
IL18 Gene Polymorphism Is Associated with Total IgE in Adult Subjects with Asthma
 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
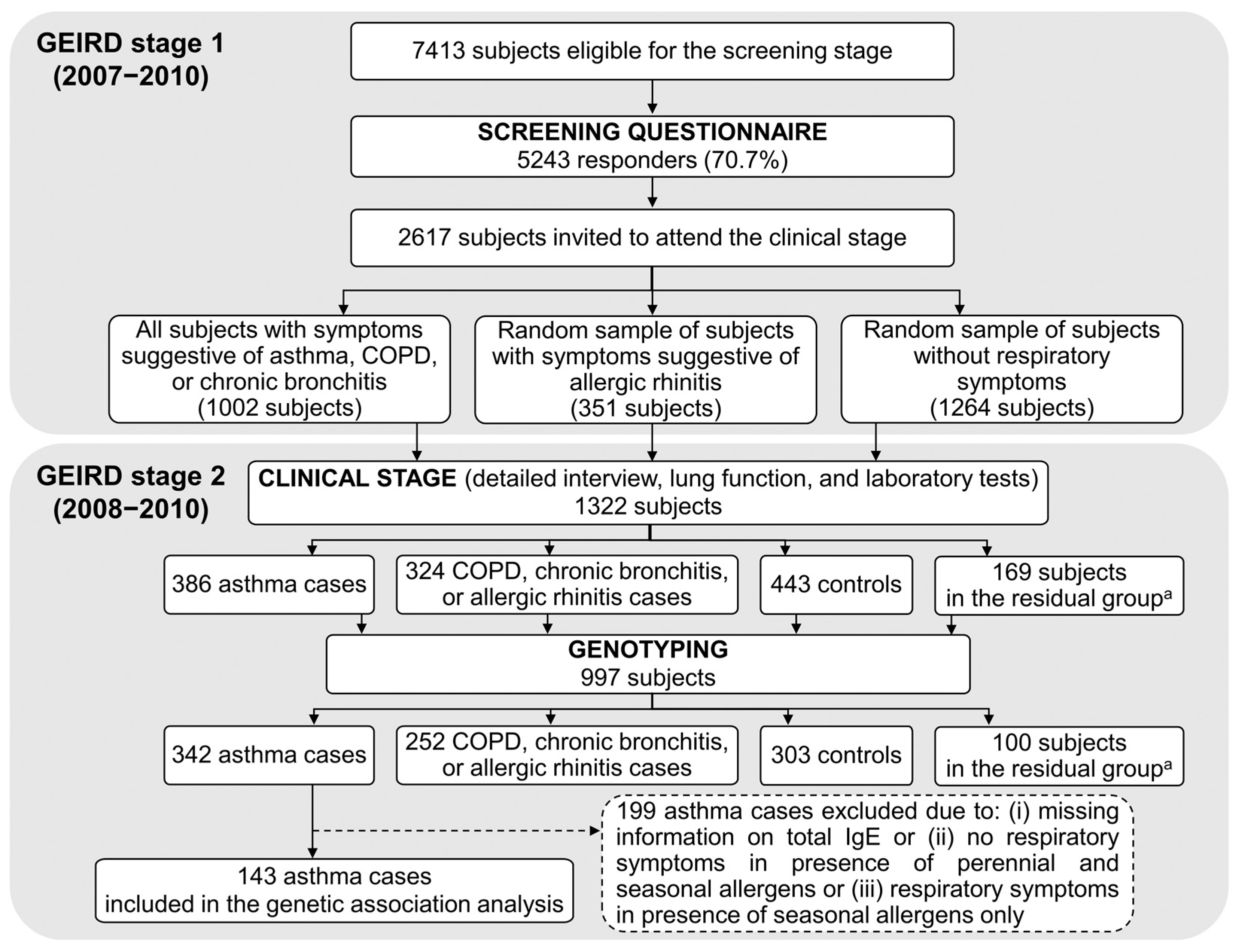
2.1. The GEIRD Survey
2.2. Identification of Cases and Controls in GEIRD
- having reported asthma at any time;
- having reported asthma-like symptoms (asthma attacks, wheezing, chest tightness, shortness of breath (SoB) at rest, SoB at night-time, SoB following strenuous activities) or the utilization of anti-asthmatic drugs in the previous 12 months, and having at least one of the following clinical characteristics:
- Positive methacholine challenge test (provocative dose (PD20) causing a 20% fall in forced expiratory volume in one second (FEV1) < 1 mg);
- Pre-bronchodilator (BD) airflow obstruction (AO) (FEV1/forced vital capacity (FVC) <lower limit of normal (LLN) [27] or <70%) and a positive reversibility test (increase in post-BD FEV1 > 12% and >200 mL with respect to pre-BD FEV1 after 400 mcg of salbutamol);
- Pre- but not post-BD AO, and post-BD FEV1 ≥ 80% predicted.
2.3. Study Subjects
2.4. Genetic Association Analysis
- Genotype failure rate ≤ 5% in all genotyped subjects (n = 997);
- Genotype failure rate ≤ 5% in all asthma cases (n = 342);
- Minimum genotype frequency ≥5% in all asthma cases (n = 342);
- Allele frequencies needed to respect Hardy–Weinberg equilibrium (HWE) in the controls (n = 303). The SNPs that were not available in the control group were excluded from the analysis [28]. p-values for testing deviation from HWE were corrected for the False Discovery Rate (FDR) using the Benjamini–Yekutieli procedure [29];
- Linkage disequilibrium (LD; squared correlation between allelic values at two loci (r2) < 0.8) in the asthma cases included in the study (n = 143) to avoid redundant testing. In case of r2 ≥ 0.8, the SNP with the highest sample size for the homozygous genotype with lower allele frequency was selected for the analysis.
2.5. Replication Analysis
3. Results
3.1. Main Characteristics of the Asthma Cases
3.2. Genetic Association and Replication Analyses
3.3. Sensitivity Analyses
4. Discussion
- SNP rs549908 in the IL18 gene was associated with total IgE in two independent samples (GEIRD and ECRHS II) of adult subjects with asthma from the general European population;
- SNP rs1063320 in the HLA-G gene was linked with this biomarker of inflammation, although statistical significance was not maintained after correcting for multiple testing in the replication sample (ECRHS II).
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kauffmann, F.; Demenais, F. Gene-environment interactions in asthma and allergic diseases: Challenges and perspectives. J. Allergy Clin. Immunol. 2012, 130, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I. ABCs of Asthma. Clin. Cornerstone 2008, 8, 9–25. [Google Scholar] [CrossRef]
- Kaur, R.; Chupp, G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J. Allergy Clin. Immunol. 2019, 144, 1–12. [Google Scholar] [CrossRef]
- Froidure, A.; Mouthuy, J.; Durham, S.R.; Chanez, P.; Sibille, Y.; Pilette, C. Asthma phenotypes and IgE responses. Eur. Respir. J. 2016, 47, 304–319. [Google Scholar] [CrossRef] [Green Version]
- Palomares, O.; Sanchez-Ramon, S.; Davila, I.; Prieto, L.; Perez de Llano, L.; Lleonart, M.; Domingo, C.; Nieto, A. dIvergEnt: How IgE Axis Contributes to the Continuum of Allergic Asthma and Anti-IgE Therapies. Int. J. Mol. Sci. 2017, 18, 1328. [Google Scholar] [CrossRef] [Green Version]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busse, W.W.; Lemanske, R.F. Asthma. N. Engl. J. Med. 2001, 344, 350–362. [Google Scholar] [CrossRef]
- Matucci, A.; Vultaggio, A.; Maggi, E.; Kasujee, I. Is IgE or eosinophils the key player in allergic asthma pathogenesis? Are we asking the right question? Respir. Res. 2018, 19, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad Al Obaidi, A.H.; Mohamed Al Samarai, A.G.; Yahya Al Samarai, A.K.; Al Janabi, J.M. The predictive value of IgE as biomarker in asthma. J. Asthma 2008, 45, 654–663. [Google Scholar] [CrossRef]
- Rath, N.; Raje, N.; Rosenwasser, L. Immunoglobulin E as a Biomarker in Asthma. Immunol. Allergy Clin. N. Am. 2018, 38, 587–597. [Google Scholar] [CrossRef]
- Omenaas, E.; Bakke, P.; Elsayed, S.; Hanoa, R.; Gulsvik, A. Total and specific serum IgE levels in adults: Relationship to sex, age and environmental factors. Clin. Exp. Allergy 1994, 24, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Carosso, A.; Bugiani, M.; Migliore, E.; Anto, J.M.; DeMarco, R. Reference values of total serum IgE and their significance in the diagnosis of allergy in young European adults. Int. Arch. Allergy Immunol. 2007, 142, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.J.; Husen, A.Z.; Khoshnaw, N.; Getta, H.A.; Hussein, Z.S.; Yassin, A.K.; Jalal, S.D.; Mohammed, R.N.; Alwan, A.F. The Effects of Smoking on IgE, Oxidative Stress and Haemoglobin Concentration. Asian Pac. J. Cancer Prev. 2020, 21, 1069–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomholt, F.K.; Nielsen, S.F.; Nordestgaard, B.G. High alcohol consumption causes high IgE levels but not high risk of allergic disease. J. Allergy Clin. Immunol. 2016, 138, 1404–1413.e13. [Google Scholar] [CrossRef] [Green Version]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, mast cells, basophils, and eosinophils. J. Allergy Clin. Immunol. 2010, 125 (Suppl. S2), S73–S80. [Google Scholar] [CrossRef]
- Weidinger, S.; Gieger, C.; Rodriguez, E.; Baurecht, H.; Mempel, M.; Klopp, N.; Gohlke, H.; Wagenpfeil, S.; Ollert, M.; Ring, J.; et al. Genome-wide scan on total serum IgE levels identifies FCER1A as novel susceptibility locus. PLoS Genet. 2008, 4, e1000166. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, M.F.; Gut, I.G.; Demenais, F.; Strachan, D.P.; Bouzigon, E.; Heath, S.; von Mutius, E.; Farrall, M.; Lathrop, M.; Cookson, W.; et al. A large-scale, consortium-based genomewide association study of asthma. N. Engl. J. Med. 2010, 363, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Imraish, A.; Abu-Thiab, T.; Zihlif, M. IL-13 and FOXO3 genes polymorphisms regulate IgE levels in asthmatic patients. Biomed. Rep. 2021, 14, 55. [Google Scholar] [CrossRef]
- Kabesch, M.; Schedel, M.; Carr, D.; Woitsch, B.; Fritzsch, C.; Weiland, S.K.; von Mutius, E. IL-4/IL-13 pathway genetics strongly influence serum IgE levels and childhood asthma. J. Allergy Clin. Immunol. 2006, 117, 269–274. [Google Scholar] [CrossRef]
- Hizawa, N.; Yamaguchi, E.; Jinushi, E.; Konno, S.; Kawakami, Y.; Nishimura, M. Increased total serum IgE levels in patients with asthma and promoter polymorphisms at CTLA4 and FCER1B. J. Allergy Clin. Immunol. 2001, 108, 74–79. [Google Scholar] [CrossRef]
- Zihlif, M.; Imraish, A.; Al-Rawashdeh, B.; Qteish, A.; Husami, R.; Husami, R.; Tahboub, F.; Jarrar, Y.; Lee, S.J. The Association of IgE Levels with ADAM33 Genetic Polymorphisms among Asthmatic Patients. J. Pers. Med. 2021, 11, 329. [Google Scholar] [CrossRef] [PubMed]
- de Marco, R.; Accordini, S.; Antonicelli, L.; Bellia, V.; Bettin, M.D.; Bombieri, C.; Bonifazi, F.; Bugiani, M.; Carosso, A.; Casali, L.; et al. The Gene-Environment Interactions in Respiratory Diseases (GEIRD) Project. Int. Arch. Allergy Immunol. 2010, 152, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Burney, P.G.; Luczynska, C.; Chinn, S.; Jarvis, D. The European Community Respiratory Health Survey. Eur. Respir. J. 1994, 7, 954–960. [Google Scholar] [CrossRef] [Green Version]
- European Community Respiratory Health Survey II Steering Committee. The European Community Respiratory Health Survey II. Eur. Respir. J. 2002, 20, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- de Marco, R.; Poli, A.; Ferrari, M.; Accordini, S.; Giammanco, G.; Bugiani, M.; Villani, S.; Ponzio, M.; Bono, R.; Carrozzi, L.; et al. The impact of climate and traffic-related NO2 on the prevalence of asthma and allergic rhinitis in Italy. Clin. Exp. Allergy 2002, 32, 1405–1412. [Google Scholar] [CrossRef]
- Accordini, S.; Calciano, L.; Bombieri, C.; Malerba, G.; Belpinati, F.; Lo Presti, A.R.; Baldan, A.; Ferrari, M.; Perbellini, L.; de Marco, R. An Interleukin 13 Polymorphism Is Associated with Symptom Severity in Adult Subjects with Ever Asthma. PLoS ONE 2016, 11, e0151292. [Google Scholar] [CrossRef] [Green Version]
- Quanjer, P.H.; Stanojevic, S.; Cole, T.J.; Baur, X.; Hall, G.L.; Culver, B.H.; Enright, P.L.; Hankinson, J.L.; Ip, M.S.; Zheng, J.; et al. Multi-ethnic reference values for spirometry for the 3-95-yr age range: The global lung function 2012 equations. Eur. Respir. J. 2012, 40, 1324–1343. [Google Scholar] [CrossRef]
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A note on exact tests of Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Statis 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- McCullagh, P.J.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman & Hall/CRC: London, UK, 1989. [Google Scholar]
- Castro-Giner, F.; Kogevinas, M.; Machler, M.; de Cid, R.; Van Steen, K.; Imboden, M.; Schindler, C.; Berger, W.; Gonzalez, J.R.; Franklin, K.A.; et al. TNFA -308G>A in two international population-based cohorts and risk of asthma. Eur. Respir. J. 2008, 32, 350–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newson, R.; the ALSPAC Study Team. Multiple-test procedures and smile plots. Stata J. 2003, 3, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Kruse, S.; Kuehr, J.; Moseler, M.; Kopp, M.V.; Kurz, T.; Deichmann, K.A.; Foster, P.S.; Mattes, J. Polymorphisms in the IL 18 gene are associated with specific sensitization to common allergens and allergic rhinitis. J. Allergy Clin. Immunol. 2003, 111, 117–122. [Google Scholar] [CrossRef]
- Nieters, A.; Linseisen, J.; Becker, N. Association of polymorphisms in Th1, Th2 cytokine genes with hayfever and atopy in a subsample of EPIC-Heidelberg. Clin. Exp. Allergy 2004, 34, 346–353. [Google Scholar] [CrossRef]
- Sebelova, S.; Izakovicova-Holla, L.; Stejskalova, A.; Schuller, M.; Znojil, V.; Vasku, A. Interleukin-18 and its three gene polymorphisms relating to allergic rhinitis. J. Hum. Genet. 2007, 52, 152–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Husseini, Z.W.; Vonk, J.M.; van den Berge, M.; Gosens, R.; Koppelman, G.H. Association of asthma genetic variants with asthma-associated traits reveals molecular pathways of eosinophilic asthma. Clin. Transl. Allergy 2023, 13, e12239. [Google Scholar] [CrossRef]
- Akhabir, L.; Sandford, A. Genetics of interleukin 1 receptor-like 1 in immune and inflammatory diseases. Curr. Genom. 2010, 11, 591–606. [Google Scholar] [CrossRef] [Green Version]
- Savenije, O.E.; Kerkhof, M.; Reijmerink, N.E.; Brunekreef, B.; de Jongste, J.C.; Smit, H.A.; Wijga, A.H.; Postma, D.S.; Koppelman, G.H. Interleukin-1 receptor-like 1 polymorphisms are associated with serum IL1RL1-a, eosinophils, and asthma in childhood. J. Allergy Clin. Immunol. 2011, 127, 750–756.e1-5. [Google Scholar] [CrossRef]
- Matloubi, M.; Ranjbar, M.; Assarehzadegan, M.A.; Fallahpour, M.; Sadeghi, F.; Soleyman-Jahi, S.; Janani, L. The Impact of Interleukin (IL)-33 Gene Polymorphisms and Environmental Factors on Risk of Asthma in the Iranian Population. Lung 2020, 198, 105–112. [Google Scholar] [CrossRef]
- Gabryelska, A.; Kuna, P.; Antczak, A.; Bialasiewicz, P.; Panek, M. IL-33 Mediated Inflammation in Chronic Respiratory Diseases-Understanding the Role of the Member of IL-1 Superfamily. Front. Immunol. 2019, 10, 692. [Google Scholar] [CrossRef] [PubMed]
- Kaplanski, G. Interleukin-18: Biological properties and role in disease pathogenesis. Immunol. Rev. 2018, 281, 138–153. [Google Scholar] [CrossRef] [Green Version]
- Sanders, N.L.; Mishra, A. Role of interleukin-18 in the pathophysiology of allergic diseases. Cytokine Growth Factor. Rev. 2016, 32, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaoka, H.; Gauvreau, G.M.; Watson, R.M.; Smith, S.G.; Dua, B.; Baatjes, A.J.; Howie, K.; Hoshino, T.; Killian, K.J.; Aizawa, H.; et al. Interleukin-18 and interleukin-18 receptor-alpha expression in allergic asthma. Eur. Respir. J. 2011, 38, 981–983. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 is a unique cytokine that stimulates both Th1 and Th2 responses depending on its cytokine milieu. Cytokine Growth Factor. Rev. 2001, 12, 53–72. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Okamura, H.; Tagawa, Y.I.; Iwakura, Y.; Nakanishi, K. Interleukin 18 together with interleukin 12 inhibits IgE production by induction of interferon-gamma production from activated B cells. Proc. Natl. Acad. Sci. USA 1997, 94, 3948–3953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, T.; Mizutani, H.; Tsutsui, H.; Noben-Trauth, N.; Yamanaka, K.; Tanaka, M.; Izumi, S.; Okamura, H.; Paul, W.E.; Nakanishi, K. IL-18 induction of IgE: Dependence on CD4+ T cells, IL-4 and STAT6. Nat. Immunol. 2000, 1, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T.; Nakanishi, K. Roles of IL-18 in basophils and mast cells. Allergol. Int. 2006, 55, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, T.; Tsutsui, H.; Tominaga, K.; Hoshino, K.; Okamura, H.; Akira, S.; Paul, W.E.; Nakanishi, K. IL-18, although antiallergic when administered with IL-12, stimulates IL-4 and histamine release by basophils. Proc. Natl. Acad. Sci. USA 1999, 96, 13962–13966. [Google Scholar] [CrossRef] [Green Version]
- White, S.R. Human leucocyte antigen-G: Expression and function in airway allergic disease. Clin. Exp. Allergy 2012, 42, 208–217. [Google Scholar] [CrossRef]
- Negrini, S.; Contini, P.; Murdaca, G.; Puppo, F. HLA-G in Allergy: Does It Play an Immunoregulatory Role? Front. Immunol. 2021, 12, 789684. [Google Scholar] [CrossRef]
- Granada, M.; Wilk, J.B.; Tuzova, M.; Strachan, D.P.; Weidinger, S.; Albrecht, E.; Gieger, C.; Heinrich, J.; Himes, B.E.; Hunninghake, G.M.; et al. A genome-wide association study of plasma total IgE concentrations in the Framingham Heart Study. J. Allergy Clin. Immunol. 2012, 129, 840–845.e21. [Google Scholar] [CrossRef] [Green Version]
- Yatagai, Y.; Sakamoto, T.; Masuko, H.; Kaneko, Y.; Yamada, H.; Iijima, H.; Naito, T.; Noguchi, E.; Hirota, T.; Tamari, M.; et al. Genome-wide association study for levels of total serum IgE identifies HLA-C in a Japanese population. PLoS ONE 2013, 8, e80941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.R.; Nicodemus-Johnson, J.; Laxman, B.; Denner, D.R.; Naureckas, E.T.; Hogarth, D.K.; Stern, R.; Minc, A.; Solway, J.; Sperling, A.; et al. Elevated levels of soluble humanleukocyte antigen-G in the airways are a marker for a low-inflammatory endotype of asthma. J. Allergy Clin. Immunol. 2017, 140, 857–860. [Google Scholar] [CrossRef] [Green Version]
- Ciprandi, G.; Corsico, A.; Pisati, P. Serum-soluble HLA-G is associated with specific IgE in patients with allergic rhinitis and asthma. Inflammation 2014, 37, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; DeAmici, M. Soluble HLA-G serum levels depend on allergy type and IgE levels. Allergy Rhinol. Provid. 2014, 5, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Ribeyre, C.; Carlini, F.; Rene, C.; Jordier, F.; Picard, C.; Chiaroni, J.; Abi-Rached, L.; Gouret, P.; Marin, G.; Molinari, N.; et al. HLA-G Haplotypes Are Differentially Associated with Asthmatic Features. Front. Immunol. 2018, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.C.; Arruda, L.K.P.; Oliveira, F.R.; Massaro, J.D.; Aquino, B.J.; Paz, M.A.; Castelli, E.C.; Mendes-Junior, C.T.; Donadi, E.A. Human leukocyte antigen-G 3’ untranslated region polymorphisms are associated with asthma severity. Mol. Immunol. 2018, 101, 500–506. [Google Scholar] [CrossRef] [Green Version]
- Castelli, E.C.; Veiga-Castelli, L.C.; Yaghi, L.; Moreau, P.; Donadi, E.A. Transcriptional and posttranscriptional regulations of the HLA-G gene. J. Immunol. Res. 2014, 2014, 734068. [Google Scholar] [CrossRef] [Green Version]
- Martins, T.B.; Bandhauer, M.E.; Bunker, A.M.; Roberts, W.L.; Hill, H.R. New childhood and adult reference intervals for total IgE. J. Allergy Clin. Immunol. 2014, 133, 589–591. [Google Scholar] [CrossRef]
- Kim, H.Y.; Choi, J.; Ahn, K.; Hahm, M.I.; Lee, S.Y.; Kim, W.K.; Chae, Y.; Park, Y.M.; Han, M.Y.; Lee, K.J.; et al. Reference Values and Utility of Serum Total Immunoglobulin E for Predicting Atopy and Allergic Diseases in Korean Schoolchildren. J. Korean Med. Sci. 2017, 32, 803–809. [Google Scholar] [CrossRef]
- Litonjua, A.A.; Celedon, J.C.; Hausmann, J.; Nikolov, M.; Sredl, D.; Ryan, L.; Platts-Mills, T.A.; Weiss, S.T.; Gold, D.R. Variation in total and specific IgE: Effects of ethnicity and socioeconomic status. J. Allergy Clin. Immunol. 2005, 115, 751–757. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Genetic Association Analysis | Replication Analysis | p-Value a,b | ||
|---|---|---|---|---|
| Sample, n | 143 | 842 | - | |
| Female, % | 49.7 | 53.9 | 0.344 | |
| Age (years), median (IQR) | 42.1 (34.9, 48.5) | 42.9 (37.2, 48.8) | 0.204 | |
| BMI, median (IQR) | 24.5 (22.0, 27.1) | 25.1 (22.8, 28.1) | 0.024 | |
| Tobacco smoking, % | Never | 48.9 | 40.3 | 0.143 |
| Past | 22.4 | 27.6 | ||
| Current | 28.7 | 32.1 | ||
| Nasal allergies, % | Absent | 30.1 | 55.2 | <0.001 |
| Present | 69.2 | 44.1 | ||
| Missing | 0.7 | 0.7 | ||
| Eczema/Skin allergies, % | 34.3 | 51.8 | <0.001 | |
| Itchy rash, % | 14.0 | 23.2 | 0.013 | |
| Pre-BD FEV1 % predicted, median (IQR) | 95.5 (85.7, 108.3) | 97.8 (86.5, 108.4) | 0.388 | |
| Pre-BD FVC % predicted, median (IQR) | 101.7 (92.2, 110.0) | 99.2 (88.8, 108.7) | 0.130 | |
| Pre-BD FEV1/FVC % predicted, median (IQR) | 93.3 (89.4, 100.4) | 95.6 (89.5, 104.3) | 0.066 | |
| Total IgE (kU/L), geometric mean (95%CI) | 111.5 (91.5, 135.8) | 54.1 (49.0, 59.7) | 0.003 |
| Genetic Association Analysis | Replication Analysis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene or Gene Region | SNP | Genotype | Sample (n = 143) | Ratio of Expected Total IgE a (95%CI) | Two-Sided p-Value | Genotype b | Sample (n = 842) | Ratio of Expected Total IgE c (95%CI) | Two-Sided p-Value | One-Sided p-Value d | FDR-Corrected One-Sided p-Value |
| IL18 | rs549908 | TT | 73 | 1.00 | - | TT | 379 | 1.00 | - | - | - |
| TG | 60 | 1.43 (1.07, 1.92) | 0.016 | TG | 375 | 1.32 (1.07, 1.62) | 0.008 | 0.004 | 0.054 | ||
| GG | 9 | 2.05 (additive effect) | - | GG | 88 | 1.74 (additive effect) | - | - | - | ||
| HLA-G | rs1063320 | CC | 46 | 1.00 | - | CC | 211 | 1.00 | - | - | - |
| CG | 66 | 0.78 (0.61, 0.998) | 0.048 | CG | 413 | 0.84 (0.71, 1.01) | 0.061 | 0.031 | 0.198 | ||
| GG | 31 | 0.60 (additive effect) | - | GG | 218 | 0.71 (additive effect) | - | - | - | ||
| NPSR1 | rs898070 | GG | 60 | 1.00 | - | GG | 329 | 1.00 | - | - | - |
| AG | 63 | 0.61 (0.48, 0.79) | 0.0001 | AG | 409 | 1.07 (0.75, 1.51) | 0.719 | 0.641 | 0.757 | ||
| AA | 19 | 0.38 (additive effect) | - | AA | 104 | 1.14 (additive effect) | - | - | |||
| NOS1 | rs12830203 | CC | 53 | 1.00 | - | CC | 419 | 1.00 | - | - | - |
| TC | 75 | 0.66 (0.50, 0.87) | 0.004 | TC | 339 | 1.07 (0.87, 1.31) | 0.527 | 0.736 | 0.798 | ||
| TT | 15 | 0.44 (additive effect) | - | TT | 84 | 1.14 (additive effect) | - | - | - | ||
| IL2RB | rs2072862 | GG | 53 | 1.00 | - | GG | 403 | 1.00 | - | - | - |
| AG | 68 | 1.47 (1.13, 1.90) | 0.004 | AG | 345 | 1.11 (0.88, 1.40) | 0.362 | 0.181 | 0.470 | ||
| AA | 21 | 2.16 (additive effect) | - | AA | 94 | 1.24 (additive effect) | - | - | - | ||
| SMAD3A | rs11635145 | AA | 46 | 1.00 | - | AA | 235 | 1.00 | - | - | - |
| AG | 72 | 1.42 (1.11, 1.82) | 0.005 | AG | 423 | 0.85 (0.67, 1.08) | 0.192 | 0.904 | 0.904 | ||
| GG | 25 | 2.03 (additive effect) | - | GG | 184 | 0.73 (additive effect) | - | - | - | ||
| NPNT | rs6817700 | GG | 37 | 1.00 | - | GG | 231 | 1.00 | - | - | - |
| AG | 79 | 1.45 (1.11, 1.89) | 0.006 | AG | 422 | 1.03 (0.89, 1.18) | 0.724 | 0.362 | 0.748 | ||
| AA | 27 | 2.10 (additive effect) | - | AA | 189 | 1.05 (additive effect) | - | - | - | ||
| NOS1 | rs733334 | GG | 35 | 1.00 | - | CC | 214 | 1.00 | - | - | - |
| AG | 73 | 0.71 (0.55, 0.91) | 0.008 | TC | 427 | 1.003 (0.83, 1.21) | 0.976 | 0.512 | 0.748 | ||
| AA | 35 | 0.51 (additive effect) | - | TT | 201 | 1.01 (additive effect) | - | - | - | ||
| IL1RL2 | rs10184597 | CC | 78 | 1.00 | - | CC | 437 | 1.00 | - | - | - |
| TC | 54 | 1.47 (1.10, 1.92) | 0.009 | TC | 355 | 1.09 (0.92, 1.29) | 0.331 | 0.166 | 0.470 | ||
| TT | 10 | 2.17 (additive effect) | - | TT | 50 | 1.18 (additive effect) | - | - | - | ||
| GPR126 | rs9389986 | TT | 58 | 1.00 | - | TT | 390 | 1.00 | - | - | - |
| AT | 61 | 0.74 (0.56, 0.96) | 0.023 | AT | 367 | 0.98 (0.81, 1.18) | 0.814 | 0.407 | 0.748 | ||
| AA | 24 | 0.54 (additive effect) | - | AA | 85 | 0.96 (additive effect) | - | - | - | ||
| VDR | rs4760648 | CC | 43 | 1.00 | - | CC | 272 | 1.00 | - | - | - |
| TC | 68 | 0.76 (0.60, 0.98) | 0.032 | TC | 401 | 0.93 (0.82, 1.05) | 0.232 | 0.116 | 0.470 | ||
| TT | 29 | 0.58 (additive effect) | - | TT | 169 | 0.86 (additive effect) | - | - | - | ||
| CHRNA3 | rs578776 | CC | 74 | 1.00 | - | GG | 443 | 1.00 | - | - | - |
| TC | 57 | 1.36 (1.01, 1.84) | 0.042 | AG | 335 | 0.97 (0.70, 1.34) | 0.851 | 0.575 | 0.748 | ||
| TT | 10 | 1.86 (additive effect) | - | AA | 64 | 0.94 (additive effect) | - | - | - | ||
| TNS1 | rs3791978 | TT | 40 | 1.00 | - | AA | 236 | 1.00 | - | - | - |
| TG | 71 | 0.77 (0.59, 0.998) | 0.048 | AC | 414 | 1.01 (0.89, 1.16) | 0.850 | 0.575 | 0.748 | ||
| GG | 32 | 0.59 (additive effect) | - | CC | 192 | 1.03 (additive effect) | - | - | - | ||
| TT (n = 73) | TG (n = 60) | GG (n = 9) | p-Value b | ||
|---|---|---|---|---|---|
| Sample, n | 52.0 | 45.0 | 66.7 | 0.439 | |
| Female, % | 42.0 (36.1, 48.3) | 42.5 (31.1, 48.1) | 44.7 (39.9, 48.5) | 0.380 | |
| Age (years), median (IQR) | 24.5 (22.1, 26.7) | 24.4 (21.7, 27.3) | 25.5 (23.1, 29.4) | 0.693 | |
| BMI, median (IQR) | 54.8 | 45.0 | 22.2 | 0.278 | |
| Tobacco smoking, % | Never | 20.6 | 21.7 | 44.4 | |
| Past | 24.7 | 33.3 | 33.3 | ||
| Current | 74.0 | 66.7 | 44.4 | 0.413 | |
| Nasal allergies, % | Absent | 35.6 | 23.3 | 33.3 | 0.342 |
| Present | 63.0 | 76.7 | 66.7 | ||
| Missing | 1.4 | 0.0 | 0.0 | ||
| Eczema/Skin allergies, % | 37.0 | 35.0 | 11.1 | 0.344 | |
| Itchy rash, % | 13.7 | 16.7 | 0.0 | 0.508 | |
| Pre-BD FEV1 % predicted, median (IQR) | 95.5 (84.7, 106.3) | 97.8 (90.1, 108.6) | 84.0 (70.5, 93.5) | 0.034 | |
| Pre-BD FVC % predicted, median (IQR) | 101.1 (92.2, 109.2) | 102.2 (94.7, 110.7) | 90.5 (85.7, 101.4) | 0.199 | |
| Pre-BD FEV1/FVC % predicted, median (IQR) | 94.1 (90.9, 100.8) | 94.1 (90.2, 100.4) | 88.2 (81.7, 93.2) | 0.059 | |
| Total IgE (kU/L), geometric mean (95%CI) | 88.4 (68.0, 115.1) | 151.8 (110.3, 209.0) | 99.9 (41.8, 238.8) | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lando, V.; Calciano, L.; Minelli, C.; Bombieri, C.; Ferrari, M.; Malerba, G.; Margagliotti, A.; Murgia, N.; Nicolis, M.; Olivieri, M.; et al. IL18 Gene Polymorphism Is Associated with Total IgE in Adult Subjects with Asthma. J. Clin. Med. 2023, 12, 3963. https://doi.org/10.3390/jcm12123963
Lando V, Calciano L, Minelli C, Bombieri C, Ferrari M, Malerba G, Margagliotti A, Murgia N, Nicolis M, Olivieri M, et al. IL18 Gene Polymorphism Is Associated with Total IgE in Adult Subjects with Asthma. Journal of Clinical Medicine. 2023; 12(12):3963. https://doi.org/10.3390/jcm12123963
Chicago/Turabian StyleLando, Valentina, Lucia Calciano, Cosetta Minelli, Cristina Bombieri, Marcello Ferrari, Giovanni Malerba, Antonino Margagliotti, Nicola Murgia, Morena Nicolis, Mario Olivieri, and et al. 2023. "IL18 Gene Polymorphism Is Associated with Total IgE in Adult Subjects with Asthma" Journal of Clinical Medicine 12, no. 12: 3963. https://doi.org/10.3390/jcm12123963