

Skeletal Class II Malocclusion: From Clinical Treatment Strategies to the Roadmap in Identifying the Genetic Bases of Development in Humans with the Support of the Collaborative Cross Mouse Population
 ,
, 
Abstract
:1. Introduction
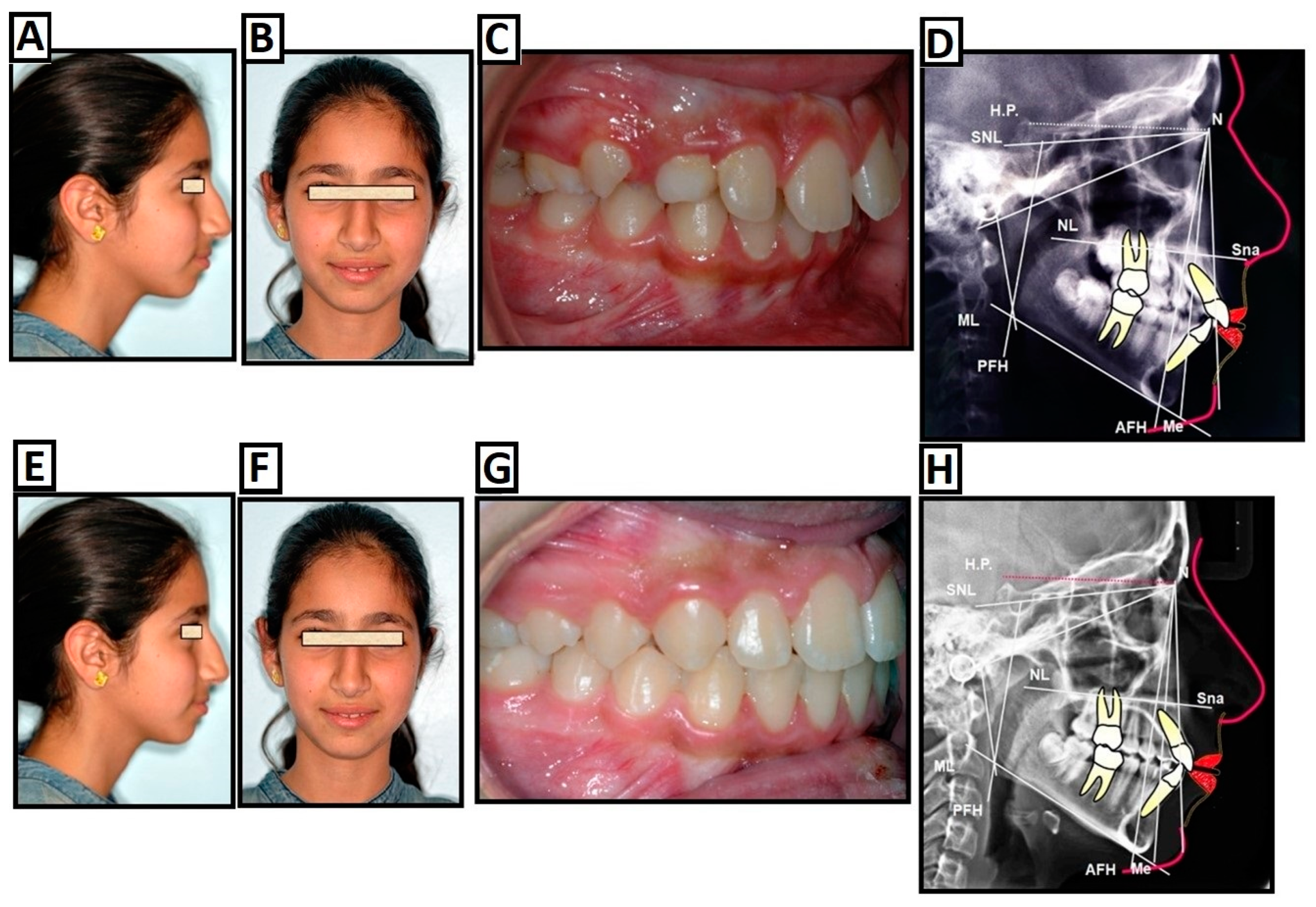
2. Clinical Treatment Strategies
2.1. Treatments Options for Skeletal Class II Malocclusion
- I.
- A causal therapeutic approach in the sense of a targeted influence on the growth component. For this purpose, functional orthodontic devices for ventral development of the mandible with simultaneous growth inhibition of the upper jaw in the sagittal and vertical directions (HG effect) are used in the therapy of class II dysgnathy.
- II.
- A dentoalveolar therapeutic approach in the sense of conservative space creation measures. The focus here is on the distalization of the 6-year molars up to the neutral position so that, after subsequent distalization of the support zone and retraction of the front of the upper jaw, a correction of the sagittal front tooth step is possible. Many factors, such as space conditions, must be considered for this treatment measure; the sagittal and vertical cranial structures, as well as dentofacial aesthetics, are considered.
- III.
- A dentoalveolar therapy approach in the sense of absolute space creation measures by extracting permanent teeth. In addition to the systematic extraction of the premolars, the extraction of two premolars should only be considered in the upper jaw if the initial situation is appropriate; the goal of treatment concerning the occlusion is therefore neutral on the canines and 1PB distal on the first malar. For these treatment measures, as with the previously described options, many factors must also be considered.
- IV.
- A skeletal correction in the sense of jaw surgery compensation. If the growth for the correction of a pronounced class II is no longer available and a conservative or an absolute space creation measure is not indicated, the treatment of the dysgnathia is only possible in this case with a combined orthodontic and maxillofacial surgical treatment.
2.2. Correction of Increased Interincisal Angle
2.3. Correction of Increased Overbite
3. Etiology of SCIIMO Development
3.1. Genetics and Heritability of Class II Malocclusion
3.2. Importance of Genetic Studies
4. A Mouse Model for Studying Class II Malocclusion
4.1. QTL Analysis in a Mouse Model
4.2. The Collaborative Cross Mouse Cohort Represents a Valuable Resource for Conducting a System Genetic Study on Class II Malocclusion
4.3. Future Direction of Developing a Novel Model for Mapping Significant and Modifier Genes Associated with Skeletal Class II Malocclusion Using the CC Model
- Crossing a mouse carrying mutant genes known to be associated with SCIIMO (e.g., EPB41, Nog, or any other genes listed in Table 1) with a set of mice possessing diverse genetic backgrounds of naturally occurring variations, it becomes possible to establish a framework for mapping and identifying modifier loci associated with the altered gene. As a result, we suggest executing a unique study by breeding SCIIMO mutant mice with various CC lines. This will produce F1 (SCIIMO+/− x CC) cells, which may have dramatically diverse susceptibilities to SCIIMO development (Figure 10). Our team successfully used a similar technique to map modifiers for the APC mutant gene, which is linked to the development of colorectal cancer [85].
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| QTL | Quantitative Trait Loci |
| RCTs | Research Clinical Trials |
| GWAS | Genome-Wide Associations |
| EWAS | Epigenetics-Wide Associations |
| FFA | Fixed Functional Appliances |
| SNPs | Single-Nucleotide Polymorphisms |
| NOG gene | noggin gene |
| RI | Recombinant Inbreed |
| GRP | Genetic Reference Population |
| CI | Confidence Interval |
References
- Guo, L.; Feng, Y.; Guo, H.G.; Liu, B.W.; Zhang, Y. Consequences of Orthodontic Treatment in Malocclusion Patients: Clinical and Microbial Effects in Adults and Children. BMC Oral Health 2016, 16, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhammadi, M.S.; Halboub, E.; Fayed, M.S.; Labib, A.; El-Saaidi, C. Global Distribution of Malocclusion Traits: A Systematic Review. Dent. Press J. Orthod. 2018, 23, 40.e1. [Google Scholar] [CrossRef] [PubMed]
- Proffit, W.R.; Fields, H.W., Jr.; Moray, L.J. Prevalence of Malocclusion and Orthodontic Treatment Need in the United States: Estimates from the NHANES III Survey. Int. J. Adult Orthod. Orthognath. Surg. 1998, 13, 97–106. [Google Scholar]
- Khateeb, E.A.A.A.; Khateeb, S.N.A. Anteroposterior and Vertical Components of Class II Division 1 and Division 2 Malocclusion. Angle Orthod. 2009, 79, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, A.A.; Yamashita, H.K.; Lederman, H.M.; Cevidanes, L.H.S.; Proffit, W.R.; Vigorito, J.W. Fränkel Appliance Therapy and the Temporomandibular Disc: A Prospective Magnetic Resonance Imaging Study. Am. J. Orthod. Dentofac. Orthop. 2002, 121, 447–457. [Google Scholar] [CrossRef]
- Stahl, F.; Baccetti, T.; Franchi, L.; McNamara, J.A. Longitudinal Growth Changes in Untreated Subjects with Class II Division 1 Malocclusion. Am. J. Orthod. Dentofac. Orthop. 2008, 134, 125–137. [Google Scholar] [CrossRef]
- Al-Dumaini, A.A.; Halboub, E.; Alhammadi, M.S.; Ishaq, R.A.R.; Youssef, M. A Novel Approach for Treatment of Skeletal Class II Malocclusion: Miniplates-Based Skeletal Anchorage. Am. J. Orthod. Dentofac. Orthop. 2018, 153, 239–247. [Google Scholar] [CrossRef]
- Cozza, P.; Marino, A.; Franchi, L. A Nonsurgical Approach to Treatment of High-Angle Class II Malocclusion. Angle Orthod. 2008, 78, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Ruf, S.; Dent, M.; Baltromejus, S.; Pancherz, H.; Dr, O. Effective Condylar Growth and Chin Position Changes in Activator Treatment: A Cephalometric Roentgenographic Study. Angle Orthod. 2001, 71, 4–11. [Google Scholar]
- Serbesis-Tsarudis, C.; Pancherz, H. “Effective” TMJ and Chin Position Changes in Class II Treatment. Angle Orthod. 2008, 78, 813–818. [Google Scholar] [CrossRef]
- Elfeky, H.Y.; Fayed, M.S.; Alhammadi, M.S.; Soliman, S.A.Z.; el Boghdadi, D.M. Three-Dimensional Skeletal, Dentoalveolar and Temporomandibular Joint Changes Produced by Twin Block Functional Appliance. J. Orofac. Orthop. 2018, 79, 245–258. [Google Scholar] [CrossRef]
- Marsico, E.; Gatto, E.; Burrascano, M.; Matarese, G.; Cordasco, G. Effectiveness of Orthodontic Treatment with Functional Appliances on Mandibular Growth in the Short Term. Am. J. Orthod. Dentofac. Orthop. 2011, 139, 24–36. [Google Scholar] [CrossRef]
- Rodrigues De Almeida, M.; Fernando, J.; Henriques, C.; Rodrigues De Almeida, R.; Ursi, W. Treatment Effects Produced by Fränkel Appliance in Patients with Class II, Division 1 Malocclusion*. Angle Orthod. 2002, 72, 418. [Google Scholar] [PubMed]
- Toth, L.R.; McNamara, J.A. Treatment Effects Produced by the Twin-Block Appliance and the FR-2 Appliance of Fränkel Compared with an Untreated Class II Sample. Am. J. Orthod. Dentofac. Orthop. 1999, 116, 597–609. [Google Scholar] [CrossRef]
- Flores-Mir, C.; Major, P.W. A Systematic Review of Cephalometric Facial Soft Tissue Changes with the Activator and Bionator Appliances in Class II Division 1 Subjects. Eur. J. Orthod. 2006, 28, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Flores-Mir, C.; Major, P.W. Cephalometric Facial Soft Tissue Changes with the Twin Block Appliance in Class II Division 1 Malocclusion Patients: A Systematic Review. Angle Orthod. 2006, 76, 876–881. [Google Scholar]
- Flores-Mir, C.; Major, M.P.; Major, P.W. Soft Tissue Changes with Fixed Functional Appliances in Class II Division 1: A Systematic Review. Angle Orthod. 2006, 76, 712–720. [Google Scholar]
- Alkan, Ö.; Kaya, Y.; Tunca, M.; Keskin, S. Changes in the Gingival Thickness and Keratinized Gingival Width of Maxillary and Mandibular Anterior Teeth after Orthodontic Treatment. Angle Orthod. 2021, 91, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Barnett, G.A.; Higgins, D.W.; Major, P.W.; Flores-Mir, C. Immediate Skeletal and Dentoalveolar Effects of the Crown- or Banded Type Herbst Appliance on Class II Division 1 Malocclusion. Angle Orthod. 2008, 78, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, S.; Nebbe, B.; Normando, D.; Lagravere, M.O.; Flores-Mir, C. Short-Term Treatment Effects Produced by the Twin-Block Appliance: A Systematic Review and Meta-Analysis. Eur. J. Orthod. 2014, 37, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Antonarakis, G.S.; Kiliaridis, S. Short-Term Anteroposterior Treatment Effects of Functional Appliances and Extraoral Traction on Class II Malocclusion: A Meta-Analysis. Angle Orthod. 2007, 77, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Koretsi, V.; Zymperdikas, V.F.; Papageorgiou, S.N.; Papadopoulos, M.A. Treatment Effects of Removable Functional Appliances in Patients with Class II Malocclusion: A Systematic Review and Meta-Analysis. Eur. J. Orthod. 2015, 37, 418–434. [Google Scholar] [CrossRef] [PubMed]
- Cacciatore, G.; Ghislanzoni, L.T.H.; Alvetro, L.; Giuntini, V.; Franchi, L. Treatment and Posttreatment Effects Induced by the Forsus Appliance: A Controlled Clinical Study. Angle Orthod. 2014, 84, 1010–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küçükkeleş, N.; Ilhan, I.; Orgun, I.A. Treatment Efficiency in Skeletal Class II Patients Treated with the Jasper Jumper: A Cephalometric Evaluation. Angle Orthod. 2007, 77, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oztoprak, M.O.; Nalbantgil, D.; Uyanlar, A.; Arun, T. A Cephalometric Comparative Study of Class II Correction with Sabbagh Universal Spring (SUS2) and Forsus FRD Appliances. Eur. J. Dent. 2012, 6, 302. [Google Scholar] [CrossRef]
- Bidjan, D.; Sallmann, R.; Eliades, T.; Papageorgiou, S.N. Orthopedic Treatment for Class Ii Malocclusion with Functional Appliances and Its Effect on Upper Airways: A Systematic Review with Meta-Analysis. J. Clin. Med. 2020, 9, 3806. [Google Scholar] [CrossRef]
- Perinetti, G.; Primožič, J.; Furlani, G.; Franchi, L.; Contardo, L. Treatment Effects of Fixed Functional Appliances Alone or in Combination with Multibracket Appliances: A Systematic Review and Meta-Analysis. Angle Orthod. 2015, 85, 480–492. [Google Scholar] [CrossRef]
- Zymperdikas, V.F.; Koretsi, V.; Papageorgiou, S.N.; Papadopoulos, M.A. Treatment Effects of Fixed Functional Appliances in Patients with Class II Malocclusion: A Systematic Review and Meta-Analysis. Eur. J. Orthod. 2016, 38, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Aslan, B.I.; Kucukkaraca, E.; Turkoz, C.; Dincer, M. Treatment Effects of the Forsus Fatigue Resistant Device Used with Miniscrew Anchorage. Angle Orthod. 2014, 84, 76–87. [Google Scholar] [CrossRef]
- Eissa, O.; El-Shennawy, M.; Gaballah, S.; El-Meehy, G.; el Bialy, T. Treatment Outcomes of Class II Malocclusion Cases Treated with Miniscrew-Anchored Forsus Fatigue Resistant Device: A Randomized Controlled Trial. Angle Orthod. 2017, 87, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Turkkahraman, H.; Eliacik, S.K.; Findik, Y. Effects of Miniplate Anchored and Conventional Forsus Fatigue Resistant Devices in the Treatment of Class II Malocclusion. Angle Orthod. 2016, 86, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkordy, S.A.; Abouelezz, A.M.; Fayed, M.M.S.; Attia, K.H.; Ishaq, R.A.R.; Mostafa, Y.A. Three-Dimensional Effects of the Mini-Implant-Anchored Forsus Fatigue Resistant Device: A Randomized Controlled Trial. Angle Orthod 2016, 86, 292–305. [Google Scholar] [CrossRef] [Green Version]
- Ozbilek, S.; Gungor, A.Y.; Celik, S. Effects of Skeletally Anchored Class II Elastics: A Pilot Study and New Approach for Treating Class II Malocclusion. Angle Orthod. 2017, 87, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Manni, A.; Migliorati, M.; Calzolari, C.; Silvestrini-Biavati, A. Herbst Appliance Anchored to Miniscrews in the Upper and Lower Arches vs Standard Herbst: A Pilot Study. Am. J. Orthod. Dentofac. Orthop. 2019, 156, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Kochar, G.D.; Londhe, S.; Shivpuri, A.; Chopra, S.; Mitra, R.; Verma, M. Management of Skeletal Class II Malocclusion Using Bimaxillary Skeletal Anchorage Supported Fixed Functional Appliances: A Novel Technique. J. Orofac. Orthop. 2021, 82, 42–53. [Google Scholar] [CrossRef]
- Elkordy, S.A.; Aboelnaga, A.A.; Fayed, M.M.S.; Aboulfotouh, M.H.; Abouelezz, A.M. Can the Use of Skeletal Anchors in Conjunction with Fixed Functional Appliances Promote Skeletal Changes? A Systematic Review and Meta-Analysis. Eur. J. Orthod. 2016, 38, 532–545. [Google Scholar] [CrossRef]
- Arvind, P.; Jain, R.K. Skeletally Anchored Forsus Fatigue Resistant Device for Correction of Class II Malocclusions—A Systematic Review and Meta-Analysis. Orthod. Craniofacial Res. 2021, 24, 52–61. [Google Scholar] [CrossRef]
- Bakdach, W.M.M.; Hadad, R. Is There Any Enhanced Treatment Effect on Class II Growing Patients When Forsus Fatigue Resistant Device Is Reinforced by Either Miniplates or Miniscrews? A Systematic Review and Meta-Analysis. Int. Orthod. 2021, 19, 15–24. [Google Scholar] [CrossRef]
- Gennaro, P.; Chisci, G.; Aboh, I.V.; Gabriele, G.; Cascino, F.; Iannetti, G. Comparative Study in Orthognathic Surgery between Dolphin Imaging Software and Manual Prediction. J. Craniofacial Surg. 2014, 25, 1577–1578. [Google Scholar] [CrossRef]
- McDowell, E.H.; Baker, I.M. The Skeletodental Adaptations in Deep Bite Correction. Am. J. Orthod. Dentofac. Orthop. 1991, 100, 370–375. [Google Scholar] [CrossRef]
- Wylie, W.L. The Relationship between Ramus Height, Dental Height, and Overbite. Am. J. Orthod. Oral Surg. 1946, 32, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Baccetti, T. Prediction of Individual Mandibular Changes Induced by Functional Jaw Orthopedics Followed by Fixed Appliances in Class II Patients. Angle Orthod. 2006, 76, 950–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seok, H.; Kim, S.G. Correction of Malocclusion by Botulinum Neurotoxin Injection into Masticatory Muscles. Toxins 2018, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Mossey, P.A. The Heritability of Malocclusion: Part 1--Genetics, Principles and Terminology. Br. J. Orthod. 1999, 26, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Mossey, P.A. The Heritability of Malocclusion: Part 2. The Influence of Genetics in Malocclusion. Br. J. Orthod. 1999, 26, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Basdra, E.K.; Kiokpasoglou, M.; Stellzig, A. The Class II Division 2 Craniofacial Type Is Associated with Numerous Congenital Tooth Anomalies. Eur. J. Orthod. 2000, 22, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Nimri, K.; Gharaibeh, T. Space Conditions and Dental and Occlusal Features in Patients with Palatally Impacted Maxillary Canines: An Aetiological Study. Eur. J. Orthod. 2005, 27, 461–465. [Google Scholar] [CrossRef]
- Watnick, S.S. Inheritance of Craniofacial Morphology. Angle Orthod. 1972, 42, 339–351. [Google Scholar]
- Nakata, M.; Yu, P.I.; Davis, B.; Nance, W.E. The Use of Genetic Data in the Prediction of Craniofacial Dimensions. Am. J. Orthod. 1973, 63, 471–480. [Google Scholar] [CrossRef]
- Ruf, S.; Pancherz, H. Class II Division 2 Malocclusion: Genetics or Environment? A Case Report of Monozygotic Twins. Angle Orthod. 1999, 69, 321–324. [Google Scholar]
- Liu, H.; Deng, H.; Cao, C.F.; Ono, H. Genetic Analysis of Dental Traits in 82 Pairs of Female-Female Twins. Chin. J. Dent. Res. 1998, 1, 12–16. [Google Scholar] [PubMed]
- Peck, S.; Peck, L.; Kataja, M. Class II Division 2 Malocclusion: A Heritable Pattern of Small Teeth in Well-Developed Jaws. Angle Orthod. 1998, 68, 9–20. [Google Scholar]
- Nakasima, A.; Ichinose, M.; Nakata, S.; Takahama, Y. Hereditary Factors in the Craniofacial Morphology of Angle’s Class II and Class III Malocclusions. Am. J. Orthod. 1982, 82, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Borecki, I.B.; Rice, J.P. Linkage Analysis of Discrete Traits. Cold Spring Harb. Protoc. 2010, 5, pdb.top69. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, D.L.; Manolio, T.A. The Emerging Importance of Genetics in Epidemiologic Research II. Issues in Study Design and Gene Mapping. Ann. Epidemiol. 1999, 9, 75–90. [Google Scholar] [CrossRef]
- Townsend, G.; Hughes, T.; Luciano, M.; Bockmann, M.; Brook, A. Genetic and Environmental Influences on Human Dental Variation: A Critical Evaluation of Studies Involving Twins. Arch. Oral Biol. 2009, 54, S45–S51. [Google Scholar] [CrossRef]
- Wang, Z.; Moult, J. SNPs, Protein Structure, and Disease. Hum. Mutat. 2001, 17, 263–270. [Google Scholar] [CrossRef]
- Manolio, T.A. Genomewide Association Studies and Assessment of the Risk of Disease. N. Engl. J. Med. 2010, 363, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Sivakumaran, S.; Agakov, F.; Theodoratou, E.; Prendergast, J.G.; Zgaga, L.; Manolio, T.; Rudan, I.; McKeigue, P.; Wilson, J.F.; Campbell, H. Abundant Pleiotropy in Human Complex Diseases and Traits. Am. J. Hum. Genet. 2011, 89, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Park, S.B.; Narita, A.; Maki, K.; Inoue, I. Genome-Wide Linkage Analysis of Mandibular Prognathism in Korean and Japanese Patients. J. Dent. Res. 2005, 84, 255–259. [Google Scholar] [CrossRef]
- Xue, F.; Wong, R.; Rabie, A.B.M. Identification of SNP Markers on 1p36 and Association Analysis of EPB41 with Mandibular Prognathism in a Chinese Population. Arch. Oral Biol. 2010, 55, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, J.H.; Frankel, W.N. The Roads from Phenotypic Variation to Gene Discovery: Mutagenesis versus QTLs. Nat. Genet. 2000, 25, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Dohmoto, A.; Shimizu, K.; Asada, Y.; Maeda, T. Quantitative Trait Loci on Chromosomes 10 and 11 Influencing Mandible Size of SMXA RI Mouse Strains. J. Dent. Res. 2002, 81, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.J.; Gómez, M.; Rey, J.A.; Ochoa, M.; Gutiérrez, S.M.; Prieto, J.C. Polymorphisms of the Noggin Gene and Mandibular Micrognathia: A First Approximation. Acta Odontológica Latinoam. 2010, 23, 13–19. [Google Scholar]
- Dai, J.; Rabie, A.B.M. Gene Therapy to Enhance Condylar Growth Using RAAV-VEGF. Angle Orthod. 2008, 78, 89–94. [Google Scholar] [CrossRef]
- Townsend, G.; Bockmann, M.; Hughes, T.; Brook, A. Genetic, Environmental and Epigenetic Influences on Variation in Human Tooth Number, Size and Shape. Odontology 2012, 100, 1–9. [Google Scholar] [CrossRef]
- Geller, F.; Feenstra, B.; Zhang, H.; Shaffer, J.R.; Hansen, T.; Esserlind, A.L.; Boyd, H.A.; Nohr, E.A.; Timpson, N.J.; Fatemifar, G.; et al. Genome-Wide Association Study Identifies Four Loci Associated with Eruption of Permanent Teeth. PLoS Genet. 2011, 7, e1002275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillas, D.; Hoggart, C.J.; Evans, D.M.; O’Reilly, P.F.; Sipilä, K.; Lähdesmäki, R.; Millwood, I.Y.; Kaakinen, M.; Netuveli, G.; Blane, D.; et al. Genome-Wide Association Study Reveals Multiple Loci Associated with Primary Tooth Development during Infancy. PLoS Genet. 2010, 6, e1000856. [Google Scholar] [CrossRef]
- Cantor, R.M.; Lange, K.; Sinsheimer, J.S. Prioritizing GWAS Results: A Review of Statistical Methods and Recommendations for Their Application. Am. J. Hum. Genet. 2010, 86, 6–22. [Google Scholar] [CrossRef] [Green Version]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the Missing Heritability of Complex Diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D. Gene-Environment-Wide Association Studies: Emerging Approaches. Nat. Rev. Genet. 2010, 11, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Leamy, L.J.; Routman, E.J.; Cheverud, J.M. Genetic Architecture of Mandible Shape in Mice: Effects of Quantitative Trait Loci Analyzed by Geometric Morphometrics. Genetics 2001, 157, 785–802. [Google Scholar] [CrossRef]
- Leamy, L.J.; Pomp, D.; Eisen, E.J.; Cheverud, J.M. Quantitative Trait Loci for Directional but Not Fluctuating Asymmetry of Mandible Characters in Mice. Genet. Res. 2000, 76, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Katz, D.C.; David Aponte, J.; Liu, W.; Green, R.M.; Mayeux, J.M.; Michael Pollard, K.; Pomp, D.; Munger, S.C.; Murray, S.A.; Roseman, C.C.; et al. Facial Shape and Allometry Quantitative Trait Locus Intervals in the Diversity Outbred Mouse Are Enriched for Known Skeletal and Facial Development Genes. PLoS ONE 2020, 15, e0233377. [Google Scholar] [CrossRef] [PubMed]
- Lone, I.M.; Iraqi, F.A. Genetics of Murine Type 2 Diabetes and Comorbidities. Mamm. Genome 2022, 33, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Ghnaim, A.; Lone, I.M.; Ben Nun, N.; Iraqi, F.A. Unraveling the Host Genetic Background Effect on Internal Organ Weight Influenced by Obesity and Diabetes Using Collaborative Cross Mice. Int. J. Mol. Sci. 2023, 24, 8201. [Google Scholar] [CrossRef]
- Lone, I.M.; Ben Nun, N.; Ghnaim, A.; Schaefer, A.S.; Houri-Haddad, Y.; Iraqi, F.A. High-Fat Diet and Oral Infection Induced Type 2 Diabetes and Obesity Development under Different Genetic Backgrounds. Anim. Model. Exp. Med. 2023, 6, 131–145. [Google Scholar] [CrossRef]
- Lone, I.M.; Zohud, O.; Nashef, A.; Kirschneck, C.; Proff, P.; Watted, N.; Iraqi, F.A. Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue. Int. J. Mol. Sci. 2023, 24, 2570. [Google Scholar] [CrossRef]
- Lone, I.M.; Midlej, K.; Ben Nun, N.; Iraqi, F.A. Intestinal Cancer Development in Response to Oral Infection with High-Fat Diet-Induced Type 2 Diabetes (T2D) in Collaborative Cross Mice under Different Host Genetic Background Effects. Mamm. Genome 2023, 34, 56–75. [Google Scholar] [CrossRef]
- Yehia, R.; Lone, I.M.; Yehia, I.; Iraqi, F.A. Studying the Pharmagenomic Effect of Portulaca Oleracea Extract on Anti-Diabetic Therapy Using the Collaborative Cross Mice. Phytomed. Plus 2023, 3, 100394. [Google Scholar] [CrossRef]
- Iraqi, F.A.; Mahajne, M.; Salaymah, Y.; Sandovski, H.; Tayem, H.; Vered, K.; Balmer, L.; Hall, M.; Manship, G.; Morahan, G.; et al. The Genome Architecture of the Collaborative Cross Mouse Genetic Reference Population. Genetics 2012, 190, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Levy, R.; Mott, R.F.; Iraqi, F.A.; Gabet, Y. Collaborative Cross Mice in a Genetic Association Study Reveal New Candidate Genes for Bone Microarchitecture. BMC Genom. 2015, 16, 1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nashef, A.; Qabaja, R.; Salaymeh, Y.; Botzman, M.; Munz, M.; Dommisch, H.; Krone, B.; Hoffmann, P.; Wellmann, J.; Laudes, M.; et al. Integration of Murine and Human Studies for Mapping Periodontitis Susceptibility. J. Dent. Res. 2018, 97, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Nashef, A.; Matthias, M.; Weiss, E.; Loos, B.G.; Jepsen, S.; van der Velde, N.; Uitterlinden, A.G.; Wellmann, J.; Berger, K.; Hoffmann, P.; et al. Translation of Mouse Model to Human Gives Insights into Periodontitis Etiology. Sci. Rep. 2020, 10, 4892 . [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorman, A.; Binenbaum, I.; Abu-Toamih Atamni, H.J.; Chatziioannou, A.; Tomlinson, I.; Mott, R.; Iraqi, F.A. Genetic Mapping of Novel Modifiers for Apc Min Induced Intestinal Polyps’ Development Using the Genetic Architecture Power of the Collaborative Cross Mice. BMC Genom. 2021, 22, 566. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | CI (cM) | Reference |
|---|---|---|
| 1 | 38–62 | [72] |
| 78–120 | [72] | |
| 12–38 | [73] | |
| 49–65 | [73] | |
| 52–53 | [74] | |
| 2 | — | [72] |
| — | [72] | |
| 21–40 | [73] | |
| 62–70 | [73] | |
| 3 | 37–53 | [72] |
| 27–61 | [72] | |
| 14–24 | [73] | |
| 69–75 | [73] | |
| 4 | 28–46 | [72] |
| 18–42 | [72] | |
| 40–60 | [73] | |
| 5 | 37–105 | [72] |
| 26–62 | [72] | |
| 29–70 | [73] | |
| 6 | 4–16 | [72] |
| 74–98 | [72] | |
| 70–100 | [72] | |
| 3–19 | [73] | |
| 99–100.2 | [74] | |
| 3.5–6 | [74] | |
| 7 | 13–65 | [72] |
| 61–77 | [72] | |
| 1–85 | [72] | |
| 37–75 | [74] | |
| 58–66 | [74] | |
| 8 | 16–36 | [73] |
| 9 | 16–38 | [72] |
| 54–88 | [72] | |
| 41–65 | [73] | |
| 10 | 9–41 | [72] |
| 65–75 | [72] | |
| 63–87 | [72] | |
| 55–83 | [72] | |
| 16–28 | [73] | |
| 31–57 | [73] | |
| 11 | 13–31 | [72] |
| 63–97 | [72] | |
| 17–55 | [72] | |
| 67–109 | [72] | |
| 51–109 | [72] | |
| 4–47 | [73] | |
| 12 | 21–33 | [72] |
| 29–70 | [72] | |
| 43–63 | [72] | |
| 32–49 | [73] | |
| 8.5–17 | [74] | |
| 13 | 79–97 | [72] |
| 7–45 | [72] | |
| 37–59 | [73] | |
| 14 | 48–64 | [72] |
| 24–64 | [72] | |
| 5–27 | [73] | |
| 38–48 | [73] | |
| 22–24 | [74] | |
| 15 | 29–53 | [72] |
| 53–85 | [72] | |
| 41–67 | [72] | |
| 11–55 | [72] | |
| 27–43 | [73] | |
| 16 | 14–44 | [72] |
| 15–33 | [73] | |
| 17 | 11–19 | [72] |
| 29–45 | [73] | |
| 45–48 | [74] | |
| 18 | 61–91 | [72] |
| 25–73 | [73] | |
| 19 | 15–61 | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lone, I.M.; Zohud, O.; Midlej, K.; Proff, P.; Watted, N.; Iraqi, F.A. Skeletal Class II Malocclusion: From Clinical Treatment Strategies to the Roadmap in Identifying the Genetic Bases of Development in Humans with the Support of the Collaborative Cross Mouse Population. J. Clin. Med. 2023, 12, 5148. https://doi.org/10.3390/jcm12155148
Lone IM, Zohud O, Midlej K, Proff P, Watted N, Iraqi FA. Skeletal Class II Malocclusion: From Clinical Treatment Strategies to the Roadmap in Identifying the Genetic Bases of Development in Humans with the Support of the Collaborative Cross Mouse Population. Journal of Clinical Medicine. 2023; 12(15):5148. https://doi.org/10.3390/jcm12155148
Chicago/Turabian StyleLone, Iqbal M., Osayd Zohud, Kareem Midlej, Peter Proff, Nezar Watted, and Fuad A. Iraqi. 2023. "Skeletal Class II Malocclusion: From Clinical Treatment Strategies to the Roadmap in Identifying the Genetic Bases of Development in Humans with the Support of the Collaborative Cross Mouse Population" Journal of Clinical Medicine 12, no. 15: 5148. https://doi.org/10.3390/jcm12155148