1. Introduction
Hepatocellular carcinoma (HCC) is the fourth most common malignancy in Japan and the fifth worldwide. The mainstay of its treatment is hepatic resection with improved outcomes; however, HCC is still characterized by frequent recurrence [
1,
2]. microRNAs (miRNAs) are small (17–21 nt), non-coding RNAs that regulate gene expression at the post-transcriptional level through the RNA interference pathway. Currently, ~2000 miRNAs have been described in humans, and a single miRNA may regulate many mRNAs. Through this mechanism, miRNAs are essential components in the regulation of many cellular and developmental processes, including developmental timing, organ development, differentiation, proliferation, immune regulation, and cancer development and progression [
3]. Depending upon their target gene(s) and level of expression, miRNAs may function as either oncogenes or tumor suppressors and assist in the promotion or suppression of cancer growth and progression [
4,
5].
We previously demonstrated that circulating miRNAs in serum extracellular vesicles (exosomes) could be novel biomarkers for predicting the recurrence and therapeutic targets of HCC [
6,
7]. Exosomes are small membrane vesicles (30–100 nm) derived from the luminal membranes of multivesicular bodies and are constitutively released by fusion with the cell membrane [
8]. Exosomes transfer not only membrane components but also nucleic acids to other cells; therefore, cell-derived exosomes have recently been described as a new mode of cell-to-cell communication [
9]. To date, 764 microRNAs (miRs) have been identified in exosomes derived from several different cell types and from multiple organisms [
10]. In this light, exosomally transported miRNAs have found a place in cancer research as carriers of genetic information [
11,
12]. Functional exosomal miRs both from cancer cells and bone marrow (BM) mesenchymal stem cells have been reported [
13,
14], but in a previous study we could not show direct evidence that exosomal serum miRNA was secreted from either HCC cells or other host cells, such as BM-derived cells [
6]. Here we briefly report the miRNA profile of BM cells to understand the role of miRNAs in the progression of HCC.
2. Materials and Methods
Seven patients who consecutively underwent hepatectomy for primary HCC were selected from records of the Department of Gastroenterological Surgery, Osaka University. The institutional review board approved this study and we obtained written informed consent from each patient. The median age of the seven patients was 60 years (range 37–72); the etiologies were hepatitis C infection in four cases, hepatitis B in two, and alcoholism in one. The number of tumors was single in five cases and multiple in two cases. After the median follow-up time of 22 months, three cases had recurrence of HCC while four cases had no recurrence. On the basis of the UICC classification (7th edition), three cases were classified as stage I, two cases as stage II, and two cases as stage IIIA.
Aspiration of BM was conducted under general anesthesia immediately before surgery, as previously described [
15]. The BM aspirate was obtained from the sternum using a BM aspiration needle. A volume of 3 mL of BM was added to 4.0 mL of Isogen-LS (Nippon Gene, Toyama, Japan), which was shaken vigorously and stored at −80 °C until RNA extraction. BM cells were separated into three fractions using a three-step automagnetic-activated cell separation system (MACS) by MACS Cell Separators (Miltenyi Biotec, Bergisch Gladbach, Germany). CD45
+, CD14
+, and CD45
−/EpCAM
+ cell fractions were collected using CD45, CD14, and EpCAM (CD326) microbeads according to the manufacturer’s instructions (Miltenyi Biotec).
RNA was extracted from each BM fraction separated by the Auto MACS system using the miRNeasy Mini Kit (Qiagen, Venlo, The Netherlands) according to the manufacturer’s protocol. Extracted total RNA was labeled with Cy3 using the miRCURY LNA Array miR labeling kit (Exiqon, Vedbaek, Denmark). Labeled RNAs were hybridized onto 3D-Gene Human microRNA Oligo chips containing 837 anti-sense probes printed in duplicate spots (Toray, Kamakura, Japan). The annotation and oligonucleotide sequences of the probes conformed to the miRBase microRNA database (Faculty of Life Sciences, University of Manchester, Mancehster, UK). After stringent washes, fluorescent signals were scanned with the 3D-Gene Scanner (Toray) and analyzed using GenePix Pro version 5.0 (Molecular Devices, Sunnyvale, CA, USA). The raw data from each spot were normalized by subtraction of the background signal mean intensity, determined by the 95% confidence intervals of the signal intensities of all blank spots. Valid measurements were considered those in which the signal intensity of both duplicate spots was greater than two standard deviations of the background signal intensity. MicroRNAs differentially expressed between groups were statistically identified using the Welch t-test. Data was uploaded in Gene Expression Omnibus datasets (GSE71762, National Center for Biotechnology Information, Bethesda, MD, USA).
3. Results and Discussion
3.1. Whole BM Fraction
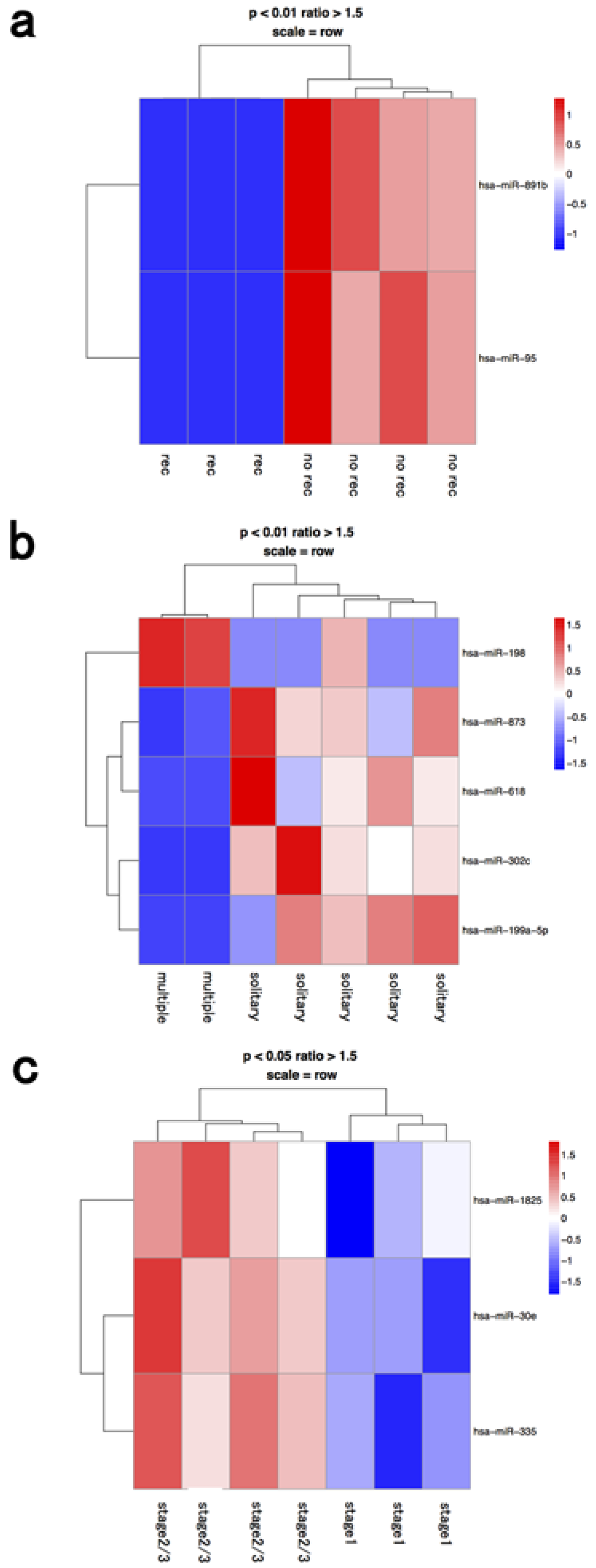
miRNA profiles in the whole BM fraction are shown in
Figure 1, depending on the recurrence factor, number of tumors, and clinical stage. Differentially expressed miRNAs were significantly different between fractions from whole bone marrow, macrophages, and lymphocytes (
Table 1). Therefore, we further analyzed the miRNA profiles of macrophage and lymphocyte fractions. Our present data showed that miRNAs of selected fractions had to be independently analyzed with respect to the origin of the microRNAs present in the BM of HCC patients. miRNA processing may occur in the cancer cells themselves or in cells within the BM microenvironment, such as hematopoietic progenitor cells, endothelial cells, progenitor cells, and macrophages [
16,
17,
18].
Figure 1.
Hierarchical clustering analysis of microRNAs of whole bone marrow cells in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in bone marrow cells from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed two, five, and three miRNAs that were significantly differentially expressed between the two groups with a >1.50-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Figure 1.
Hierarchical clustering analysis of microRNAs of whole bone marrow cells in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in bone marrow cells from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed two, five, and three miRNAs that were significantly differentially expressed between the two groups with a >1.50-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Table 1.
MicroRNAs in the bone marrow cells that were significantly correlated with clinical significance of HCC patients.
Table 1.
MicroRNAs in the bone marrow cells that were significantly correlated with clinical significance of HCC patients.
| BM Fraction | Clinical Significance | Upregulated microRNA | Downregulated microRNA |
|---|
| whole BM | recurrence | | hsa-mir-891b |
| | recurrence | | hsa-mir-95 |
| | multiple | hsa-mir-198 | hsa-mir-873 |
| | multiple | | hsa-mir-618 |
| | multiple | | hsa-mir-302c |
| | multiple | | hsa-mir-199a-5p |
| | stage 2≥ | hsa-mir-1825 | |
| | stage 2≥ | hsa-mir-30e | |
| | stage 2≥ | hsa-mir-335 | |
| lymphocyte | recurrence | hsa-mir-148a | hsa-mir-190 |
| | recurrence | hsa-mir-361-5p | hsa-mir-503 |
| | recurrence | hsa-mir-320d | hsa-mir-544 |
| | multiple | hsa-mir-654-5p | hsa-mir-517a |
| | multiple | | hsa-mir-497 |
| | multiple | | hsa-mir-454 |
| | multiple | | hsa-mir-22 |
| | stage 2≥ | hsa-mir-1537 | hsa-mir-345 |
| | stage 2≥ | hsa-mir-513b | hsa-mir-553 |
| | stage 2≥ | hsa-mir-15a | hsa-mir-653 |
| | stage 2≥ | hsa-mir-517a | hsa-mir-577 |
| | stage 2≥ | hsa-mir-28-5p | |
| macrophage | recurrence | hsa-mir-1207-3p | hsa-mir-1277 |
| | recurrence | hsa-mir-937 | hsa-mir-1279 |
| | recurrence | | hsa-mir-184 |
| | recurrence | | hsa-mir-563 |
| | recurrence | | hsa-mir-96 |
| | recurrence | | hsa-mir-302b |
| | multiple | hsa-mir-1 | hsa-mir-10b |
| | multiple | hsa-mir-889 | hsa-mir-204 |
| | multiple | hsa-mir-658 | hsa-mir-654-3p |
| | multiple | | hsa-mir-302a |
| | stage 2≥ | hsa-mir-555 | hsa-mir-942 |
| | stage 2≥ | hsa-mir-1293 | hsa-mir-1227 |
| | stage 2≥ | | hsa-mir-598 |
| | stage 2≥ | | hsa-mir-518d-3p |
3.2. Lymphocyte Fraction
Inflammation appears to be a crucial factor in hepatocarcinogenesis since HCC typically occurs in patients with chronic inflammatory liver diseases, such as viral hepatitis or non-alcoholic steatohepatitis [
19,
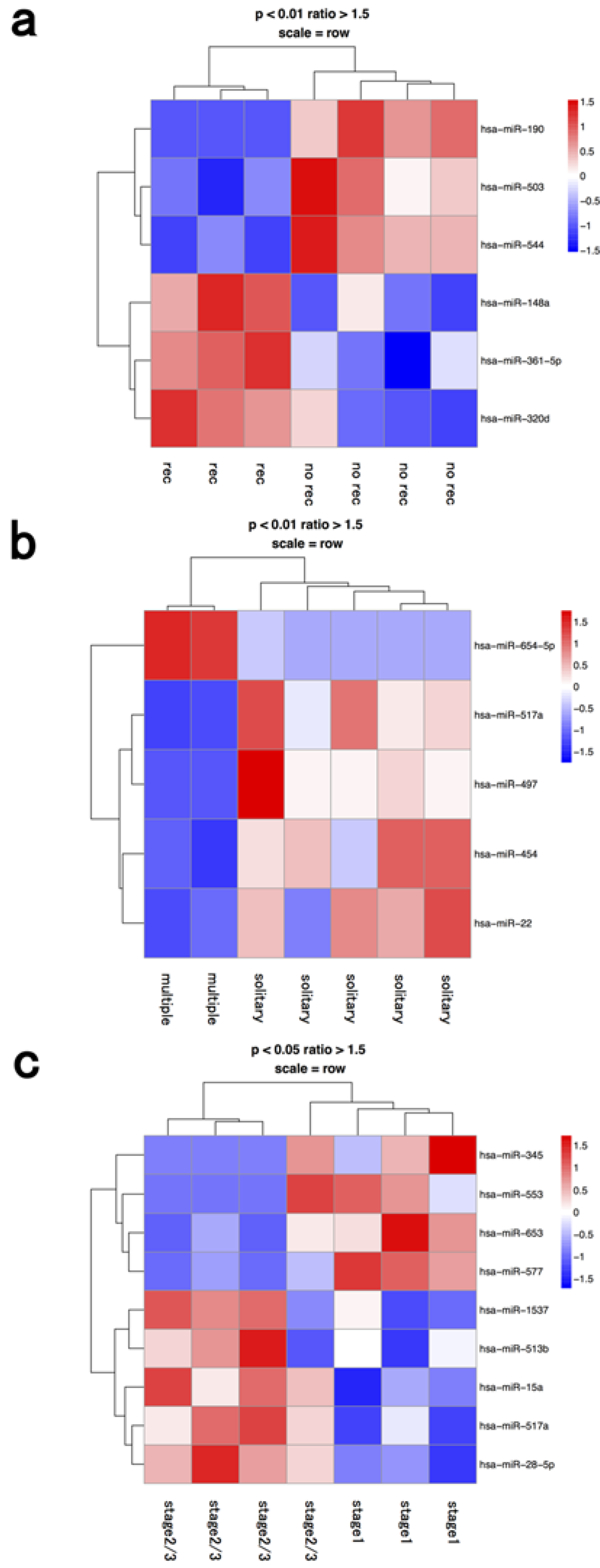
20]. One miR (hsa-miR-654-5p) was upregulated and four miRs (hsa-miR-517a, 497, 454, 22) were downregulated in the lymphocyte fraction of cases with multiple tumors compared to ones with a solitary tumor (
Figure 2 and
Table 1). miR-517a is located in the chromosome 19 miRNA cluster, and is considered to be a tumor-suppressive miRNA that is suppressed by epigenetic modifications [
21]. miR-517a inhibited cell proliferation by blocking G2/M transition in HCC [
21], and markedly induced bladder cancer cell apoptosis [
22]. One of the target genes of miR-517a in HCC was reported to be Pyk2, which was associated with MAP kinase signaling pathways [
23]. miR-497, clustered at 17p13.1, is also reported to be a tumor suppressor and shows significant growth-suppressive activity with induction of G1 arrest in HCC [
24]. Potential target genes of miR-497 in cancers had been reported to be insulin-like growth factor 1 receptor, WEE1, HDGF, VEGF-A, Akt, and IKKβ [
25,
26,
27,
28].
3.3. Macrophage Fraction
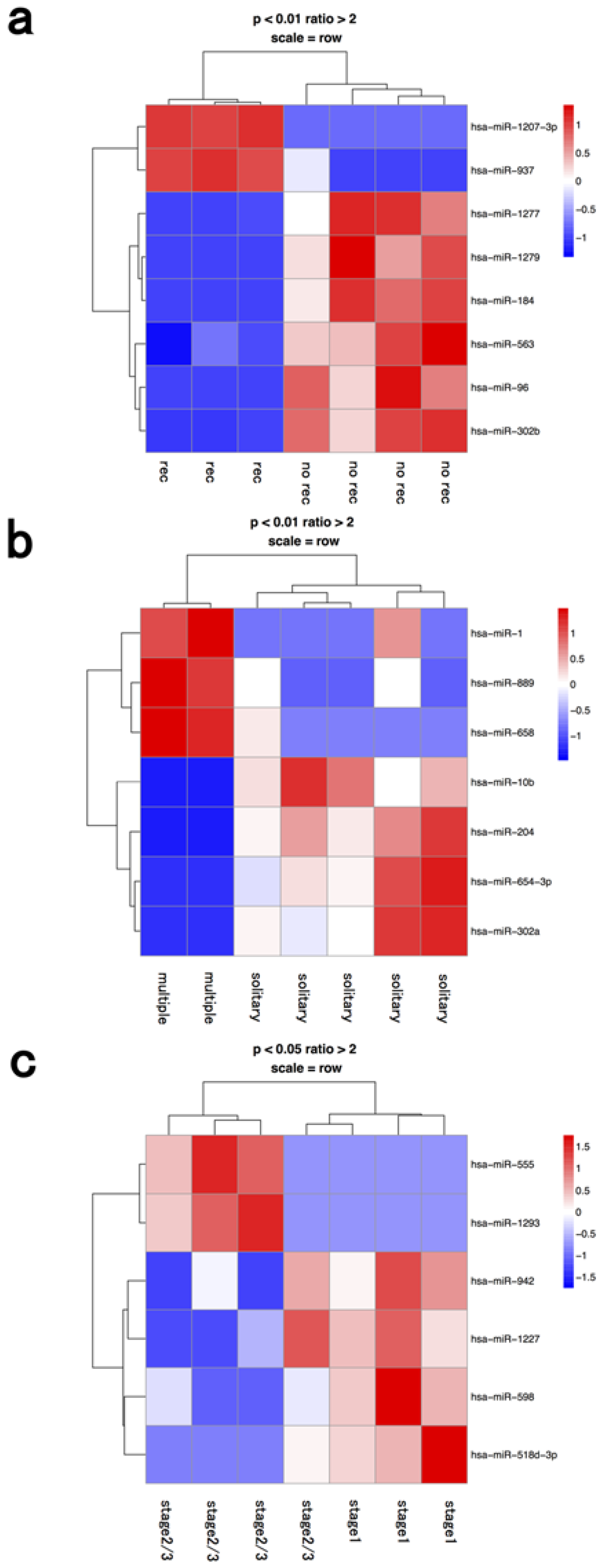
Two miRNAs were upregulated (hsa-miR-1207-3p, 937) and six miRNAs (hsa-miR-1277, 1279, 184, 563, 96, 302b) were downregulated in the macrophage fractions from cases with post-operative recurrence compared to fractions from cases with no recurrence (
Figure 3 and
Table 1).
Previous studies have shown that miR-184 can act either as an oncogenic- or a tumor suppressive-miRNA in various human cancers, depending on cellular context [
29,
30]. Lin
et al. reported that decreased miR-184 promotes cancer cell invasiveness by an increase in CDC25A and c-myc expression [
31]. In HCC, miR-302b acts as a tumor suppressor by targeting AKT2, suppressing G1 regulators (Cyclin A, Cyclin D1, CDK2) and increasing p27Kip1 phosphorylation at Ser10 [
32]. Recent studies have revealed that tumor-associated macrophages (TAMs) are an important component of the tumor microenvironment and can promote tumor progression [
35,
36]. TAMs were reported to be associated with metastasis, angiogenesis, epithelial-mesenchymal transition (EMT), and poor prognosis in HCC [
35,
37,
38]. Macrophages have multiple biological roles, including antigen presentation, target cell cytotoxicity, removal of foreign bodies, tissue remodeling, regulation of inflammation, induction of immunity, thrombosis, and endocytosis. Aucher
et al. recently reported that transfer of miRNAs from macrophages functionally inhibited proliferation of HCC cells [
39]. Although the mechanisms by which TAMs promote tumor progression are poorly understood, our data implied that they might act through altered miRNA expression.
4. Conclusions
These results suggest that miRNA profiles in separated fractions of bone marrow cells are associated with metastasis, angiogenesis, epithelial-mesenchymal transition (EMT), and poor prognosis in HCC. In this study, the number of cases was small, therefore our data are preliminary and further analysis including validation studies with large cohorts and in vitro studies to search target genes and pathways of identified miRNAs are warranted. We revealed the miR profiling of BM cells associated with stage/recurrence of HCC in this study. It is possible that miR profiling of BM niche was affected by the progression of HCC or disseminated cancer cells. On the other hand, altered miR expression of BM cells might help the survival or proliferation of cancer cells in BM. At present, it is difficult to conclude that one specific mechanism is superior to others, and further studies are recommended. Our data could provide a database to seek new concepts in immunotherapy targeting miRNAs of BM cells to improve the outcome of patients with HCC in the near future.
Figure 2.
Hierarchical clustering analysis of microRNAs of lymphocyte fraction in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in macrophages from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed six, five, and nine miRNAs that were significantly differentially expressed between the two groups with a >1.50-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Figure 2.
Hierarchical clustering analysis of microRNAs of lymphocyte fraction in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in macrophages from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed six, five, and nine miRNAs that were significantly differentially expressed between the two groups with a >1.50-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Figure 3.
Hierarchical clustering analysis of microRNAs of macrophage fraction in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in lymphocytes from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed eight, seven, and six miRNAs that were significantly differentially expressed between the two groups with a >2-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Figure 3.
Hierarchical clustering analysis of microRNAs of macrophage fraction in seven patients with hepatocellular carcinoma. Heat map of the miRNA profile in lymphocytes from hepatocellular carcinoma patients with (a) recurrence (n = 3) and no recurrence (n = 4); (b) solitary (n = 5) and multiple tumors (n = 2); (c) stage 1 (n = 3) and ≥stage 2 (n = 4). Cluster analysis showed eight, seven, and six miRNAs that were significantly differentially expressed between the two groups with a >2-fold change, respectively. Colors range from blue to red, corresponding to low to high expression, respectively. p values <0.01 or <0.05, unpaired t test. rec: recurrence.
Acknowledgments
We thank Kazumi Oda, Michiko Kasagi, Satoko Kohno, and Tomoko Kawano for their technical assistance. This work was supported in part by the following grants and foundations: Japan Society for the Promotion of Science (JSPS) Grant-in-Aid for Scientific Research, grant number 24592005, and Grant-in Aid for Young Scientists S, grant number 21679006.
Author Contributions
Keishi Sugimachi, Shotaro Sakimura and Koshi Mimori contribute to conception and design of the study, the acquisition of data and writing the manuscript. Akira Tomokuni and Hidetoshi Eguchi (Osaka University) contribute to the collection of samples and the acquisition of data. Ryutaro Uchi, Hidenari Hirata, Hisateru Komatsu, Yoshiaki Shinden, Tomohiro Iguchi, Hidetoshi Eguchi (Kyushu University), Takaaki Masuda and Kazutoyo Morita contribute to the analysis of data. Ken Shirabe, Yoshihiko Maehara and Masaki Mori contribute to the interpretation of data.
Conflicts of Interest
Keishi Sugimachi and co-authors have no conflicts of interest.
References
- Okita, K. Clinical aspects of hepatocellular carcinoma in japan. Intern. Med. 2006, 45, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M. Updated treatment approach to hepatocellular carcinoma. J. Gastroenterol. 2005, 40, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Taketomi, A.; Shirabe, K.; Umeda, K.; Kayashima, H.; Ninomiya, M.; Uchiyama, H.; Soejima, Y.; Maehara, Y. Clinical significance and potential of hepatic microRNA-122 expression in hepatitis c. Liver Int. 2011, 31, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Revi. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Sugimachi, K.; Matsumura, T.; Hirata, H.; Uchi, R.; Ueda, M.; Ueo, H.; Shinden, Y.; Iguchi, T.; Eguchi, H.; Shirabe, K.; et al. Identification of a bona fide microRNA biomarker in serum exosomes that predicts hepatocellular carcinoma recurrence after liver transplantation. Br. J. Cancer 2015, 112, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, T.; Sugimachi, K.; Iinuma, H.; Takahashi, Y.; Kurashige, J.; Sawada, G.; Ueda, M.; Uchi, R.; Ueo, H.; Takano, Y.; et al. Exosomal microRNA in serum is a novel biomarker of recurrence in human colorectal cancer. Br. J. Cancer 2015, 113, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; Garcia-Santos, G.; Ghajar, C.; et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through met. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathivanan, S.; Fahner, C.J.; Reid, G.E.; Simpson, R.J. Exocarta 2012: Database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 2012, 40, D1241–D1244. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Ding, W.Q. Intercellular communication by exosome-derived microRNAs in cancer. Int. J. Mol. Sci. 2013, 14, 14240–14269. [Google Scholar] [CrossRef] [PubMed]
- Cereghetti, D.M.; Lee, P.P. Tumor-derived exosomes contain microRNAs with immunological function: Implications for a novel immunosuppression mechanism. MicroRNA (Shariqah, United Arab Emirates) 2014, 2, 194–204. [Google Scholar] [CrossRef]
- Ono, M.; Kosaka, N.; Tominaga, N.; Yoshioka, Y.; Takeshita, F.; Takahashi, R.U.; Yoshida, M.; Tsuda, H.; Tamura, K.; Ochiya, T. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci. Signal. 2014, 7, ra63. [Google Scholar] [CrossRef] [PubMed]
- Mimori, K.; Fukagawa, T.; Kosaka, Y.; Ishikawa, K.; Iwatsuki, M.; Yokobori, T.; Hirasaki, S.; Takatsuno, Y.; Sakashita, H.; Ishii, H.; et al. A large-scale study of mt1-mmp as a marker for isolated tumor cells in peripheral blood and bone marrow in gastric cancer cases. Ann. Surg. Oncol. 2008, 15, 2934–2942. [Google Scholar] [CrossRef] [PubMed]
- Ota, D.; Mimori, K.; Yokobori, T.; Iwatsuki, M.; Kataoka, A.; Masuda, N.; Ishii, H.; Ohno, S.; Mori, M. Identification of recurrence-related microRNAs in the bone marrow of breast cancer patients. Int. J. Oncol. 2011, 38, 955–962. [Google Scholar] [PubMed]
- Akiyoshi, S.; Fukagawa, T.; Ueo, H.; Ishibashi, M.; Takahashi, Y.; Fabbri, M.; Sasako, M.; Maehara, Y.; Mimori, K.; Mori, M. Clinical significance of mir-144-zfx axis in disseminated tumour cells in bone marrow in gastric cancer cases. Br. J. Cancer 2012, 107, 1345–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeyama, H.; Yamamoto, H.; Yamashita, S.; Wu, X.; Takahashi, H.; Nishimura, J.; Haraguchi, N.; Miyake, Y.; Suzuki, R.; Murata, K.; et al. Decreased mir-340 expression in bone marrow is associated with liver metastasis of colorectal cancer. Mol. Cancer Ther. 2014, 13, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Shirabe, K.; Mano, Y.; Muto, J.; Matono, R.; Motomura, T.; Toshima, T.; Takeishi, K.; Uchiyama, H.; Yoshizumi, T.; Taketomi, A.; et al. Role of tumor-associated macrophages in the progression of hepatocellular carcinoma. Surg. Today 2012, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mossanen, J.C.; Tacke, F. Role of lymphocytes in liver cancer. Oncoimmunology 2013, 2, e26468. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.F.; Xu, X.; Huang, J.; Fei, Q.L.; Chen, F.; Li, Y.D.; Han, Z.G. Down-regulation of mir-517a and mir-517c promotes proliferation of hepatocellular carcinoma cells via targeting pyk2. Cancer Lett. 2013, 329, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, T.; Kawakami, K.; Enokida, H.; Chiyomaru, T.; Kagara, I.; Tatarano, S.; Yoshino, H.; Arimura, H.; Nishiyama, K.; Seki, N.; et al. Restoration of mir-517a expression induces cell apoptosis in bladder cancer cell lines. Oncol. Rep. 2011, 25, 1661–1668. [Google Scholar] [PubMed]
- Sun, C.K.; Man, K.; Ng, K.T.; Ho, J.W.; Lim, Z.X.; Cheng, Q.; Lo, C.M.; Poon, R.T.; Fan, S.T. Proline-rich tyrosine kinase 2 (pyk2) promotes proliferation and invasiveness of hepatocellular carcinoma cells through c-src/erk activation. Carcinogenesis 2008, 29, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Furuta, M.; Kozaki, K.; Tanimoto, K.; Tanaka, S.; Arii, S.; Shimamura, T.; Niida, A.; Miyano, S.; Inazawa, J. The tumor-suppressive mir-497-195 cluster targets multiple cell-cycle regulators in hepatocellular carcinoma. PLoS ONE 2013, 8, e60155. [Google Scholar] [CrossRef] [PubMed]
- Creevey, L.; Ryan, J.; Harvey, H.; Bray, I.M.; Meehan, M.; Khan, A.R.; Stallings, R.L. MicroRNA-497 increases apoptosis in mycn amplified neuroblastoma cells by targeting the key cell cycle regulator wee1. Mol. Cancer 2013, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.T.; Jiang, C.C.; Wang, G.P.; Li, Y.P.; Wang, C.Y.; Guo, X.Y.; Yang, R.H.; Feng, Y.; Wang, F.H.; Tseng, H.Y.; et al. MicroRNA-497 targets insulin-like growth factor 1 receptor and has a tumour suppressive role in human colorectal cancer. Oncogene 2013, 32, 1910–1920. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.J.; Duan, L.J.; Qian, X.Q.; Xu, D.; Liu, H.L.; Zhu, Y.J.; Qi, J. Tumor-suppressive microRNA-497 targets ikkbeta to regulate NF-kappab signaling pathway in human prostate cancer cells. Am. J. Cancer Res. 2015, 5, 1795–1804. [Google Scholar] [PubMed]
- Shao, X.J.; Miao, M.H.; Xue, J.; Xue, J.; Ji, X.Q.; Zhu, H. The down-regulation of microRNA-497 contributes to cell growth and cisplatin resistance through pi3k/akt pathway in osteosarcoma. Cell Physiol. Biochem. 2015, 36, 2051–2062. [Google Scholar] [CrossRef] [PubMed]
- Phua, Y.W.; Nguyen, A.; Roden, D.L.; Elsworth, B.; Deng, N.; Nikolic, I.; Yang, J.; McFarland, A.; Russell, R.; Kaplan, W.; et al. MicroRNA profiling of the pubertal mouse mammary gland identifies mir-184 as a candidate breast tumour suppressor gene. Breast Cancer Res. 2015, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.S.; Liu, X.B.; Wong, B.Y.; Ng, R.W.; Yuen, A.P.; Wei, W.I. Mature mir-184 as potential oncogenic microRNA of squamous cell carcinoma of tongue. Clinical cancer research : an official journal of the American Association for Cancer Research 2008, 14, 2588–2592. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.C.; Lin, P.L.; Cheng, Y.W.; Wu, T.C.; Chou, M.C.; Chen, C.Y.; Lee, H. MicroRNA-184 deregulated by the microRNA-21 promotes tumor malignancy and poor outcomes in non-small cell lung cancer via targeting cdc25a and c-myc. Ann. Surg. Oncol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yao, J.; Zhang, X.; Guo, B.; Le, X.; Cubberly, M.; Li, Z.; Nan, K.; Song, T.; Huang, C. MiRNA-302b suppresses human hepatocellular carcinoma by targeting akt2. Mol. Cancer Res. 2014, 12, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, M.J.; Ren, A.M.; Wu, H.F.; Han, W.M.; Tan, R.Y.; Tu, R.Q. A ten-microRNA signature identified from a genome-wide microRNA expression profiling in human epithelial ovarian cancer. PLoS ONE 2014, 9, e96472. [Google Scholar] [CrossRef] [PubMed]
- Ujihira, T.; Ikeda, K.; Suzuki, T.; Yamaga, R.; Sato, W.; Horie-Inoue, K.; Shigekawa, T.; Osaki, A.; Saeki, T.; Okamoto, K.; et al. MicroRNA-574-3p, identified by microRNA library-based functional screening, modulates tamoxifen response in breast cancer. Sci. Rep. 2015, 5, 7641. [Google Scholar] [CrossRef] [PubMed]
- Shirabe, K.; Motomura, T.; Muto, J.; Toshima, T.; Matono, R.; Mano, Y.; Takeishi, K.; Ijichi, H.; Harada, N.; Uchiyama, H.; et al. Tumor-infiltrating lymphocytes and hepatocellular carcinoma: Pathology and clinical management. Int. J. Clin. Oncol. 2010, 15, 552–558. [Google Scholar] [CrossRef] [PubMed]
- De Visser, K.E.; Eichten, A.; Coussens, L.M. Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 2006, 6, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Rolny, C.; Capparuccia, L.; Casazza, A.; Mazzone, M.; Vallario, A.; Cignetti, A.; Medico, E.; Carmeliet, P.; Comoglio, P.M.; Tamagnone, L. The tumor suppressor semaphorin 3b triggers a prometastatic program mediated by interleukin 8 and the tumor microenvironment. J. Exp. Med. 2008, 205, 1155–1171. [Google Scholar] [CrossRef] [PubMed]
- Van Zijl, F.; Mair, M.; Csiszar, A.; Schneller, D.; Zulehner, G.; Huber, H.; Eferl, R.; Beug, H.; Dolznig, H.; Mikulits, W. Hepatic tumor-stroma crosstalk guides epithelial to mesenchymal transition at the tumor edge. Oncogene 2009, 28, 4022–4033. [Google Scholar] [CrossRef] [PubMed]
- Aucher, A.; Rudnicka, D.; Davis, D.M. MicroRNAs transfer from human macrophages to hepato-carcinoma cells and inhibit proliferation. J. Immunol. (Baltimore, Md.: 1950) 2013, 191, 6250–6260. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).




{kind=link}
{kind=link}
{kind=link}

