
A Comprehensive Analysis of the Expression Profiles of KCTD Proteins in Acute Lymphoblastic Leukemia: Evidence of Selective Expression of KCTD1 in T-ALL
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Patients Sample and Cell Lines
2.3. RNA Extraction, RT-PCR, and Real-Time PCR
- RPS18fw = 5′-CGATGGGCGGCGGAAAATA-3′;
- RPS18rev = 5′-CTGCTTTCCTCAACACCACA-3′;
- KCTD1fw= 5′- TGAGTGGTGAGGACACAGTC-3′;
- KCTD1rev= 5′- CCTGGTCCCTGCCTAAAGAA-3′;
- KCTD3fw= 5′-CATCAGTCCAGCAACTTCCG-3′;
- KCTD3rev= 5′-AAGGCCTCATGGAACCGTAA-3′;
- KCTD12fw = 5′-TTTCTCCAAGCCCAGCAAAC-3′;
- KCTD12rev= 5′-TACAGATAGGCAGCCCTTGG-3′;
- KCTD11fw= 5′-GAGGGGAGCCCACATTTTCA-3;
- KCTD11rev= 5′-GAGTCTAGTCGGAAGCCGTG-3′;
- KCTD15fw = 5′-TGTCATGGCAACAGAACGTG-3′;
- KCTD15rev = 5′-CAGAGATCCCACCGCTGTAT-3′.
2.4. Protein Extraction
2.5. Western Blotting Analysis
2.6. KCTD1 Knockdown
2.7. Cell Cycle Analyses
2.8. Transcriptomic Analyses
3. Results
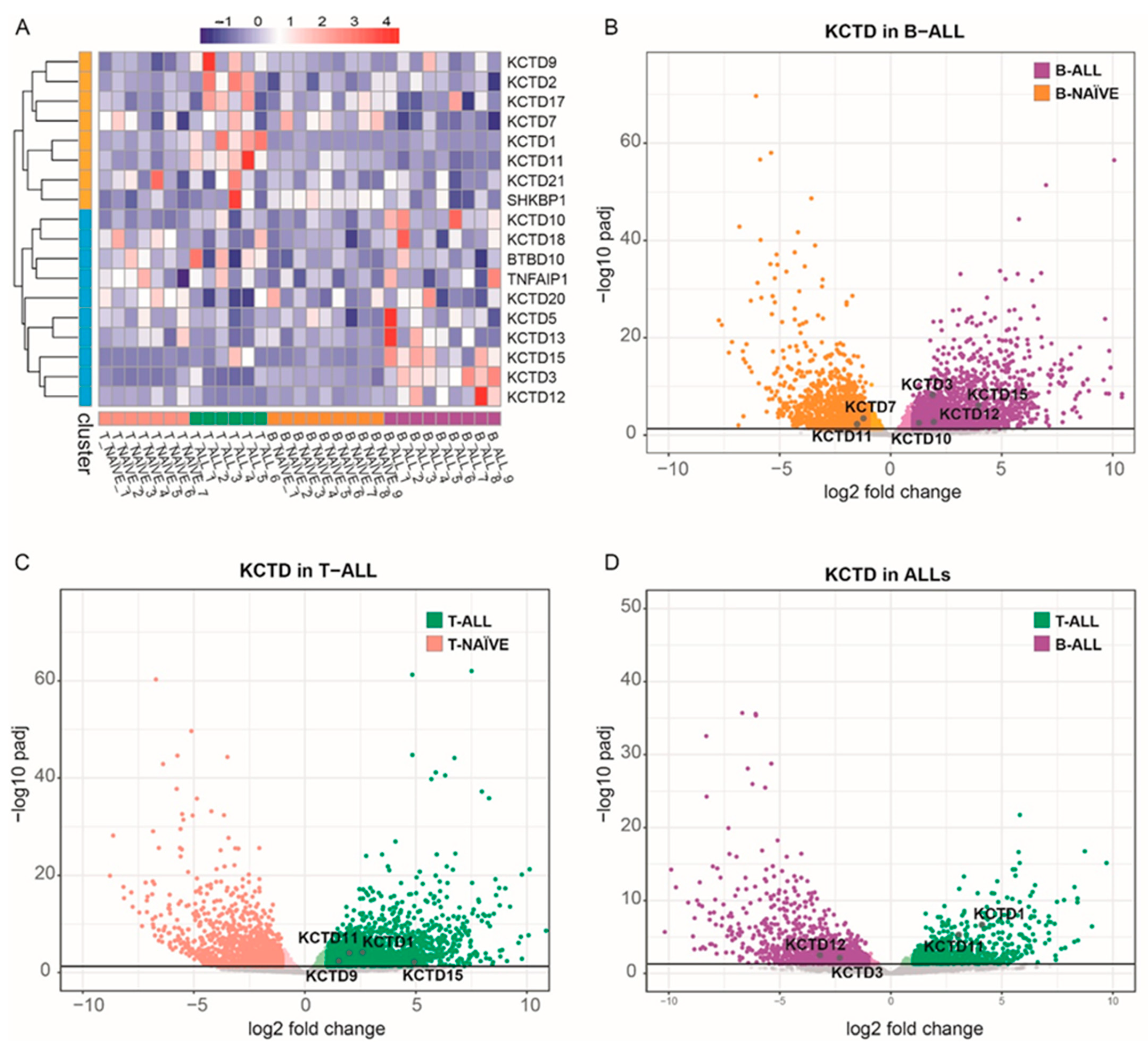
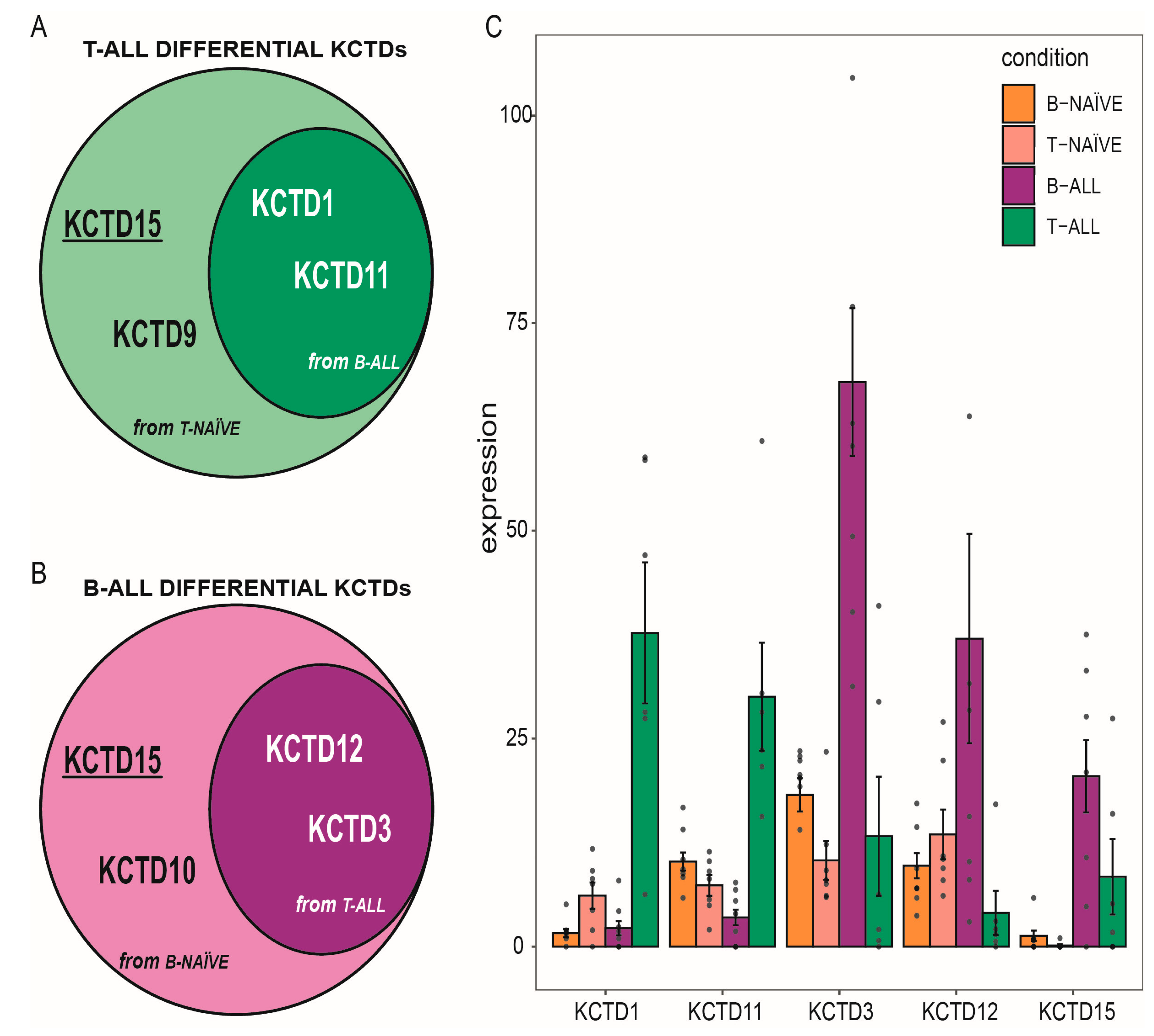
3.1. KCTD Family Transcriptomic Profile in ALLs
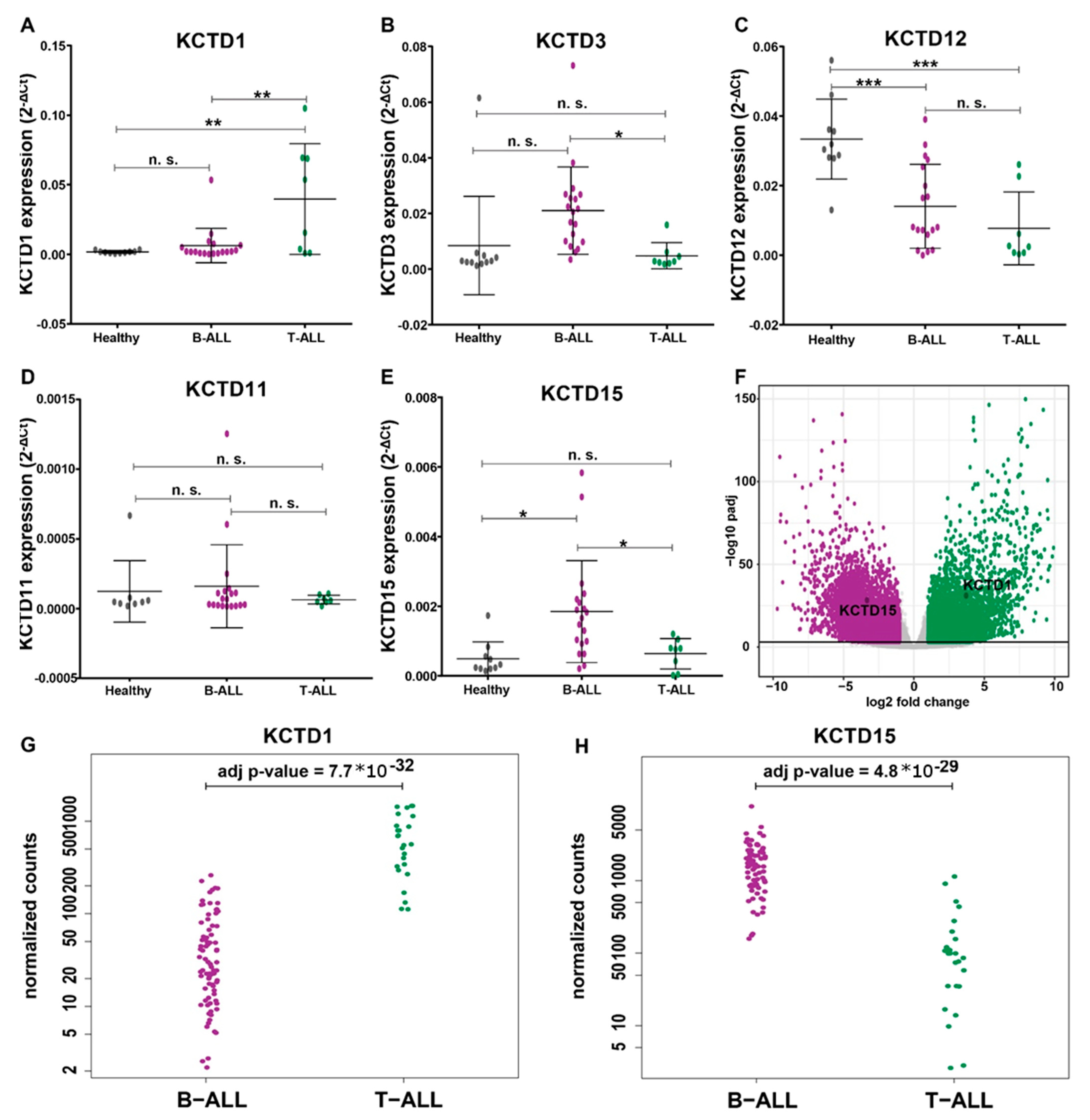
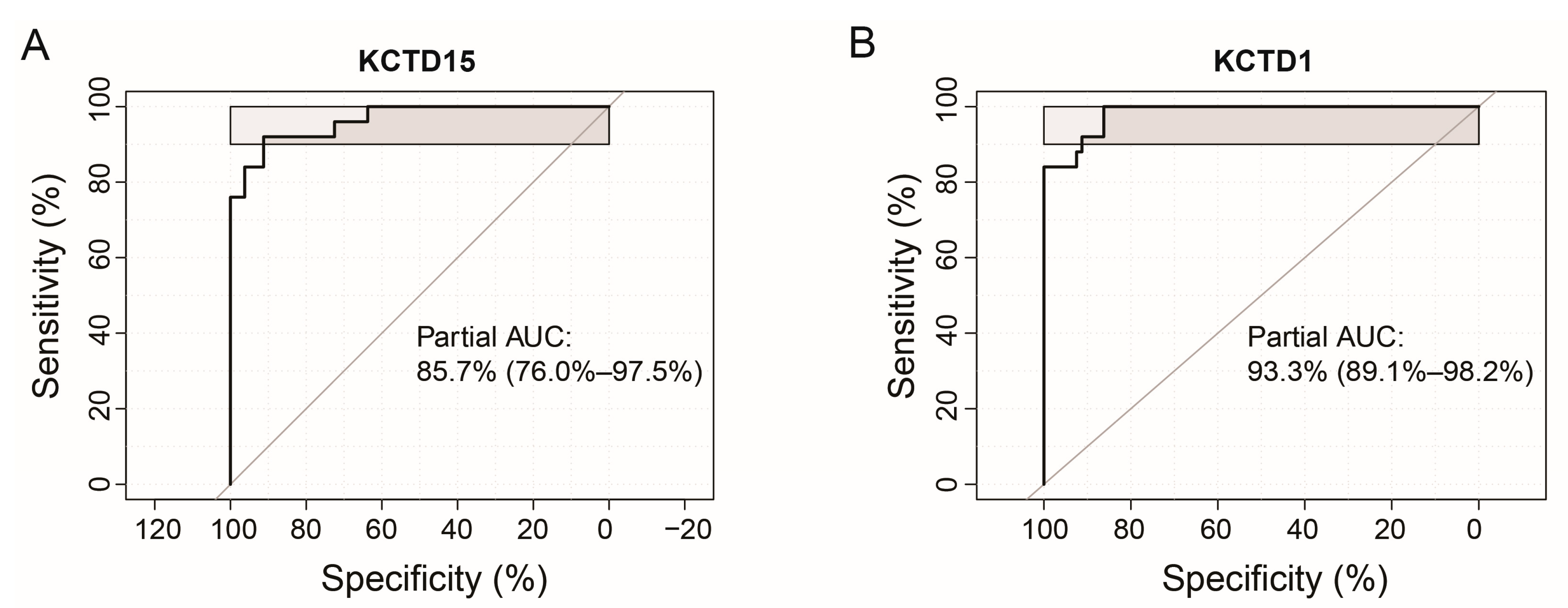
3.2. KCTDs’ Expression Validation in B-ALL and T-ALL Cohorts
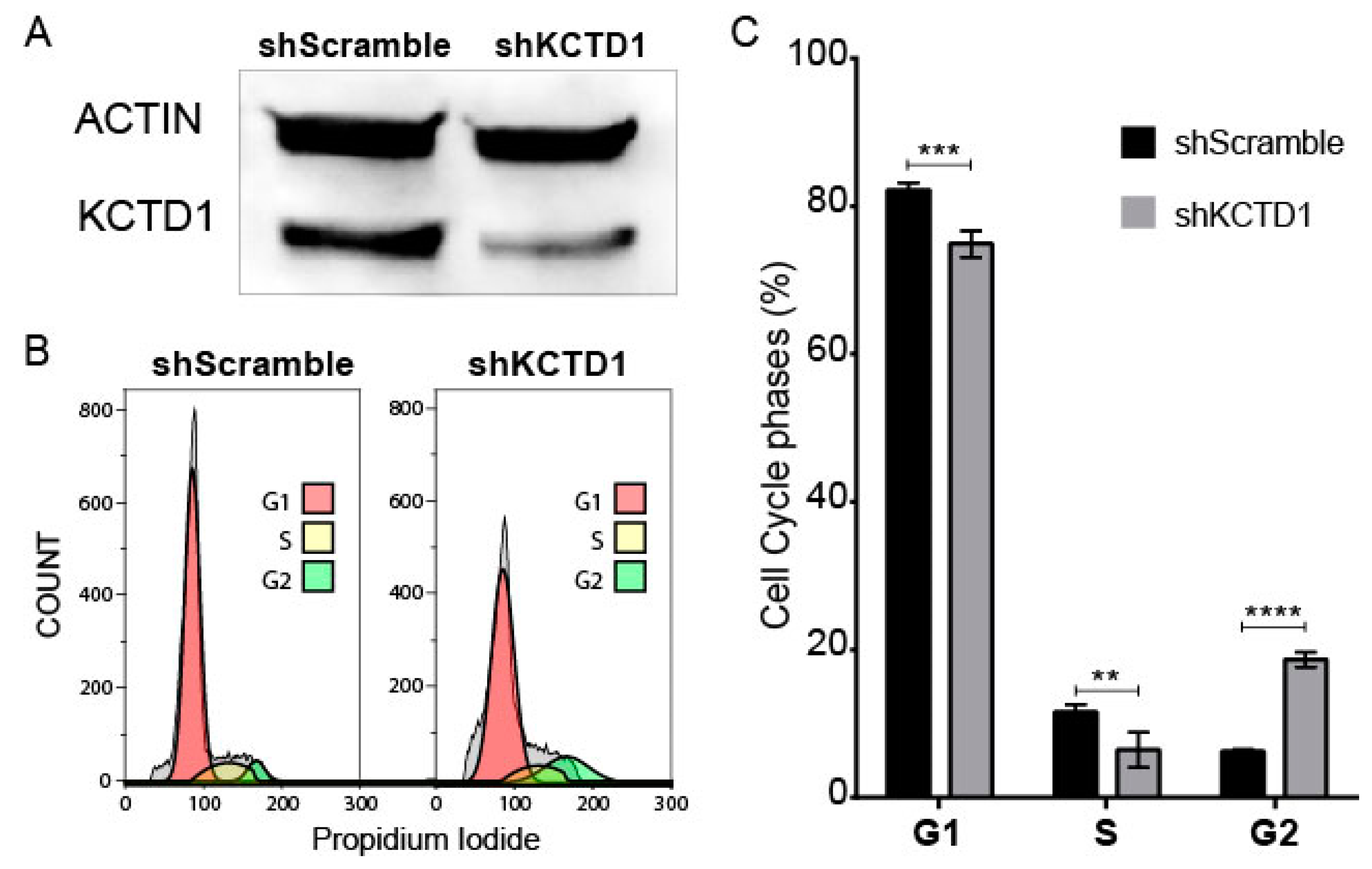
3.3. KCTD1 Knock down Determines a Variation in the Cell Cycle of Jurkat Cells
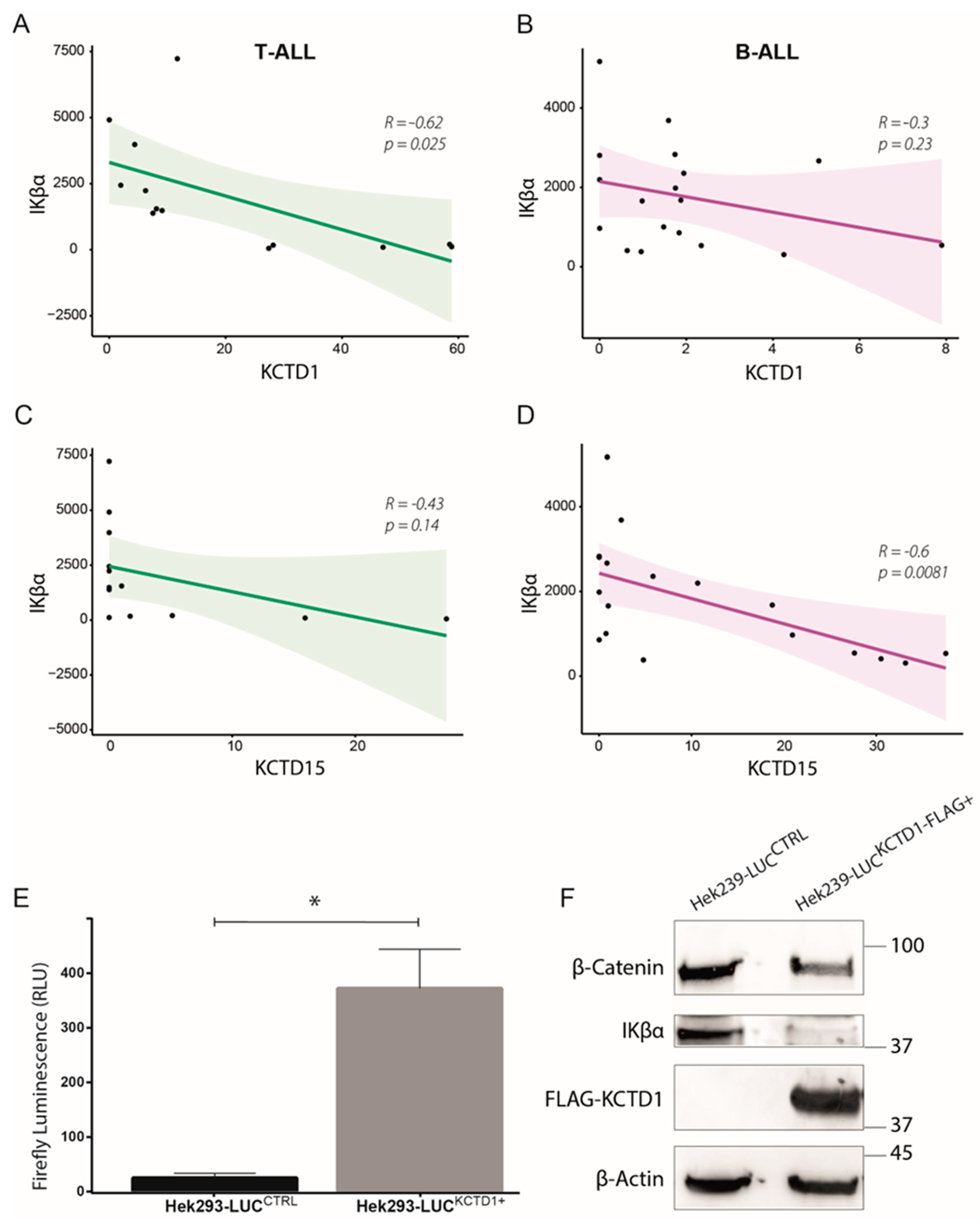
3.4. KCTD1 as Possible Regulator of the NF-κB Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Inaba, H.; Mullighan, C.G. Pediatric Acute Lymphoblastic Leukemia. Haematologica 2020, 105, 2524–2539. [Google Scholar] [CrossRef] [PubMed]
- PDQ Pediatric Treatment Editorial Board. Childhood Acute Lymphoblastic Leukemia Treatment (PDQ®): Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute: Bethesda, MD, USA, 2002. [Google Scholar]
- Huang, F.-L.; Liao, E.-C.; Li, C.-L.; Yen, C.-Y.; Yu, S.-J. Pathogenesis of Pediatric B-Cell Acute Lymphoblastic Leukemia: Molecular Pathways and Disease Treatments. Oncol. Lett. 2020, 20, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Cordo’, V.; van der Zwet, J.C.G.; Canté-Barrett, K.; Pieters, R.; Meijerink, J.P.P. T-Cell Acute Lymphoblastic Leukemia: A Roadmap to Targeted Therapies. Blood Cancer Discov. 2021, 2, 19–31. [Google Scholar] [CrossRef]
- Teachey, D.T.; Pui, C.-H. Comparative Features and Outcomes between Paediatric T-Cell and B-Cell Acute Lymphoblastic Leukaemia. Lancet Oncol. 2019, 20, e142–e154. [Google Scholar] [CrossRef] [PubMed]
- Teachey, D.T.; O’Connor, D. How I Treat Newly Diagnosed T-Cell Acute Lymphoblastic Leukemia and T-Cell Lymphoblastic Lymphoma in Children. Blood 2020, 135, 159–166. [Google Scholar] [CrossRef]
- Vlierberghe, P.V.; Ferrando, A. The Molecular Basis of T Cell Acute Lymphoblastic Leukemia. J. Clin. Investig. 2012, 122, 3398–3406. [Google Scholar] [CrossRef]
- Piya, S.; Yang, Y.; Bhattacharya, S.; Sharma, P.; Ma, H.; Mu, H.; He, H.; Ruvolo, V.; Baran, N.; Davis, R.E.; et al. Targeting the NOTCH1-MYC-CD44 Axis in Leukemia-Initiating Cells in T-ALL. Leukemia 2022, 36, 1261–1273. [Google Scholar] [CrossRef]
- Richter-Pechańska, P.; Kunz, J.B.; Rausch, T.; Erarslan-Uysal, B.; Bornhauser, B.; Frismantas, V.; Assenov, Y.; Zimmermann, M.; Happich, M.; von Knebel-Doeberitz, C.; et al. Pediatric T-ALL Type-1 and Type-2 Relapses Develop along Distinct Pathways of Clonal Evolution. Leukemia 2022, 36, 1759–1768. [Google Scholar] [CrossRef]
- Greaves, M. A Causal Mechanism for Childhood Acute Lymphoblastic Leukaemia. Nat. Rev. Cancer 2018, 18, 471–484. [Google Scholar] [CrossRef]
- Swaminathan, S.; Klemm, L.; Park, E.; Papaemmanuil, E.; Ford, A.; Kweon, S.-M.; Trageser, D.; Hasselfeld, B.; Henke, N.; Mooster, J.; et al. Mechanisms of Clonal Evolution in Childhood Acute Lymphoblastic Leukemia. Nat. Immunol. 2015, 16, 766–774. [Google Scholar] [CrossRef]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A Guide to Cancer Immunotherapy: From T Cell Basic Science to Clinical Practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Smaldone, G.; Beneduce, G.; Incoronato, M.; Pane, K.; Franzese, M.; Coppola, L.; Cordella, A.; Parasole, R.; Ripaldi, M.; Nassa, G.; et al. KCTD15 Is Overexpressed in Human Childhood B-Cell Acute Lymphoid Leukemia. Sci. Rep. 2019, 9, 20108. [Google Scholar] [CrossRef] [PubMed]
- Smaldone, G.; Coppola, L.; Pane, K.; Franzese, M.; Beneduce, G.; Parasole, R.; Menna, G.; Vitagliano, L.; Salvatore, M.; Mirabelli, P. KCTD15 Deregulation Is Associated with Alterations of the NF-ΚB Signaling in Both Pathological and Physiological Model Systems. Sci. Rep. 2021, 11, 18237. [Google Scholar] [CrossRef] [PubMed]
- Angrisani, A.; Di Fiore, A.; De Smaele, E.; Moretti, M. The Emerging Role of the KCTD Proteins in Cancer. Cell Commun. Signal. 2021, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Esposito, L.; Balasco, N.; Smaldone, G.; Berisio, R.; Ruggiero, A.; Vitagliano, L. AlphaFold-Predicted Structures of KCTD Proteins Unravel Previously Undetected Relationships among the Members of the Family. Biomolecules 2021, 11, 1862. [Google Scholar] [CrossRef]
- Imbert, V.; Peyron, J.-F. NF-ΚB in Hematological Malignancies. Biomedicines 2017, 5, 27. [Google Scholar] [CrossRef]
- Buono, L.; Iside, C.; De Matteo, A.; Stellato, P.; Beneduce, G.; de Vera d’Aragona, R.P.; Parasole, R.; Salvatore, M.; Smaldone, G.; Mirabelli, P. Specific LncRNA Signatures Discriminate Childhood Acute Leukaemias: A Pilot Study. Cancer Cell Int. 2022, 22, 373. [Google Scholar] [CrossRef]
- Mirabelli, P.; Incoronato, M.; Coppola, L.; Infante, T.; Parente, C.A.; Nicolai, E.; Soricelli, A.; Salvatore, M. SDN Biobank: Bioresource of Human Samples Associated with Functional and/or Morphological Bioimaging Results for the Study of Oncological, Cardiological, Neurological, and Metabolic Diseases. Open J. Bioresour. 2017, 4, 2. [Google Scholar] [CrossRef]
- Carrizzo, A.; Iside, C.; Nebbioso, A.; Carafa, V.; Damato, A.; Sciarretta, S.; Frati, G.; Di Nonno, F.; Valenti, V.; Ciccarelli, M.; et al. SIRT1 Pharmacological Activation Rescues Vascular Dysfunction and Prevents Thrombosis in MTHFR Deficiency. Cell. Mol. Life Sci. 2022, 79, 410. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Buono, L.; Corbacho, J.; Naranjo, S.; Almuedo-Castillo, M.; Moreno-Marmol, T.; de la Cerda, B.; Sanabria-Reinoso, E.; Polvillo, R.; Díaz-Corrales, F.-J.; Bogdanovic, O.; et al. Analysis of Gene Network Bifurcation during Optic Cup Morphogenesis in Zebrafish. Nat. Commun. 2021, 12, 3866. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-Catenin Signalling: Function, Biological Mechanisms, and Therapeutic Opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, C.; Wang, F.; Huang, W.; Liang, Z.; Xiao, Y.; Wei, K.; Wan, Z.; Hu, X.; Xiang, S.; et al. KCTD1 Suppresses Canonical Wnt Signaling Pathway by Enhancing β-Catenin Degradation. PLoS ONE 2014, 9, e94343. [Google Scholar] [CrossRef] [PubMed]
- Skoblov, M.; Marakhonov, A.; Marakasova, E.; Guskova, A.; Chandhoke, V.; Birerdinc, A.; Baranova, A. Protein Partners of KCTD Proteins Provide Insights about Their Functional Roles in Cell Differentiation and Vertebrate Development. BioEssays 2013, 35, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Balasco, N.; Smaldone, G.; Vitagliano, L. The Structural Versatility of the BTB Domains of KCTD Proteins and Their Recognition of the GABAB Receptor. Biomolecules 2019, 9, 323. [Google Scholar] [CrossRef]
- Smaldone, G.; Pirone, L.; Pedone, E.; Marlovits, T.; Vitagliano, L.; Ciccarelli, L. The BTB Domains of the Potassium Channel Tetramerization Domain Proteins Prevalently Assume Pentameric States. FEBS Lett. 2016, 590, 1663–1671. [Google Scholar] [CrossRef]
- Smaldone, G.; Pirone, L.; Balasco, N.; Gaetano, S.D.; Pedone, E.M.; Vitagliano, L. Cullin 3 Recognition Is Not a Universal Property among KCTD Proteins. PLoS ONE 2015, 10, e0126808. [Google Scholar] [CrossRef]
- Pirone, L.; Smaldone, G.; Spinelli, R.; Barberisi, M.; Beguinot, F.; Vitagliano, L.; Miele, C.; Di Gaetano, S.; Raciti, G.A.; Pedone, E. KCTD1: A Novel Modulator of Adipogenesis through the Interaction with the Transcription Factor AP2α. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 158514. [Google Scholar] [CrossRef]
- Zheng, S.; Abreu, N.; Levitz, J.; Kruse, A.C. Structural Basis for KCTD-Mediated Rapid Desensitization of GABAB Signalling. Nature 2019, 567, 127–131. [Google Scholar] [CrossRef]
- Dos Santos, N.R.; Ghezzo, M.N.; Da Silva, R.C.; Fernandes, M.T. NF-ΚB in T-Cell Acute Lymphoblastic Leukemia: Oncogenic Functions in Leukemic and in Microenvironmental Cells. Cancers 2010, 2, 1838. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buono, L.; Iside, C.; Pecoraro, G.; De Matteo, A.; Beneduce, G.; Penta de Vera d’Aragona, R.; Parasole, R.; Mirabelli, P.; Vitagliano, L.; Salvatore, M.; et al. A Comprehensive Analysis of the Expression Profiles of KCTD Proteins in Acute Lymphoblastic Leukemia: Evidence of Selective Expression of KCTD1 in T-ALL. J. Clin. Med. 2023, 12, 3669. https://doi.org/10.3390/jcm12113669
Buono L, Iside C, Pecoraro G, De Matteo A, Beneduce G, Penta de Vera d’Aragona R, Parasole R, Mirabelli P, Vitagliano L, Salvatore M, et al. A Comprehensive Analysis of the Expression Profiles of KCTD Proteins in Acute Lymphoblastic Leukemia: Evidence of Selective Expression of KCTD1 in T-ALL. Journal of Clinical Medicine. 2023; 12(11):3669. https://doi.org/10.3390/jcm12113669
Chicago/Turabian StyleBuono, Lorena, Concetta Iside, Giovanni Pecoraro, Antonia De Matteo, Giuliana Beneduce, Roberta Penta de Vera d’Aragona, Rosanna Parasole, Peppino Mirabelli, Luigi Vitagliano, Marco Salvatore, and et al. 2023. "A Comprehensive Analysis of the Expression Profiles of KCTD Proteins in Acute Lymphoblastic Leukemia: Evidence of Selective Expression of KCTD1 in T-ALL" Journal of Clinical Medicine 12, no. 11: 3669. https://doi.org/10.3390/jcm12113669