Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study)

 , ,
, ,
Abstract
:1. Introduction
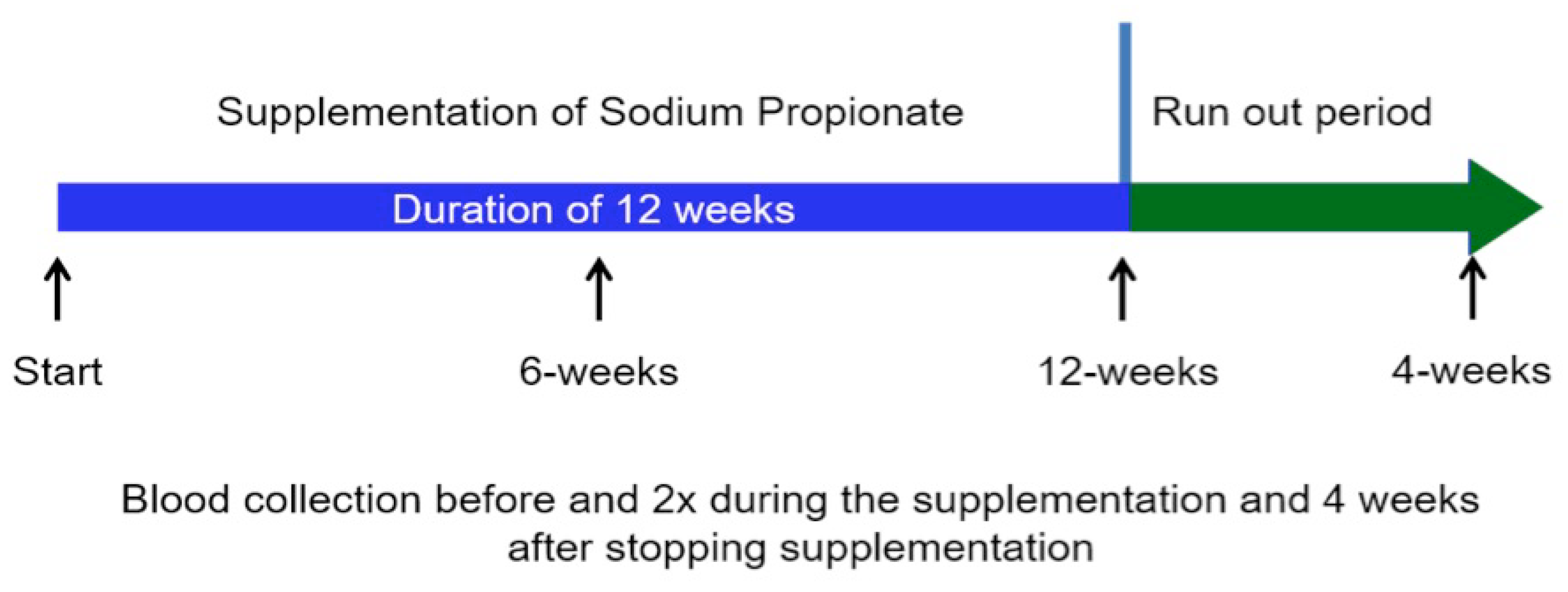
2. Subjects and Methods
2.1. Biochemical Measurements
2.2. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vaziri, N.D. Oxidative stress in uremia: Nature, mechanisms, and potential consequences. Semin. Nephrol. 2004, 24, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Andersen, K.; Stecher, B. The intestinal microbiota, a leaky gut, and abnormal immunity in kidney disease. Kidney Int. 2013, 83, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- de Boer, I.H.; Zelnick, L.; Afkarian, M.; Ayers, E.; Curtin, L.; Himmelfarb, J.; Ikizler, T.A.; Kahn, S.E.; Kestenbaum, B.; Utzschneider, K. Impaired glucose and insulin homeostasis in moderate-severe chronic kidney disease. J. Am. Soc. Nephrol. 2016, 27, 2861–2871. [Google Scholar] [PubMed]
- Vaziri, N.D.; Pahl, M.V.; Crum, A.; Norris, K. Effect of uremia on structure and function of immune system. J. Ren. Nutr. 2012, 22, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Miele, L.; Giorgio, V.; Alberelli, M.A.; De Candia, E.; Gasbarrini, A.; Grieco, A. Impact of gut microbiota on obesity, diabetes, and cardiovascular disease risk. Curr. Cardiol. Rep. 2015, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.H.; May, C.; Wilck, N.; et al. Dietary fatty acids directly impact central nervous system autoimmunity via the small intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Lun, H.; Yang, W.; Zhao, S.; Jiang, M.; Xu, M.; Liu, F.; Wang, Y. Altered gut microbiota and microbial biomarkers associated with chronic kidney disease. Microbiologyopen 2018. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; De Santis, T.Z.; Ni, Z.; Nguyen, T.H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.L.; Savoj, J.; Nakata, M.B.; Vaziri, N.D. Altered microbiome in chronic kidney disease: Systemic effects of gut-derived uremic toxins. Clin. Sci. 2018, 132, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Zhao, Y.Y.; Pahl, M.V. Altered intestinal microbial flora and impaired epithelial barrier structure and function in ckd: The nature, mechanisms, consequences and potential treatment. Nephrol. Dial. Transplant. 2016, 31, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.; Kesper, M.S.; Marschner, J.A.; Konrad, L.; Ryu, M.; Kumar, Vr.S.; Kulkarni, O.P.; Mulay, S.R.; Romoli, S.; Demleitner, J.; et al. Intestinal Dysbiosis, Barrier Dysfunction, and Bacterial Translocation Account for CKD-Related Systemic Inflammation. J. Am. Soc. Nephrol. 2017, 28, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Aronov, P.A.; Luo, F.J.; Plummer, N.S.; Quan, Z.; Holmes, S.; Hostetter, T.H.; Meyer, T.W. Colonic contribution to uremic solutes. J. Am. Soc. Nephrol. 2011, 22, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
- Barreto, F.C.; Barreto, D.V.; Liabeuf, S.; Meert, N.; Glorieux, G.; Temmar, M.; Choukroun, G.; Vanholder, R.; Massy, Z.A.; European Uremic Toxin Work, G. Serum indoxyl sulfate is associated with vascular disease and mortality in chronic kidney disease patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, B.R.; Marzocco, S.; Nardone, L.; Sirico, M.L.; De Simone, E.; Di Natale, G.; Di Micco, L. Urea and impairment of the gut-kidney axix in chronic kidney disease. G. Ital. Nefrol. 2017, 34, 1–14. [Google Scholar]
- Cosola, C.; Rocchetti, M.T.; Sabatino, A.; Fiaccadori, E.; Di Iorio, B.R. Micorbiota issue in CKD: How promising are gut-target approaches. J. Nephrol. 2018, in press. [Google Scholar]
- Lau, W.L.; Kalantar-Zadeh, K.; Vaziri, N.D. The gut as a source of inflammation in chronic kidney disease. Nephron 2015, 130, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Diamanti, A.P.; Manuela, R.M.; Laganà, B.; D’Amelio, R. Microbiota and chronic inflammatory arthritis: An interwoven link. J. Transl. Med. 2016, 14, 233. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in esrd. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Puddu, A.; Sanguineti, R.; Montecucco, F.; Viviani, G.L. Evidence for the gut microbiota short-chain fatty acids as key pathophysiological molecules improving diabetes. Mediators Inflamm. 2014, 2014, 162021. [Google Scholar] [CrossRef] [PubMed]
- Pisano, A.; D’Arrigo, G.; Coppolino, G.; Bolignano, D. Biotic supplements for renal patients: A Systematic review and meta-analysis. Nutrients 2018. [Google Scholar] [CrossRef] [PubMed]
- Pei, M.; Wei, L.; Hu, S.; Yang, B.; Si, J.; Yang, H.; Zhai, J. Probiotics, prebiotics and synbiotics for chronic kidney disease: Protocol for a systematic review and meta-analysis. BMJ Open 2018. [Google Scholar] [CrossRef] [PubMed]
- Harig, J.M.; Soergel, K.H.; Komorowski, R.A.; Wood, C.M. Treatment of diversion colitis with short-chain-fatty acid irrigation. N. Engl. J. Med. 1989, 320, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S. Microbiota replacement therapies: Innovation in gastrointestinal care. Clin. Pharmacol. Ther. 2018, 103, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Reparaz, J.; Kirby, T.O.; Kasper, L.H. The gut microbiome and multiple sclerosis. Cold Spring Harb. Perspect. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells. Immunobiology 2015, 220, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor gpr43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.L.; Yap, Y.A.; McLeod, K.H.; Mackay, C.R.; Marino, E. Dietary metabolites and the gut microbiota: An alternative approach to control inflammatory and autoimmune diseases. Clin. Transl. Immunol. 2016, 5, e82. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory t cells by suppression of histone deacetylases and regulation of the mtor-s6k pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Haga, S.; Aoyama, Y.; Kiriyama, S. Short-chain fatty acids suppress cholesterol synthesis in rat liver and intestine. J. Nutr. 1999, 129, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Raj, D.S. The gut microbiome, kidney disease, and targeted interventions. J. Am. Soc. Nephrol. 2014, 25, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, V.M.; Wei, G.; Baird, B.C.; Murtaugh, M.; Chonchol, M.B.; Raphael, K.L.; Greene, T.; Beddhu, S. High dietary fiber intake is associated with decreased inflammation and all-cause mortality in patients with chronic kidney disease. Kidney Int. 2012, 81, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenepoel, P.; Meijers, B.K. Dietary fiber and protein: Nutritional therapy in chronic kidney disease and beyond. Kidney Int. 2012, 81, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; Plummer, N.S.; Gardner, C.D.; Hostetter, T.H.; Meyer, T.W. Effect of increasing dietary fiber on plasma levels of colon-derived solutes in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2014, 9, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Bellizzi, V.; Conte, G.; Borrelli, S.; Cupisti, A.; De Nicola, L.; Di Iorio, B.R.; Cabiddu, G.; Mandreoli, M.; Paoletti, E.; Piccoli, G.B.; et al. Controversial issues in CKD clinical practice: Position statement of the CKD-treatment working group of the Italian Society of Nephrology. J. Nephrol. 2017, 30, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Furuland, H.; McEwan, P.; Evans, M.; Linde, C.; Ayoubkhani, D.; Bakhai, A.; Palaka, E.; Bennett, H.; Qin, L. Serum potassium as a predictor of adverse clinical outcomes in patients with chronic kidney disease: New risk equations using the UK clinical practice research datalink. BMC Nephrol. 2018, 19, 211. [Google Scholar] [CrossRef] [PubMed]
- Yallapragada, S.G.; Nash, C.B.; Robinson, D.T. Early-life exposure to antibiotics, alterations in the intestinal microbiome, and risk of metabolic disease in children and adults. Pediatr. Ann. 2015, 44, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Dostal, A.; Lacroix, C.; Pham, V.T.; Zimmermann, M.B.; Del’homme, C.; Bernalier-Donadille, A.; Chassard, C. Iron supplementation promotes gut microbiota metabolic activity but not colitis markers in human gut microbiota-associated rats. Br. J. Nutr. 2014, 111, 2135–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-González, J.F.; Mora-Fernández, C.; Muros de Fuentes, M.; Donate-Correa, J.; Cazaña-Pérez, V.; García-Pérez, J. Effect of phosphate binders on serum inflammatory profile, soluble CD14, and endotoxin levels in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2011, 6, 2272–2279. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Chen, X.; Xia, T.; Zhang, F.; Gao, S.; Chen, W. Lc-ms/ms method for simultaneous determination of serum p-cresyl sulfate and indoxyl sulfate in patients undergoing peritoneal dialysis. Biomed. Chromatogr. 2016, 30, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of propionic acid (E 280), sodium propionate (E 281), calcium propionate (E 282) and potassium propionate (E 283) as food additives. EFSA J. 2014, 12, 3779. [Google Scholar] [CrossRef]
- CFR-Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfCFR/CFRSearch.cfm?fr=184.1141a (accessed on 24 August 2018).
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. Tgf-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, V.; Roumeliotis, S.; Gorny, X.; Eleftheriadis, T.; Mertens, P.R. Oxidative stress in patients undergoing peritoneal dialysis: A current review of the literature. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Podrez, E.A.; Abu-Soud, H.M.; Hazen, S.L. Myeloperoxidase-generated oxidants and atherosclerosis. Free Radic. Biol. Med. 2000, 28, 1717–1725. [Google Scholar] [CrossRef]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an active disease biomarker: Recent biochemical and pathological perspectives. Med. Sci. 2018. [Google Scholar] [CrossRef]
- Stenvinkel, P.; Alvestrand, A. Inflammation in end-stage renal disease: Sources, consequences, and therapy. Semin. Dial. 2002, 15, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C.; et al. The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro. Diabetes Obes. Metab. 2017, 19, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, Z.; Hu, J.; Du, J.; Mitch, W.E. Insulin resistance accelerates muscle protein degradation: Activation of the ubiquitin-proteasome pathway by defects in muscle cell signaling. Endocrinology 2006, 147, 4160–4168. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, K.; Shoji, T.; Emoto, M.; Tahara, H.; Koyama, H.; Ishimura, E.; Miki, T.; Tabata, T.; Nishizawa, Y. Insulin resistance as an independent predictor of cardiovascular mortality in patients with end-stage renal disease. J. Am. Soc. Nephrol. 2002, 13, 1894–1900. [Google Scholar] [CrossRef] [PubMed]
- Karaboyas, A.; Morgenstern, H.; Pisoni, R.L.; Zee, J.; Vanholder, R.; Jacobson, S.H.; Inaba, M.; Loram, L.C.; Port, F.K.; Robinson, B.M. Association between serum ferritin and mortality: Findings from the USA, Japan and European dialysis outcomes and practice patterns study. Nephrol. Dial. Transplant. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kalantar-Zadeh, K.; Rodriguez, R.A.; Humphreys, M.H. Association between serum ferritin and measures of inflammation, nutrition and iron in haemodialysis patients. Nephrol. Dial. Transplant. 2004, 19, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, M.T.; Vilaca, S.S.; Carvalho, M.; Fernandes, A.P.; Dusse, L.M.; Gomes, K.B. Resistance of dialyzed patients to erythropoietin. Rev. Bras. Hematol. Hemoterapia 2015, 37, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, N.; Hori, D.; Flavahan, S.; Steppan, J.; Flavahan, N.A.; Berkowitz, D.E.; Pluznick, J.L. Microbial short chain fatty acid metabolites lower blood pressure via endothelial g protein-coupled receptor 41. Physiol. Genomics. 2016, 48, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L. Microbial short-chain fatty acids and blood pressure regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Vanholder, R.; Schepers, E.; Pletinck, A.; Nagler, E.V.; Glorieux, G. The uremic toxicity of indoxyl sulfate and p-cresyl sulfate: A systematic review. J. Am. Soc. Nephrol. 2014, 25, 1897–1907. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; Glorieux, G.; Meijers, B. P-cresol sulfate and indoxyl sulfate: Some clouds are gathering in the uremic toxin sky. Kidney Int. 2017, 92, 1323–1324. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; Lin, C.; Cecelja, M.; Mangino, M.; Matey-Hernandez, M.L.; Keehn, L.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Kuo, C.F.; Chowienczyk, P.; Valdes, A.M. Gut microbial diversity is associated with lower arterial stiffness in women. Eur. Heart J. 2018, 39, 2390–2397. [Google Scholar] [CrossRef] [PubMed]
- Meijers, B.K.; De Loor, H.; Bammens, B.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. P-cresyl sulfate and indoxyl sulfate in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 1932–1938. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.C.; Kuo, K.L.; Wu, C.C.; Tarng, D.C. Indoxyl sulfate: A novel cardiovascular risk factor in chronic kidney disease. J. Am. Heart Assoc. 2017. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The orphan g protein-coupled receptors gpr41 and gpr43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.S.; Anderson, J.W.; Bridges, S.R. Propionate inhibits hepatocyte lipid synthesis. Proc. Soc. Exp. Biol. Med. 1990, 195, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Boillot, J.; Alamowitch, C.; Berger, A.M.; Luo, J.; Bruzzo, F.; Bornet, F.R.; Slama, G. Effects of dietary propionate on hepatic glucose production, whole-body glucose utilization, carbohydrate and lipid metabolism in normal rats. Br. J. Nutr. 1995, 73, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, T.; Loo, R.L.; Anastasovska, J.; Gibson, G.R.; Tuohy, K.M.; Sharma, R.K.; Swann, J.R.; Deaville, E.R.; Sleeth, M.L.; Thomas, E.L.; et al. Differential effects of two fermentable carbohydrates on central appetite regulation and body composition. PLoS ONE 2012, 7, e43263. [Google Scholar] [CrossRef] [PubMed]
- Correa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Sunkara, L.T.; Jiang, W.; Bible, M.; Carter, S.; Ma, X.; Qiao, S.; Zhang, G. Induction of porcine host defense peptide gene expression by short-chain fatty acids and their analogs. PLoS ONE 2013, 8, e72922. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.C.; Garner, C.D.; Slauch, J.M.; Dwyer, Z.W.; Lawhon, S.D.; Frye, J.G.; McClelland, M.; Ahmer, B.M.; Altier, C. The intestinal fatty acid propionate inhibits salmonella invasion through the post-translational control of hild. Mol. Microbiol. 2013, 87, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Oliveira, V.; Amano, M.T.; Correa-Costa, M.; Castoldi, A.; Felizardo, R.J.; de Almeida, D.C.; Bassi, E.J.; Moraes-Vieira, P.M.; Hiyane, M.I.; Rodas, A.C.; et al. Gut bacteria products prevent aki induced by ischemia-reperfusion. J. Am. Soc. Nephrol. 2015, 26, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Sunkara, L.T.; Jiang, W.; Zhang, G. Modulation of antimicrobial host defense peptide gene expression by free fatty acids. PLoS ONE 2012, 7, e49558. [Google Scholar] [CrossRef] [PubMed]
- Tayebi Khosroshahi, H.; Vaziri, N.D.; Abedi, B.; Asl, B.H.; Ghojazadeh, M.; Jing, W.; Vatankhah, A.M. Effect of high amylose resistant starch (ham-rs2) supplementation on biomarkers of inflammation and oxidative stress in hemodialysis patients: A randomized clinical trial. Hemodial. Int. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Liu, S.M.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Farzaneh, S.H.; Kieffer, D.A.; Adams, S.H.; Martin, R.J. High amylose resistant starch diet ameliorates oxidative stress, inflammation, and progression of chronic kidney disease. PLoS ONE 2014, 9, e114881. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Tsukushi, S.; Ise, M.; Miyazaki, T.; Tsubakihara, Y.; Owada, A.; Shiigai, T. Indoxyl sulfate and progression of renal failure: Effects of a low-protein diet and oral sorbent on indoxyl sulfate production in uremic rats and undialyzed uremic patients. Miner. Electrolyte Metab. 1997, 23, 179–184. [Google Scholar] [PubMed]
- Vaziri, N.D.; Yuan, J.; Khazaeli, M.; Masuda, Y.; Ichii, H.; Liu, S.M. Oral activated charcoal adsorbent (ast-120) ameliorates chronic kidney disease-induced intestinal epithelial barrier disruption. Am. J. Nephrol. 2013, 37, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Schulman, G.; Berl, T.; Beck, G.J.; Remuzzi, G.; Ritz, E.; Arita, K.; Kato, A.; Shimizu, M. Randomized placebo-controlled eppic trials of ast-120 in ckd. J. Am. Soc. Nephrol. 2015, 26, 1732–1746. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameter | Mean ± SD |
| Age (years) | 69.9 ± 11.2 |
| Gender (male/female) | 11/9 |
| Height (cm) | 166 ± 6 |
| Weight (kg) | 64.8 ± 13.5 |
| BMI (kg/m2) | 26.1 ± 2.5 |
| Dialysis vintage (months) | 45 ± 19 |
| Causes of renal failure | n (%) |
| Chronic glomerulonephritis | 7 (35) |
| Polycystic kidney disease | 2 (10) |
| Chronic pyelonephritis | 2 (10) |
| Diabetes mellitus | 9 (45) |
| Comorbidities | n (%) |
| Hypertension | 18 (90) |
| Cerebral vasculopathy | 4 (20) |
| Peripheral vasculopathy | 11 (55) |
| Recurrent cystitis | 1 (5) |
| Chronic obstructive pulmonary disease | 2 (10) |
| Start | 6 Weeks | 12 Weeks | 16 Weeks | p | |
|---|---|---|---|---|---|
| Body weight (kg) | 64.8 ± 13.5 | 64.2 ± 13.5 | 64.3 ± 13.0 | 64.5 ± 13.2 | NS |
| Systolic blood pressure (mm Hg) | 144 ± 39 | 135 ± 41 | 130 ± 40 | 140 ± 49 | NS |
| Diastolic blood pressure (mm Hg) | 70 ± 11 | 67 ± 11 | 69 ± 12 | 72 ± 13 | NS |
| Creatinine (mg/dL) | 7.6 ± 2.5 | 7.7 ± 2.6 | 7.7 ± 2.9 | 7.8 ± 2.0 | NS |
| Urea (mg/dL) | 131 ± 42 | 129 ± 45 | 127 ± 55 | 133 ± 52 | NS |
| Uric acid (mg/dL) | 5.2 ± 1.4 | 5.4 ± 1.6 | 5.5 ± 1.7 | 5.2 ± 1.9 | NS |
| Na (mmol/L) | 137 ± 3 | 138 ± 3 | 139 ± 5 | 139 ± 6 | NS |
| K (mmol/L) | 5.4 ± 0.9 | 5.1 ± 0.8 | 5.0 ± 1.1 | 5.2 ± 1.0 | NS |
| Ca (mg/dL) | 8.9 ± 1.0 | 9.0 ± 0.7 | 9.1 ± 0.8 | 9.0 ± 0.9 | NS |
| Phosphate (mg/dL) | 4.4 ± 1.5 | 4.4 ± 2.0 | 4.2 ± 1.9 | 4.3 ± 2.1 | NS |
| Alkaline phosphatase (U/L) | 120 ± 69 | 134 ± 74 | 130 ± 70 | 142 ± 81 | NS |
| PTH intact (ng/L) | 259 ± 148 | 330 ± 152 | 301 ± 132 | 288 ± 178 | NS |
| Bicarbonate (mmol/L) | 22.6 ± 2.6 | 23.0 ± 2.7 | 23.0 ± 2.2 | 23.0 ± 2.5 | NS |
| Albumin (g/dL) | 3.6 ± 0.4 | 3.8 ± 0.3 | 3.8 ± 0.4 | 3.7 ± 0.5 | NS |
| LDL-cholesterol (mg/dL) | 116 ± 21 | 110 ± 24 | 102 ± 42 | 110 ± 31 | NS |
| Triglycerides (mg/dL) | 191 ± 103 | 195 ± 98 | 189 ± 102 | 199 ± 123 | NS |
| Hemoglobin (g/dL) | 11.6 ± 1.4 | 12.2 ± 1.5 | 12.3 ± 1.4 | 12.0 ± 1.8 | NS |
| Serum iron (µg/dL) | 51 ± 27 | 55 ± 22 | 55 ± 22 | 48 ± 33 | NS |
| Transferrin (g/L) | 180 ± 44 | 204 ± 53 | 221 ± 73 | 170 ± 49 | NS |
| TSAT (%) | 22 ± 15 | 39 ± 18 | 40 ± 20 | 25 ± 19 | 0.002 |
| Ferritin (ng/mL) | 226 ± 258 | 63 ± 80 | 73 ± 77 | 212 ± 188 | 0.01 |
| Glucose (mg/dL) | 124 ± 18 | 121 ± 16 | 120 ± 19 | 125 ± 20 | NS |
| HbA1 c (%) | 6.6 ± 1.2 | 6.7 ± 1.2 | 6.6 ± 1.3 | 6.7 ± 1.4 | NS |
| Insulin (IU/mL) | 27 ± 26 | 19 ± 15 | 20 ± 15 | 29 ± 28 | 0.024 |
| HOMA-Index | 11.7 ± 13.8 | 8.3 ± 7.4 | 8.1 ± 7.0 | 12.1 ± 9.6 | 0.041 |
| Kt/V | 1.31 ± 0.6 | 1.29 ± 0.3 | 1.32 ± 0.4 | 1.30 ± 0.5 | NS |
| nPCR | 1.10 ± 0.15 | 1.16 ± 0.29 | 1.18 ± 0.32 | 1.11 ± 0.19 | NS |
| Start | 6 Weeks | 12 Weeks | 16 Weeks | p | |
|---|---|---|---|---|---|
| hs-C-reactive Protein (mg/L) | 5.98 ± 3.7 | 3.6 ± 6.3 | 3.2 ± 4.3 | 6.0 ± 4.2 | 0.003 |
| IL-2 (pg/mL) | 7.79 ± 2.3 | 6.16 ± 2.22 | 5.69 ± 2.02 | 7.95 ± 3.1 | <0.007 |
| IL-6 (pg/mL) | 2.06 ± 1.36 | 1.79 ± 0.94 | 1.72 ± 0.84 | 2.1 ± 1.5 | NS |
| IL-10 (pg/mL) | 1.57 ± 0.62 | 2.59 ± 1.32 | 2.69 ± 1.23 | 1.47 ± 0.81 | <0.001 |
| IL-17 (pg/mL) | 240 ± 37 | 212 ± 24 | 204 ± 17 | 244 ± 44 | <0.01 |
| TNF-α (pg/mL) | 38.8 ± 12.6 | 38.5 ± 13.5 | 37.4 ± 8.3 | 38.4 ± 12.2 | NS |
| IFN-γ (pg/mL) | 0.49 ± 0.47 | 0.39 ± 0.33 | 0.36 ± 0.18 | 0.52 ± 0.45 | NS |
| Endotoxin (EU/mL) | 1.99 ± 0.88 | 2.36 ± 0.97 | 2.3 ± 0.79 | 2.02 ± 1.02 | NS |
| TGF-β (pg/mL) | 620.5 ± 220.9 | 505.1 ± 147.8 | 495.0 ± 152.7 | 612.4 ± 214.2 | <0.05 |
| Malondialdehyde (ng/mL) | 389.2 ± 133.6 | 287.8 ± 103 | 266.8 ± 92.2 | 368.9 ± 109 | <0.01 |
| GSH-PX activity (micromole/L) | 127.2 ± 75.3 | 94.5 ± 53.5 | 92.4 ± 52.4 | 138.4 ± 102.2 | <0.01 |
| Indoxyl sulfate (mg/L) | 73.1 ± 29.3 | 58.9 ± 23.3 | 50.9 ± 24.0 | 70.7 ± 26.7 | <0.001 |
| P-cresol sulfate (mg/L) | 37.9 ± 27.3 | 24.9 ± 13.8 | 19.1 ± 12.6 | 35.5 ± 24.7 | <0.001 |
| Before SP Supplementation | After SP Supplementation | p-Value | |
|---|---|---|---|
| Physical Components | 58 ± 21 | 80 ± 19 | 0.001 |
| Physical Functions | 25 ± 15 | 33 ± 18 | 0.148 |
| Pain Tolerance | 35 ± 16 | 58 ± 17 | 0.001 |
| General Health | 27 ± 20 | 34 ± 16 | 0.152 |
| Vitality | 23 ± 10 | 45 ± 18 | 0.001 |
| Social Functions | 41 ± 16 | 70 ± 14 | 0.001 |
| Role Emotional | 33 ± 17 | 33 ± 13 | 0.948 |
| Mental Health | 29 ± 18 | 48 ± 22 | 0.005 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzocco, S.; Fazeli, G.; Di Micco, L.; Autore, G.; Adesso, S.; Dal Piaz, F.; Heidland, A.; Di Iorio, B. Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study). J. Clin. Med. 2018, 7, 315. https://doi.org/10.3390/jcm7100315
Marzocco S, Fazeli G, Di Micco L, Autore G, Adesso S, Dal Piaz F, Heidland A, Di Iorio B. Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study). Journal of Clinical Medicine. 2018; 7(10):315. https://doi.org/10.3390/jcm7100315
Chicago/Turabian StyleMarzocco, Stefania, Gholamreza Fazeli, Lucia Di Micco, Giuseppina Autore, Simona Adesso, Fabrizio Dal Piaz, August Heidland, and Biagio Di Iorio. 2018. "Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study)" Journal of Clinical Medicine 7, no. 10: 315. https://doi.org/10.3390/jcm7100315