
Ileocolonic Healing after Small Ileocecal Resection in Mice: NOD2 Deficiency Impairs Anastomotic Healing by Local Mechanisms
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animal Monitoring
2.2. Operative Procedure
2.3. Bursting Pressure Measurement
2.4. Tissue Harvest
2.5. Protein Isolation and Hydroxyproline (OHP) Measurement
2.6. RNA Isolation
2.7. Quantitative Real-Time PCR
2.8. Histology
2.9. Naphtol-AS-D-Chloresterase (ASD) Staining
2.10. Quantification of Anastomotic Granulation Tissue
2.11. Isolation of Wound Fibroblasts
2.12. Multiplex Analysis of Plasma Cytokines
2.13. Statistics
3. Results
3.1. Wellness Score, Vital Parameters and Survival
3.2. Anastomotic Healing Measured by Leak Rate, Bursting Pressure and Formation of Granulation Tissue
3.3. mRNA Expression of Proteins Related to Matrix Turnover in Anastomosis
3.4. mRNA Expression of Proteins Related to Matrix Turnover in Wound-Derived Fibroblasts
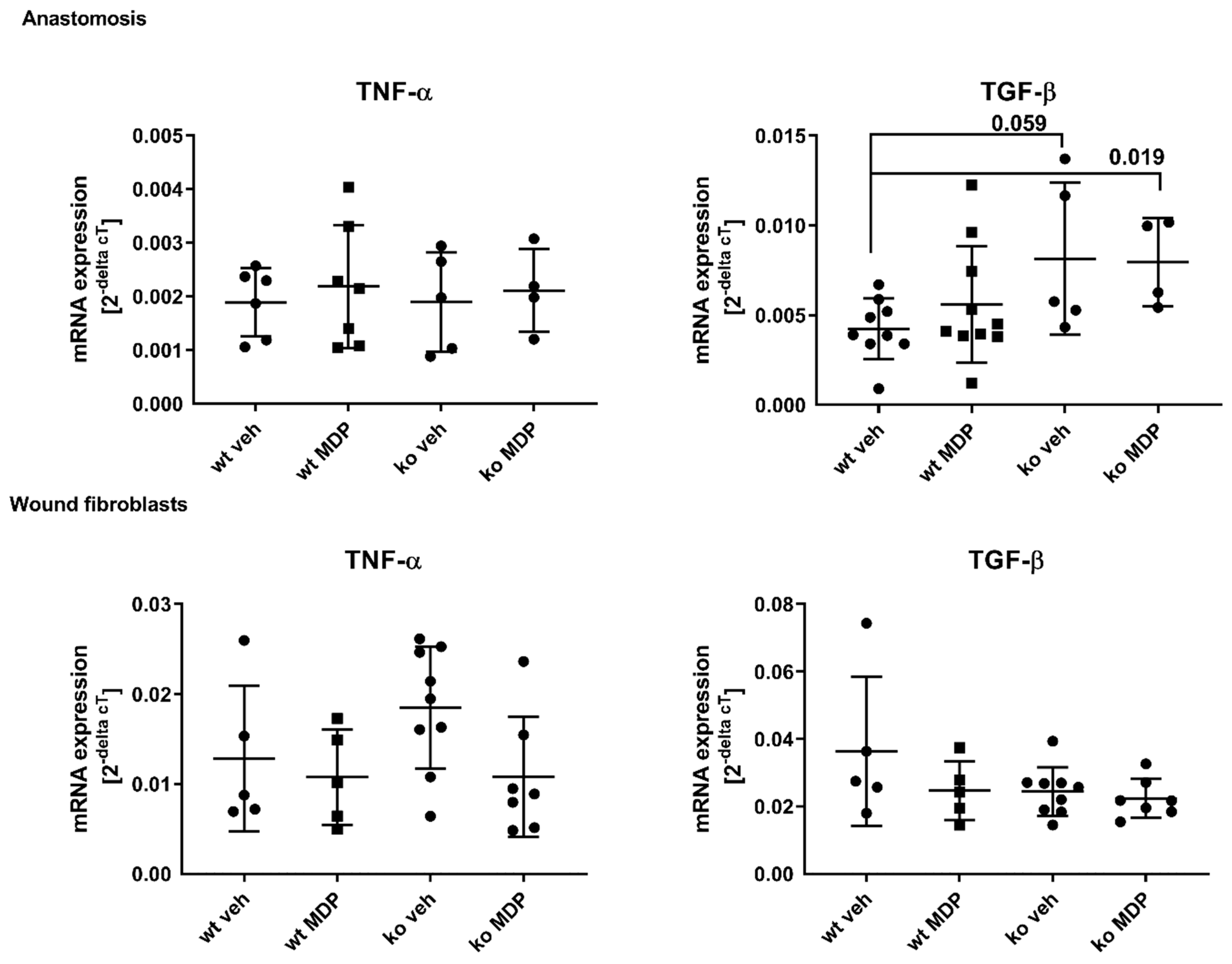
3.5. Cytokine Expression in the Anastomosis and in Wound Derived Fibroblasts
3.6. Plasma Cytokines
4. Discussion
4.1. The Effect of NOD2 Deficiency on Ileocolonic Anastomotic Healing Is Local
4.2. Local Dysbiosis May Be Involved in Impaired Anastomotic Healing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hammond, J.; Lim, S.; Wan, Y.; Gao, X.; Patkar, A. The burden of gastrointestinal anastomotic leaks: An evaluation of clinical and economic outcomes. J. Gastrointest. Surg. 2014, 18, 1176–1185. [Google Scholar] [PubMed]
- Fumery, M.; Seksik, P.; Auzolle, C.; Munoz-Bongrand, N.; Gornet, J.-M.; Boschetti, G.; Cotte, E.; Buisson, A.; Dubois, A.; Pariente, B.; et al. Postoperative Complications after Ileocecal Resection in Crohn’s Disease: A Prospective Study from the REMIND Group. Am. J. Gastroenterol. 2017, 112, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Gershuni, V.M.; Friedman, E.S. The Microbiome-Host Interaction as a Potential Driver of Anastomotic Leak. Curr. Gastroenterol. Rep. 2019, 21, 4. [Google Scholar] [CrossRef] [PubMed]
- Williamson, A.J.; Alverdy, J.C. Influence of the Microbiome on Anastomotic Leak. Clin. Colon Rectal Surg. 2021, 34, 439–446. [Google Scholar] [CrossRef] [PubMed]
- 2017 European Society of Coloproctology (ESCP) Collaborating Group. Association of mechanical bowel preparation with oral antibiotics and anastomotic leak following left sided colorectal resection: An International, Multi-Centre, Prospective Audit. Colorectal Dis. 2018, 6 (Suppl. 6), 15–32. [Google Scholar]
- Shogan, B.D.; Smith, D.P.; Christley, S.; A Gilbert, J.; Zaborina, O.; Alverdy, J.C. Intestinal anastomotic injury alters spatially defined microbiome composition and function. Microbiome 2014, 2, 35. [Google Scholar] [CrossRef]
- Berlin, P.; Barrantes, I.; Reiner, J.; Schröder, E.; Vollmar, B.; Bull, J.; Kreikemeyer, B.; Lamprecht, G.; Witte, M. Dysbiosis and reduced small intestinal function are required to induce intestinal insufficiency in mice. Am. J. Physiology. Gastrointest. Liver Physiol. 2023, 324, G10–G23. [Google Scholar] [CrossRef]
- Shogan, B.D.; Belogortseva, N.; Luong, P.M.; Zaborin, A.; Lax, S.; Bethel, C.; Ward, M.; Muldoon, J.P.; Singer, M.; An, G.; et al. Collagen degradation and mmp9 activation by Enterococcus faecalis contribute to intestinal anastomotic leak. Sci. Transl. Med. 2015, 7, 286ra68. [Google Scholar]
- Ågren, M.S.; Andersen, T.L.; Andersen, L.; Schiødt, C.B.; Surve, V.; Andreassen, T.T.; Risteli, J.; Franzén, L.E.; Delaissé, J.-M.; Heegaard, A.-M.; et al. Nonselective matrix metalloproteinase but not tumor necrosis factor-α inhibition effectively preserves the early critical colon anastomotic integrity. Int. J. Color. Dis. 2011, 26, 329–337. [Google Scholar] [CrossRef]
- Hingh, I.H.J.T.d.; Siemonsma, M.A.; Man, B.M.d.; Lomme, R.M.L.M.; Hendriks, T. The matrix metalloproteinase inhibitor BB-94 improves the strength of intestinal anastomoses in the rat. Int. J. Color. Dis. 2002, 17, 348–354. [Google Scholar] [CrossRef]
- Bedirli, A.; Salman, B.; Pasaoglu, H.; Ofluoglu, E.; Sakrak, O. Effects of nuclear factor-κB inhibitors on colon anastomotic healing in rats. J. Surg. Res. 2011, 171, 355–360. [Google Scholar] [CrossRef]
- Lee, R.H.; Efron, D.T.; Tantry, U.; Stuelten, C.; Moldawer, L.L.; Abouhamze, A.; Barbul, A. Inhibition of tumor necrosis factor-alpha attenuates wound breaking strength in rats. Wound Repair Regen. 2000, 8, 547–553. [Google Scholar] [CrossRef]
- Hugot, J.P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 2001, 411, 599–603. [Google Scholar] [CrossRef]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H.; et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef]
- Schardey, J.; Zehl, S.; Kappenberger, A.S.; Zimmermann, P.; Beigel, F.; Schiergens, T.S.; Kasparek, M.S.; Kühn, F.; Werner, J.; Wirth, U. It is not NOD2–genetic and clinical risk factors for postoperative complications following ileocolic resection in Crohn’s disease. Int. J. Color. Dis. 2022, 37, 1901–1908. [Google Scholar] [CrossRef]
- Kline, B.P.; Weaver, T.; Brinton, D.L.; Deiling, S.; Yochum, G.S.; Berg, A.S.; Koltun, W.A. Clinical and Genetic Factors Associated with Complications After Crohn’s Ileocolectomy. Dis. Colon Rectum 2020, 63, 357–364. [Google Scholar] [CrossRef]
- Witte, M.; Reiner, J.; Bannert, K.; Jaster, R.; Maschmeier, C.; Schafmayer, C.; Lamprecht, G.; Berlin, P. Ileocolonic Healing After Extended Small Bowel Resection in Mice: NOD2 Deficiency Impairs Anastomotic Healing and Postoperative Outcome. Inflamm. Bowel Dis. 2021, 27, 1503–1512. [Google Scholar] [CrossRef]
- Sidiq, T.; Yoshihama, S.; Downs, I.; Kobayashi, K.S. Nod2: A Critical Regulator of Ileal Microbiota and Crohn’s Disease. Front. Immunol. 2016, 7, 367. [Google Scholar] [CrossRef]
- Brown, G.L.; Foshee, H.; Pietsch, J.; Polk, H.C. Muramyl dipeptide enhances survival from experimental peritonitis. Arch. Surg. 1986, 121, 47–49. [Google Scholar] [CrossRef]
- Campbell, L.; Williams, H.; Crompton, R.A.; Cruickshank, S.M.; Hardman, M.J. Nod2 deficiency impairs inflammatory and epithelial aspects of the cutaneous wound-healing response. J. Pathol. 2013, 229, 121–131. [Google Scholar] [CrossRef]
- Williams, H.; Crompton, R.A.; Thomason, H.A.; Campbell, L.; Singh, G.; McBain, A.J.; Cruickshank, S.M.; Hardman, M.J. Cutaneous Nod2 Expression Regulates the Skin Microbiome and Wound Healing in a Murine Model. J. Investig. Dermatol. 2017, 137, 2427–2436. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, S.-M.; Wakenshaw, L.; Cardone, J.; Howdle, P.-D.; Murray, P.-J.; Carding, S.-R. Evidence for the involvement of NOD2 in regulating colonic epithelial cell growth and survival. World J. Gastroenterol. 2008, 14, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Berlin, P.; Reiner, J.; Witte, M.; Wobar, J.; Lindemann, S.; Barrantes, I.; Kreikemeyer, B.; Bastian, M.; Schäffler, H.; Bannert, K.; et al. Nod2 deficiency functionally impairs adaptation to short bowel syndrome via alterations of the epithelial barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G727–G738. [Google Scholar] [CrossRef] [PubMed]
- Komen, N.; van der Wal, H.-C.; Ditzel, M.; Kleinrensink, G.-J.; Jeekel, H.; Lange, J.F. Colorectal anastomotic leakage: A new experimental model. J. Surg. Res. 2009, 155, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wiegerinck, M.; Hyoju, S.K.; Mao, J.; Zaborin, A.; Adriaansens, C.; Salzman, E.; Hyman, N.H.; Zaborina, O.; van Goor, H.; Alverdy, J.C. Novel de novo synthesized phosphate carrier compound ABA-PEG20k-Pi20 suppresses collagenase production in Enterococcus faecalis and prevents colonic anastomotic leak in an experimental model. Br. J. Surg. 2018, 105, 1368–1376. [Google Scholar] [CrossRef]
- Witte, M.B.; Barbul, A.; Schick, M.A.; Vogt, N.; Becker, H.D. Upregulation of arginase expression in wound-derived fibroblasts. J. Surg. Res. 2002, 105, 35–42. [Google Scholar] [CrossRef]
- Lala, S.; Ogura, Y.; Osborne, C.; Hor, S.Y.; Bromfield, A.; Davies, S.; Ogunbiyi, O.; Nuñez, G.; Keshav, S. Crohn’s disease and the NOD2 gene: A role for paneth cells. Gastroenterology 2003, 125, 47–57. [Google Scholar] [CrossRef]
- Pauleau, A.-L.; Murray, P.J. Role of nod2 in the response of macrophages to toll-like receptor agonists. Mol. Cell. Biol. 2003, 23, 7531–7539. [Google Scholar] [CrossRef]
- Wehkamp, J.; Harder, J.; Weichenthal, M.; Schwab, M.; Schäffeler, E.; Schlee, M.; Herrlinger, K.R.; Stallmach, A.; Noack, F.; Fritz, P.; et al. NOD2 (CARD15) mutations in Crohn’s disease are associated with diminished mucosal alpha-defensin expression. Gut 2004, 53, 1658–1664. [Google Scholar] [CrossRef]
- Balasubramanian, I.; Gao, N. From sensing to shaping microbiota: Insights into the role of NOD2 in intestinal homeostasis and progression of Crohn’s disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G7–G13. [Google Scholar] [CrossRef]
- Petnicki-Ocwieja, T.; Hrncir, T.; Liu, Y.-J.; Biswas, A.; Hudcovic, T.; Tlaskalova-Hogenova, H.; Kobayashi, K.S. Nod2 is required for the regulation of commensal microbiota in the intestine. Proc. Natl. Acad. Sci. USA 2009, 106, 15813–15818. [Google Scholar] [CrossRef]
- Brown, K.; Abbott, D.W.; Uwiera, R.R.E.; Inglis, G.D. Removal of the cecum affects intestinal fermentation, enteric bacterial community structure, and acute colitis in mice. Gut Microbes 2018, 9, 218–235. [Google Scholar] [CrossRef]
- Rehman, A.; Sina, C.; Gavrilova, O.; Häsler, R.; Ott, S.; Baines, J.F.; Schreiber, S.; Rosenstiel, P. Nod2 is essential for temporal development of intestinal microbial communities. Gut 2011, 60, 1354–1362. [Google Scholar] [CrossRef]
- Robertson, S.J.; Zhou, J.Y.; Geddes, K.; Rubino, S.J.; Cho, J.H.; Girardin, S.E.; Philpott, D.J. Nod1 and Nod2 signaling does not alter the composition of intestinal bacterial communities at homeostasis. Gut Microbes 2013, 4, 222–231. [Google Scholar] [CrossRef]
- Jang, K.K.; Heaney, T.; London, M.; Ding, Y.; Yeung, F.; Ercelen, D.; Chen, Y.-H.; Axelrad, J.; Gurunathan, S.; Marijke Keestra-Gounder, A.; et al. Antimicrobial overproduction sustains intestinal inflammation by inhibiting Enterococcus colonization. bioRxiv, 2023; preprint. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Shaw, M.H.; Warner, N.; Park, J.-H.; Chen, F.; Ogura, Y.; Núñez, G. Cutting edge: Crohn’s disease-associated Nod2 mutation limits production of proinflammatory cytokines to protect the host from Enterococcus faecalis-induced lethality. J. Immunol. 2011, 187, 2849–2852. [Google Scholar] [CrossRef]
- Hajjar, R.; Gonzalez, E.; Fragoso, G.; Oliero, M.; Alaoui, A.A.; Calvé, A.; Vennin Rendos, H.; Djediai, S.; Cuisiniere, T.; Laplante, P.; et al. Gut microbiota influence anastomotic healing in colorectal cancer surgery through modulation of mucosal proinflammatory cytokines. Gut 2022, 72, 1143–1154. [Google Scholar] [CrossRef]
- Alam, A.; Leoni, G.; Quiros, M.; Wu, H.; Desai, C.; Nishio, H.; Jones, R.M.; Nusrat, A.; Neish, A.S. The microenvironment of injured murine gut elicits a local pro-restitutive microbiota. Nat. Microbiol. 2016, 1, 15021. [Google Scholar] [CrossRef]
- Devine, A.A.; Gonzalez, A.; Speck, K.E.; Knight, R.; Helmrath, M.; Lund, P.K.; Azcarate-Peril, M.A. Impact of ileocecal resection and concomitant antibiotics on the microbiome of the murine jejunum and colon. PloS ONE 2013, 8, e73140. [Google Scholar] [CrossRef]
- Moltzau Anderson, J.; Lipinski, S.; Sommer, F.; Pan, W.-H.; Boulard, O.; Rehman, A.; Falk-Paulsen, M.; Stengel, S.T.; Aden, K.; Häsler, R.; et al. NOD2 Influences Trajectories of Intestinal Microbiota Recovery After Antibiotic Perturbation. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 365–389. [Google Scholar] [CrossRef]
- Vissers, M.; Hartman, Y.; Groh, L.; de Jong, D.J.; de Jonge, M.I.; Ferwerda, G. Recognition of Streptococcus pneumoniae and muramyl dipeptide by NOD2 results in potent induction of mmp9, which can be controlled by lipopolysaccharide stimulation. Infect. Immun. 2014, 82, 4952–4958. [Google Scholar] [CrossRef] [PubMed]
- Sparreboom, C.L.; Komen, N.; Rizopoulos, D.; Verhaar, A.P.; Dik, W.A.; Wu, Z.; van Westreenen, H.L.; Doornebosch, P.G.; Dekker, J.W.T.; Menon, A.G.; et al. A multicentre cohort study of serum and peritoneal biomarkers to predict anastomotic leakage after rectal cancer resection. Color. Dis. 2020, 22, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Komen, N.; Slieker, J.; Willemsen, P.; Mannaerts, G.; Pattyn, P.; Karsten, T.; de Wilt, H.; van der Harst, E.; van Leeuwen, W.; Decaestecker, C.; et al. Polymerase chain reaction for Enterococcus faecalis in drain fluid: The first screening test for symptomatic colorectal anastomotic leakage. The Appeal-study: Analysis of parameters predictive for evident anastomotic leakage. Int. J. Color. Dis. 2014, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Kawamura, T.; Andoh, A.; Nishida, A.; Shioya, M.; Yagi, Y.; Nishimura, T.; Hashimoto, T.; Tsujikawa, T.; Yasui, H.; Fujiyama, Y. Inhibitory effects of short-chain fatty acids on matrix metalloproteinase secretion from human colonic subepithelial myofibroblasts. Dig. Dis. Sci. 2009, 54, 238–245. [Google Scholar] [CrossRef]
- Economou, M.; Trikalinos, T.; Loizou, K.; Tsianos, E.; Ioannidis, J. Differential effects of NOD2 variants on Crohn’s disease risk and phenotype in diverse populations: A metaanalysis. Am. J. Gastroenterol. 2004, 99, 2393–2404. [Google Scholar] [CrossRef]
- Lesage, S.; Zouali, H.; Cezard, J.-P.; Colombel, J.-F.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.; Gassull, M.; Binder, B.; et al. CARD15/NOD2 Mutational Analysis and Genotype-Phenotype Correlation in 612 Patients with Inflammatory Bowel Disease. Am. J. Hum. Genet. 2002, 70, 845–857. [Google Scholar] [CrossRef]
- Maeda, S.; Hsu, L.-C.; Liu, H.; Bankston, L.A.; Limura, M.; Kagnoff, M.F.; Eckmann, L.; Karin, M. Nod2 Mutation in Crohn’s Disease Potentiates NF-kB Activity and IL-1b Processing. Science 2005, 307, 734–738. [Google Scholar] [CrossRef]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nunez, G.; Flavell, R.A. Nod2-Dependent Regulation of Innate and Adaptive Immunity in the Intestinal Tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef]
- Dongliang, L.; Lichuan, L.; Liu, L.; Zhiqiang, Z. Does intraoperative indocyanine green fluorescence angiography decrease the incidence of anastomotic leakage in colorectal surgery? A systematic review and meta-analysis. Int. J. Colorect. Dis. 2021, 36, 57–66. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witte, M.B.; Saupe, J.; Reiner, J.; Bannert, K.; Schafmayer, C.; Lamprecht, G.; Berlin, P. Ileocolonic Healing after Small Ileocecal Resection in Mice: NOD2 Deficiency Impairs Anastomotic Healing by Local Mechanisms. J. Clin. Med. 2023, 12, 3601. https://doi.org/10.3390/jcm12103601
Witte MB, Saupe J, Reiner J, Bannert K, Schafmayer C, Lamprecht G, Berlin P. Ileocolonic Healing after Small Ileocecal Resection in Mice: NOD2 Deficiency Impairs Anastomotic Healing by Local Mechanisms. Journal of Clinical Medicine. 2023; 12(10):3601. https://doi.org/10.3390/jcm12103601
Chicago/Turabian StyleWitte, Maria B., Johannes Saupe, Johannes Reiner, Karen Bannert, Clemens Schafmayer, Georg Lamprecht, and Peggy Berlin. 2023. "Ileocolonic Healing after Small Ileocecal Resection in Mice: NOD2 Deficiency Impairs Anastomotic Healing by Local Mechanisms" Journal of Clinical Medicine 12, no. 10: 3601. https://doi.org/10.3390/jcm12103601