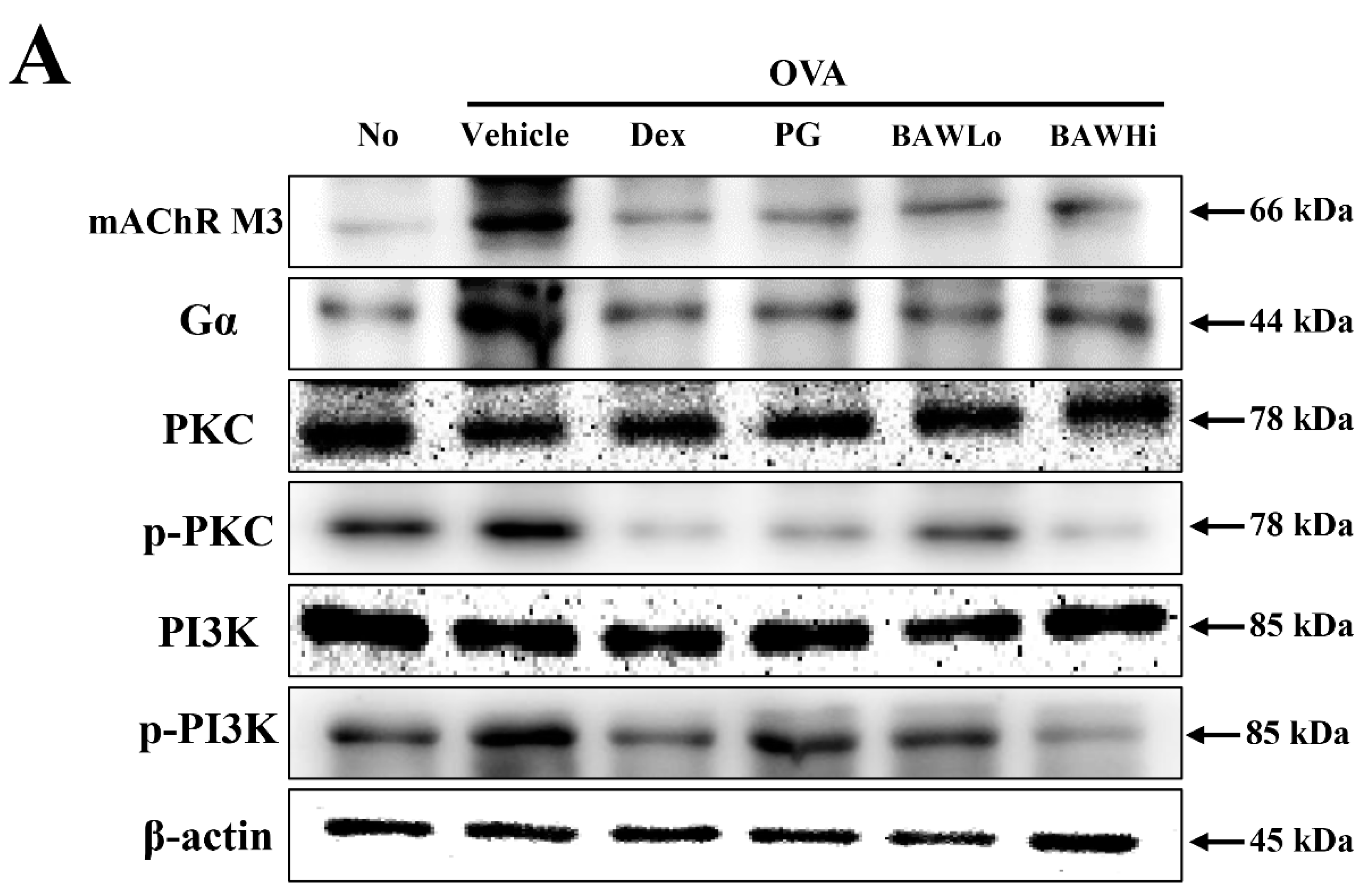
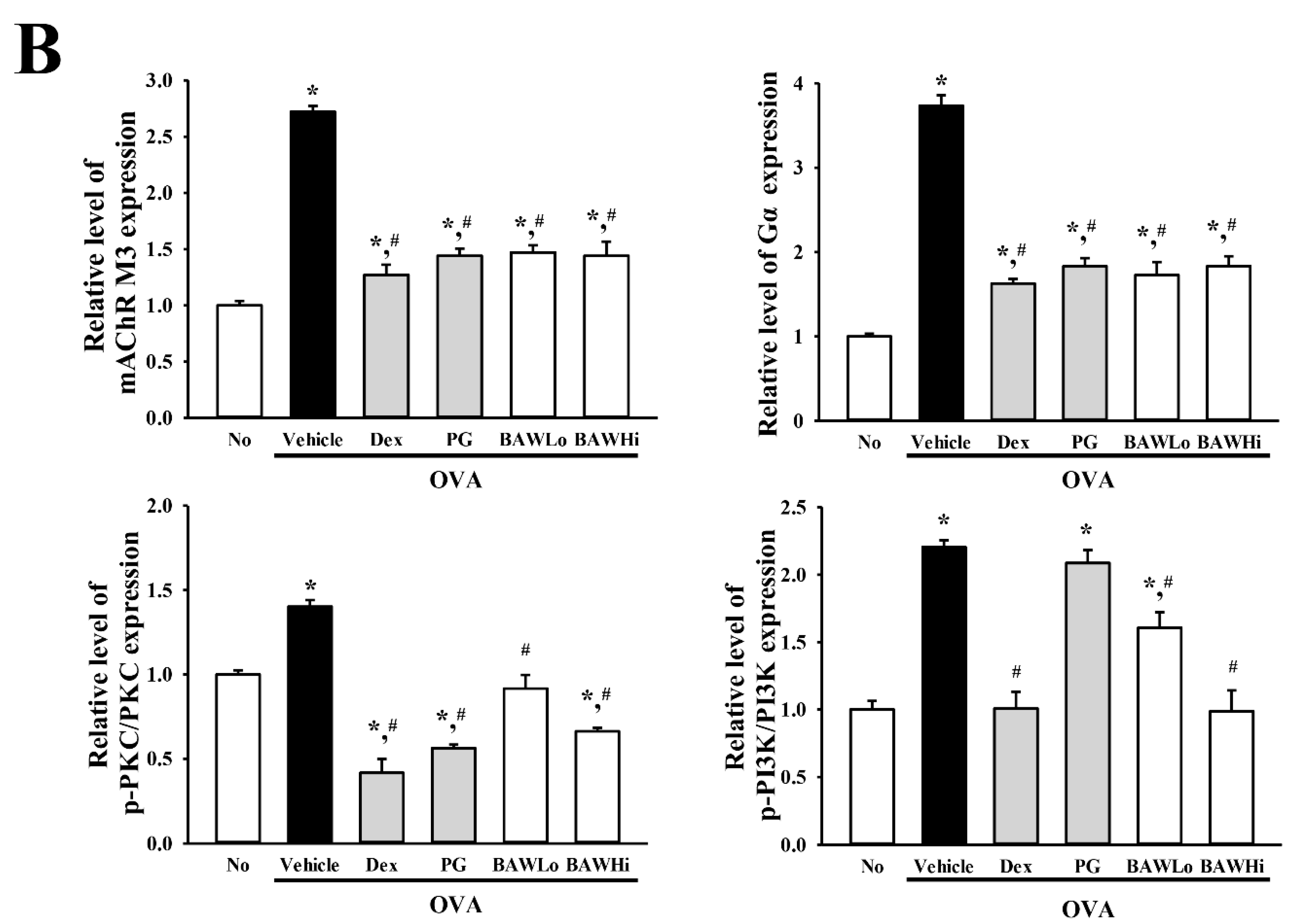
3.7. The Mechanism of BAW Action on Cholinergic Regulation of Airway Inflammation and Remodeling
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Endo, Y.; Hirahara, K.; Yagi, R.; Tumes, D.J.; Nakayama, T. Pathogenic memory type Th2 cells in allergic inflammation. Trends Immunol. 2014, 35, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.L. Antilipid agents may provide allergy protection. Ann. Allergy Asthma Immunol. 2013, 110, 1. [Google Scholar] [CrossRef] [PubMed]
- Porter, P.C.; Yang, T.; Luong, A.; Delclos, G.L.; Abramson, S.L.; Kheradmand, F.; Corry, D.B. Proteinases as molecular adjuvants in allergic airway disease. Biochim. Biophys. Acta 2011, 1810, 1059–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wüthrich, B. Epidemiology of the allergic diseases: Are they really on the increase? Int. Arch. Allergy Appl. Immunol. 1989, 90, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.R.; Lane, S.J.; Cidlowski, J.A.; Staynov, D.Z.; Lee, T.H. Glucocorticoid resistance in asthma is associated with elevated in vivo expression of the glucocorticoid receptor β-isoform. J. Allergy Clin. Immunol. 2000, 105, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, S.; Higashi, Y.; Sato, N.; Sasakawa, T.; Sengoku, T.; Ohkubo, Y.; Amaya, T.; Goto, T. Tacrolimus suppressed the production of cytokines involved in atopic dermatitis by direct stimulation of human PBMC system (comparison with steroids). Int. Immunopharmacol. 2001, 1, 1219–1226. [Google Scholar] [CrossRef]
- Schafer-Korting, M.; Schmid, M.H.; Korting, H.C. Topical glucocorticoids with improved risk-benefit ratio. Drug Saf. 1996, 14, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Simons, F.E.R. The antiallergic effects of antihistamines (H1-receptor antagonists). J. Allergy Clin. Immunol. 1992, 90, 705–715. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, W.Y.; Shin, I.S.; Shin, H.K.; Lee, M.Y. Samsoeum water extract attenuates allergic airway inflammation via modulation of Th1/Th2 cytokines and decrease of iNOS expression in asthmatic mice. BMC Complement. Altern. Med. 2015, 15, 47. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Bang, M.A.; Kim, G.; Cho, S.S.; Park, D.H. Erythronium japonicum attenuates histopathological lung abnormalities in a mouse model of ovalbumin-induced asthma. Int. J. Mol. Med. 2016, 37, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.H.; Bui, T.T.; Song, C.H.; Shin, H.S.; Shon, D.H.; Chai, O.H. Trigonella foenum-graecum alleviates airway inflammation of allergic asthma in ovalbumin-induced mouse model. Biochem. Biophys. Res. Commun. 2017, 482, 1284–1288. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.I.G.; Rios-Santos, F.; Balogun, S.O.; Almeida, D.A.T.; Damazo, A.S.; Cruz, T.C.D.; Pavan, E.; Barbosa, R.D.S.; Alvim, T.D.C.; Soares, I.M.; et al. Hydroethanolic extract from Echinodorus scaber Rataj leaves inhibits inflammation in ovalbumin-induced allergic asthma. J. Ethnopharmacol. 2017, 203, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Zemmouri, H.; Sekiou, O.; Ammar, S.; Feki, A.; Bouaziz, M.; Messarah, M.; Boumendjel, A. Urtica dioica attenuates ovalbumin-induced inflammation and lipid peroxidation of lung tissues in rat asthma model. Pharm. Biol. 2017, 55, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, L.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Enhancement of the antioxidant capacity of soy whey by fermentation with Lactobacillus plantarum B1–6. J. Funct. Foods 2015, 12, 33–44. [Google Scholar] [CrossRef]
- Kim, H.; Lee, E.; Lim, T.; Jung, J.; Lyu, Y. Inhibitory effect of Asparagus cochinchinensis on tumor necrosis factor-alpha secretion from astrocytes. Int. J. Immunopharmacol. 1998, 20, 153–162. [Google Scholar] [CrossRef]
- Lee, D.Y.; Choo, B.K.; Yoon, T.; Cheon, M.S.; Lee, H.W.; Lee, A.Y.; Kim, H.K. Anti-inflammatory effects of Asparagus cochinchinensis extract in acute and chronic cutaneous inflammation. J. Ethnopharmacol. 2009, 121, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.E.; Lee, H.A.; Kim, J.E.; Go, J.; Seo, E.J.; Yun, W.B.; Kim, D.S.; Son, H.J.; Lee, C.Y.; Lee, H.S.; et al. Therapeutic effect of ethyl acetate extract from Asparagus cochinchinensis on phthalic anhydride-induced skin inflammation. Lab. Anim. Res. 2016, 32, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Choi, H.L.; Park, S.; Lee, G.; Kim, M.; Min, J.K.; Min, B.I.; Bae, H. The effects of the standardized herbal formula PM014 on pulmonary inflammation and airway responsiveness in a murine model of cockroach allergen-induced asthma. J. Ethnopharmacol. 2014, 155, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.E.; Lee, H.A.; Kim, J.E.; Yun, W.B.; An, B.S.; Yang, S.Y.; Kim, D.S.; Lee, C.Y.; Lee, H.S.; Bae, C.J.; et al. Saponin-enriched extract of Asparagus cochinchinensis alleviates airway inflammation and remodeling in ovalbumin-induced asthma model. Int. J. Mol. Med. 2017, 40, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Song, B.R.; Kim, H.R.; Kim, J.E.; Yun, W.B.; Park, J.J.; Lee, M.L.; Choi, J.Y.; Lee, H.S.; Hwang, D.Y. Butanol extracts of Asparagus cochinchinensis fermented with Weissella cibaria inhibit iNOS-mediated COX-2 induction pathway and inflammatory cytokines in LPS-stimulated RAW264.7 macrophage cells. Exp. Ther. Med. 2017, 14, 4986–4994. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Ko, E.K.; Kim, D.H.; Jang, K.K.; Park, S.E.; Lee, H.S.; Kim, Y.C. Secoiridoid glucosides with free radical scavenging activity from the leaves of Syringa dilatata. Phytother. Res. 2003, 17, 417–419. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, K.Y.; Lee, M.Y.; Jung, D.; Cho, E.S.; Son, H.Y. Antioxidant and antiasthmatic effects of saucerneol D in a mouse model of airway inflammation. Int. Immunopharmacol. 2011, 11, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Zhou, E.; Fu, Y.; Wei, Z.; Yu, Y.; Zhang, X.; Yang, Z. Thymol attenuates allergic airway inflammation in ovalbumin (OVA)-induced mouse asthma. Fitoterapia 2014, 96, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Shin, I.S.; Lim, H.S.; Seo, C.S.; Ha, H.; Shin, H.K. Kochia scoparia fruit attenuates allergic airway inflammation in ovalbumin (OVA)-induced murine asthma model. Inhal. Toxicol. 2011, 23, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, E.J.; Park, M.N.; Kim, J.E.; Kim, S.C.; Jeung, E.B.; Lee, G.S.; Hwang, D.Y.; An, B.S. The adverse effect of 4-tert-octylphenol on fat metabolism in pregnant rats via regulation of lipogenic proteins. Environ. Toxicol. Pharmacol. 2015, 40, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Patel, K.D. Matrix metalloproteinase-2 (MMP-2) and MMP-9 in pulmonary pathology. Exp. Lung Res. 2005, 31, 599–621. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, K.; Yoshida, T. Physiological state control of fermentation processes. Biotechnol. Bioeng. 1989, 33, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Stanbury, P.F.; Whiitaker, A.; Hall, S.J. Principles of Fermentation Technology, 2nd ed.; Butterworth-Heinemann: Oxford, UK, 1995; pp. 35–122. [Google Scholar]
- Bae, E.A.; Min, S.W.; Lee, B.; Kim, N.J.; Baek, N.I.; Han, E.J.; Chung, H.G.; Kim, D.H. Antiasthmic effect of fermented Artemisia princeps in asthmic mice induced by ovalbumin. J. Microbiol. Biotechnol. 2007, 17, 1554–1557. [Google Scholar] [PubMed]
- Do, J.S.; Seo, H.J.; Hwang, J.K.; Kim, J.H.; Nam, S.Y. Effective microorganism fermentation extract (EM-X) attenuates airway hyperreactivity and inflammation through selective inhibition of the TH2 response independently of antioxidant activity. Int. J. Mol. Med. 2007, 20, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.; Busse, W.W. The eosinophil-quo vadis? J. Allergy Clin. Immunol. 2004, 113, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Camateros, P.; Tamaoka, M.; Hassan, M.; Marino, R.; Moisan, J.; Marion, D.; Guiot, M.C.; Martin, J.G.; Radzioch, D. Chronic asthma-induced airway remodeling is prevented by toll-like receptor-7/8 ligand S28463. Am. J. Respir. Crit. Care Med. 2007, 175, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Higa, T.; Ke, B. Clinical and Basic Medical Research on EMX, 1st ed.; EMRO: Okinawa, Japan, 2001; pp. 1–3. [Google Scholar]
- Ngoc, P.L.; Gold, D.R.; Tzianabos, A.O.; Weiss, S.T.; Celedon, J.C. Cytokines, allergy, and asthma. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Li-Weber, M.; Krammer, P.H. Regulation of IL4 gene expression by T cells and therapeutic perspectives. Nat. Rev. Immunol. 2003, 3, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Makinde, T.; Murphy, R.F.; Agrawal, D.K. The regulatory role of TGF-beta in airway remodeling in asthma. Immunol. Cell Biol. 2007, 85, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.G.; Duguet, A.; Eidelman, D.H. The contribution of airway smooth muscle to airway narrowing and airway hyperresponsiveness in disease. Eur. Respir. J. 2000, 16, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Guisa, J.M.; Powers, C.; File, D.; Cochrane, E.; Jimenez, N.; Debley, J.S. Airway epithelial cells from asthmatic children differentially express proremodeling factors. J. Allergy Clin. Immunol. 2012, 129, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, H.; Yilmaz, O.; Karaman, M.; Bagriyanik, H.A.; Firinci, F.; Kiray, M. Role of vascular endothelial growth factor antagonism on airway remodeling in asthma. Ann. Allergy Asthma Immunol. 2013, 110, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Kim, Y.; Lee, E.; Park, S.; Jung, K.H.; Gu, M.J.; Hong, S.P.; Kim, J. Vitex rotundifolia L. prevented airway eosinophilic inflammation and airway remodeling in an ovalbumin-induced asthma mouse model. Int. Immunol. 2013, 25, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Tian, B.P.; Zhang, L.H.; Hua, W.; Xia, L.X.; Chen, Z.H.; Li, W.; Shen, H.H. Prevention of allergic airway hyperresponsiveness and remodeling in mice by Astragaliradix antiasthmatic decoction. BMC Complement. Altern. Med. 2013, 13, 369. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Shin, I.S.; Jeon, W.Y.; Shin, N.; Shin, H.K. Bangpungtongseong-san, a traditional herbal medicine, attenuates chronic asthmatic effects induced by repeated ovalbumin challenge. Int. J. Mol. Med. 2014, 33, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, L.H.; Wu, Y.F.; Lai, T.W.; Wang, H.S.; Xiao, H.; Che, L.Q.; Ying, S.M.; Li, W.; Chen, Z.H.; et al. Suhuang antitussive capsule at lower doses attenuates airway hyperresponsiveness, inflammation, and remodeling in a murine model of chronic asthma. Sci. Rep. 2016, 6, 21515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosens, R.; Zaagsma, J.; Meurs, H.; Halayko, A.J. Muscarinic receptor signaling in the pathophysiology of asthma and COPD. Respir. Res. 2006, 7, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Rack´e, K.; Matthiesen, S. The airway cholinergic system: Physiology and pharmacology. Pulm. Pharmacol. Ther. 2004, 17, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Wess, J.; Eglen, R.M.; Gautam, D. Muscarinic acetylcholine receptors: Mutant mice provide new insights for drug development. Nat. Rev. Drug Discov. 2007, 6, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, K.E. Cholinergic pathways in the lungs and anticholinergic therapy for chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005, 2, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Profita, M.; Di Giorgi, R.; Sala, A.; Bonanno, A.; Riccobono, L.; Mirabella, F.; Gjomarkaj, M.; Bonsignore, G.; Bousquet, J.; Vignola, A.M. Muscarinic receptors, leukotriene B4 production and neutrophilic inflammation in COPD patients. J. Allergy 2005, 60, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Profita, M.; Bonanno, A.; Siena, L.; Bruno, A.; Ferraro, M.; Montalbano, A.M.; Albano, G.D.; Riccobono, L.; Casarosa, P.; Pieper, M.P.; et al. Smoke, choline acetyltransferase, muscarinic receptors, and fibroblast proliferation in chronic obstructive pulmonary disease. J. Pharmacol. Exp. Ther. 2009, 329, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Bos, I.S.T.; Gosens, R.; Zuidhof, A.B.; Schaafsma, D.; Halayko, A.J.; Meurs, H.; Zaagsma, J. Inhibition of allergen-induced airway remodelling by tiotropium and budesonide: A comparison. Eur. Respir. J. 2007, 30, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Kistemaker, L.E.; Bos, S.T.; Mudde, W.M.; Hylkema, M.N.; Hiemstra, P.S.; Wess, J.; Meurs, H.; Kerstjens, H.A.; Gosens, R. Muscarinic m3 receptors contribute to allergen-induced airway remodeling in mice. Am. J. Respir. Cell Mol. Biol. 2014, 50, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Gosens, R.; Bos, I.S.T.; Zaagsma, J.; Meurs, H. Protective effects of tiotropium bromide in the progression of airway smooth muscle remodeling. Am. J. Respir. Crit. Care Med. 2005, 171, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Lee, E.B. Pharmacological studies on Platycodon grandiflorum A. DC. 3. Activities of crude platycodin on respiratory and circulatory systems and its other pharmacological activities. Yakugaku Zasshi 1972, 92, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.B. Pharmacological studies on Platycodon grandiflorum A. DC. IV. A comparison of experimental pharmacological effects of crude platycodin with clinical indications of Platycodi radix. Yakugaku Zasshi 1973, 93, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.C.; Kang, O.H.; Choi, J.G.; Lee, Y.S.; Brice, O.O.; Jung, H.J.; Hong, S.H.; Lee, Y.M.; Shin, D.W.; Kim, Y.S.; et al. Anti-allergic activity of a Platycodon root ethanol extract. Int. J. Mol. Sci. 2010, 11, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yang, S.; Du, J.; Jinfu, Y.; Shumin, W. Platycodin D attenuates airway inflammation in a mouse model of allergic asthma by regulation NF-κB pathway. Inflammation 2015, 38, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Hur, Y.G.; Kim, W.G.; Park, B.W.; Ahn, K.S.; Kim, J.J.; Bae, H. Inhibitory effect of Platycodon grandiflorum on T(H)1 and T(H)2 immune responses in a murine model of 2,4-dinitrofluorobenzene-induced atopic dermatitis-like skin lesions. Ann. Allergy Asthma Immunol. 2011, 106, 54–61. [Google Scholar] [CrossRef] [PubMed]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.Y.; Kim, J.E.; Park, J.J.; Lee, M.R.; Song, B.R.; Park, J.W.; Kang, M.J.; Lee, H.S.; Son, H.J.; Hong, J.T.; et al. The Anti-Inflammatory Effects of Fermented Herbal Roots of Asparagus cochinchinensis in an Ovalbumin-Induced Asthma Model. J. Clin. Med. 2018, 7, 377. https://doi.org/10.3390/jcm7100377
Choi JY, Kim JE, Park JJ, Lee MR, Song BR, Park JW, Kang MJ, Lee HS, Son HJ, Hong JT, et al. The Anti-Inflammatory Effects of Fermented Herbal Roots of Asparagus cochinchinensis in an Ovalbumin-Induced Asthma Model. Journal of Clinical Medicine. 2018; 7(10):377. https://doi.org/10.3390/jcm7100377
Chicago/Turabian StyleChoi, Jun Young, Ji Eun Kim, Jin Ju Park, Mi Rim Lee, Bo Ram Song, Ji Won Park, Mi Ju Kang, Hee Seob Lee, Hong Joo Son, Jin Tae Hong, and et al. 2018. "The Anti-Inflammatory Effects of Fermented Herbal Roots of Asparagus cochinchinensis in an Ovalbumin-Induced Asthma Model" Journal of Clinical Medicine 7, no. 10: 377. https://doi.org/10.3390/jcm7100377