
Alteration of Serum Proteome in Levo-Thyroxine-Euthyroid Thyroidectomized Patients

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients’ Population
2.2. Proteomic and Enrichment Analysis
2.3. Bidimensional Western Blot Validation of A1AT and APOAI
3. Results
3.1. Comparison of Post-Surgical Thyroid Hormone Levels between “Reduced” and “Stable” FT3 Patients
3.2. Proteomic Analysis Results
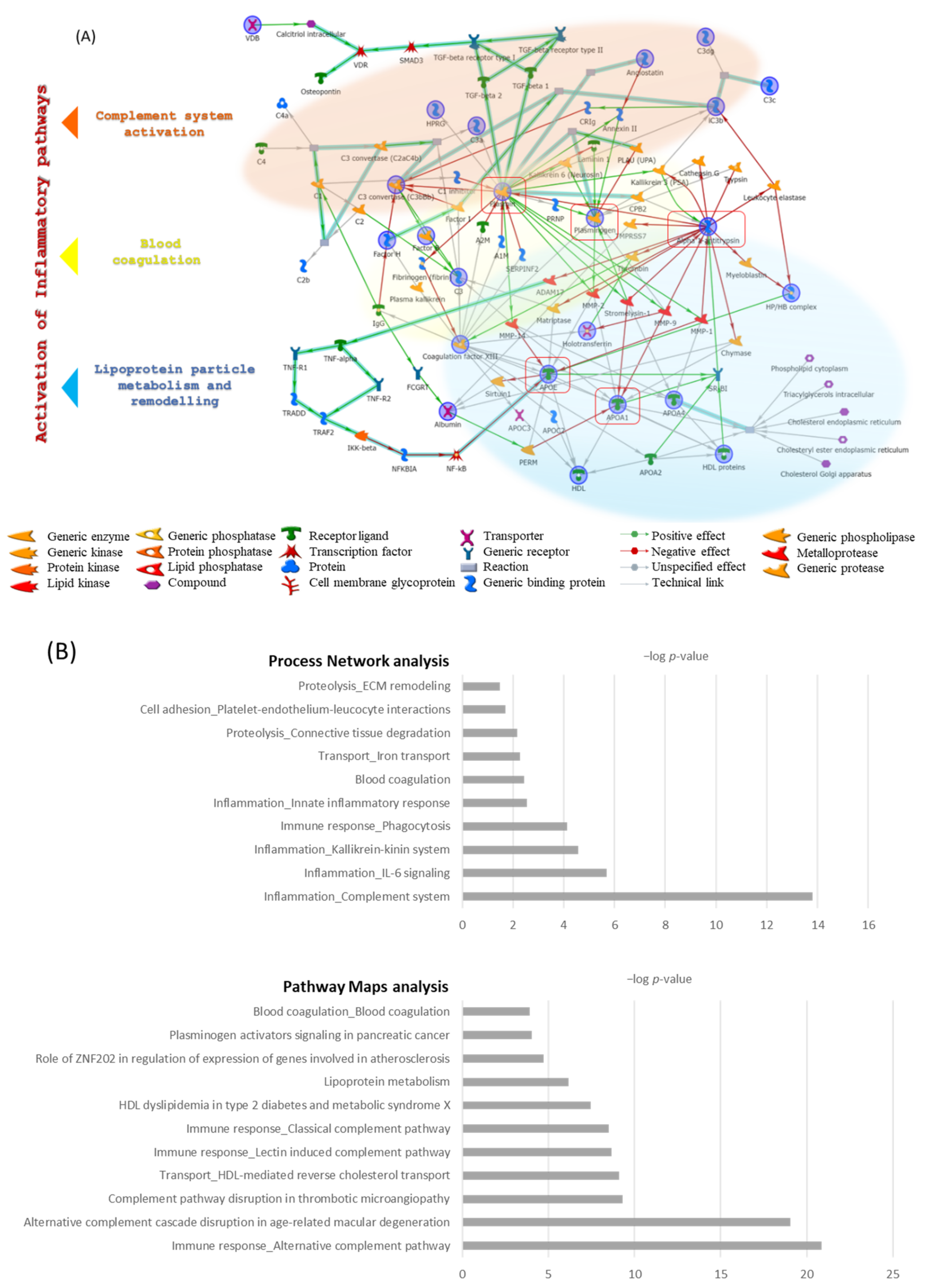
3.3. Enrichment Analysis Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garber, J.R.; Cobin, R.H.; Gharib, H.; Hennessey, J.V.; Klein, I.; Mechanick, J.I.; Pessah-Pollack, R.; Singer, P.A.; Woeber, K.A. American Association of Clinical Endocrinologists and American Thyroid Association Taskforce on Hypothyroidism in Adults Clinical Practice Guidelines for Hypothyroidism in Adults: Cosponsored by the American Association of Clinical Endocrinologists and the American Thyroid Association. Endocr. Pract. 2012, 18, 988–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- e Drigo, R.A.; Fonseca, T.L.; Werneck-de-Castro, J.P.S.; Bianco, A.C. Role of the Type 2 Iodothyronine Deiodinase (D2) in the Control of Thyroid Hormone Signaling. Biochim. Biophys. Acta 2013, 1830, 3956–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, P.; Chau, W.-F.; Roberts, N.; Vedhara, K.; Greenwood, R.; Dayan, C.M. Psychological Well-Being in Patients on “adequate” Doses of l-Thyroxine: Results of a Large, Controlled Community-Based Questionnaire Study. Clin. Endocrinol. 2002, 57, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.M.; Duntas, L.; Fadeyev, V.; Nygaard, B.; Vanderpump, M.P.J. 2012 ETA Guidelines: The Use of L-T4 + L-T3 in the Treatment of Hypothyroidism. Eur. Thyroid J. 2012, 1, 55–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagna, M.G.; Dentice, M.; Cantara, S.; Ambrosio, R.; Maino, F.; Porcelli, T.; Marzocchi, C.; Garbi, C.; Pacini, F.; Salvatore, D. DIO2 Thr92Ala Reduces Deiodinase-2 Activity and Serum-T3 Levels in Thyroid-Deficient Patients. J. Clin. Endocrinol. Metab. 2017, 102, 1623–1630. [Google Scholar] [CrossRef]
- Pagni, F.; L’Imperio, V.; Bono, F.; Garancini, M.; Roversi, G.; De Sio, G.; Galli, M.; Smith, A.J.; Chinello, C.; Magni, F. Proteome Analysis in Thyroid Pathology. Expert Rev. Proteom. 2015, 12, 375–390. [Google Scholar] [CrossRef]
- Engelmann, B.; Bischof, J.; Dirk, A.-L.; Friedrich, N.; Hammer, E.; Thiele, T.; Führer, D.; Homuth, G.; Brabant, G.; Völker, U. Effect of Experimental Thyrotoxicosis onto Blood Coagulation: A Proteomics Study. Eur. Thyroid J. 2015, 4, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Pietzner, M.; Engelmann, B.; Kacprowski, T.; Golchert, J.; Dirk, A.L.; Hammer, E.; Iwen, K.A.; Nauck, M.; Wallaschofski, H.; Führer, D.; et al. Plasma Proteome and Metabolome Characterization of an Experimental Human Thyrotoxicosis Model. Available online: https://pubmed.ncbi.nlm.nih.gov/28065164/ (accessed on 2 December 2020).
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Jammah, A.A.; Ekhzaimy, A.A. Differences in the Plasma Proteome of Patients with Hypothyroidism before and after Thyroid Hormone Replacement: A Proteomic Analysis. Int. J. Mol. Sci. 2018, 19, 88. [Google Scholar] [CrossRef] [Green Version]
- Landi, C.; Bargagli, E.; Magi, B.; Prasse, A.; Muller-Quernheim, J.; Bini, L.; Rottoli, P. Proteome Analysis of Bronchoalveolar Lavage in Pulmonary Langerhans Cell Histiocytosis. J. Clin. Bioinform. 2011, 1, 31. [Google Scholar] [CrossRef] [Green Version]
- Sulpizio, M.; Falone, S.; Amicarelli, F.; Marchisio, M.; Di Giuseppe, F.; Eleuterio, E.; Di Ilio, C.; Angelucci, S. Molecular Basis Underlying the Biological Effects Elicited by Extremely Low-Frequency Magnetic Field (ELF-MF) on Neuroblastoma Cells. J. Cell. Biochem. 2011, 112, 3797–3806. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE Database and Related Tools and Resources in 2019: Improving Support for Quantification Data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Ozbey, O.; Zingel, O. An Immunoblotting Method for High-Resolution Isoelectric Focusing of Protein Isoforms on Immobilized PH Gradients. Electrophoresis 2001, 22, 1887–1893. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic Transfer of Proteins from Polyacrylamide Gels to Nitrocellulose Sheets: Procedure and Some Applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Cantara, S.; Ricci, C.; Maino, F.; Marzocchi, C.; Pacini, F.; Castagna, M.G. Variants in MCT10 Protein Do Not Affect FT3 Levels in Athyreotic Patients. Endocrine 2019, 66, 551–556. [Google Scholar] [CrossRef]
- Bitencourt, C.S.; Duarte, C.G.; Azzolini, A.E.C.S.; Assis-Pandochi, A.I. Alternative Complement Pathway and Factor B Activities in Rats with Altered Blood Levels of Thyroid Hormone. Braz. J. Med. Biol. Res. 2012, 45, 216–221. [Google Scholar] [CrossRef] [Green Version]
- Afshar-Kharghan, V. The Role of the Complement System in Cancer. J. Clin. Investig. 2017, 127, 780–789. [Google Scholar] [CrossRef] [Green Version]
- Barrington, R.; Zhang, M.; Fischer, M.; Carroll, M.C. The Role of Complement in Inflammation and Adaptive Immunity. Immunol. Rev. 2001, 180, 5–15. [Google Scholar] [CrossRef]
- Lopatko Fagerström, I.; Ståhl, A.; Mossberg, M.; Tati, R.; Kristoffersson, A.-C.; Kahn, R.; Bascands, J.-L.; Klein, J.; Schanstra, J.P.; Segelmark, M.; et al. Blockade of the Kallikrein-Kinin System Reduces Endothelial Complement Activation in Vascular Inflammation. eBioMedicine 2019, 47, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Ekdahl, K.N.; Teramura, Y.; Hamad, O.A.; Asif, S.; Duehrkop, C.; Fromell, K.; Gustafson, E.; Hong, J.; Kozarcanin, H.; Magnusson, P.U.; et al. Dangerous Liaisons: Complement, Coagulation, and Kallikrein/Kinin Cross-Talk Act as a Linchpin in the Events Leading to Thromboinflammation. Immunol. Rev. 2016, 274, 245–269. [Google Scholar] [CrossRef]
- Türemen, E.E.; Çetinarslan, B.; Şahin, T.; Cantürk, Z.; Tarkun, İ. Endothelial Dysfunction and Low Grade Chronic Inflammation in Subclinical Hypothyroidism Due to Autoimmune Thyroiditis. Endocr. J. 2011, 58, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Himbert, C.; Ose, J.; Lin, T.; Warby, C.A.; Gigic, B.; Steindorf, K.; Schrotz-King, P.; Abbenhardt-Martin, C.; Zielske, L.; Boehm, J.; et al. Inflammation- and Angiogenesis-Related Biomarkers Are Correlated with Cancer-Related Fatigue in Colorectal Cancer Patients: Results from the ColoCare Study. Eur. J. Cancer Care 2019, 28, e13055. [Google Scholar] [CrossRef]
- Herlofson, K.; Heijnen, C.J.; Lange, J.; Alves, G.; Tysnes, O.-B.; Friedman, J.H.; Fagundes, C.P. Inflammation and Fatigue in Early, Untreated Parkinson’s Disease. Acta Neurol. Scand. 2018, 138, 394–399. [Google Scholar] [CrossRef]
- Bellei, E.; Rustichelli, C.; Bergamini, S.; Monari, E.; Baraldi, C.; Lo Castro, F.; Tomasi, A.; Ferrari, A. Proteomic Serum Profile in Menstrual-Related and Post Menopause Migraine. J. Pharm. Biomed. Anal. 2020, 184, 113165. [Google Scholar] [CrossRef]
- Landi, C.; Santinelli, L.; Bianchi, L.; Shaba, E.; Ceccarelli, G.; Cavallari, E.N.; Borrazzo, C.; Pinacchio, C.; Scagnolari, C.; Vullo, V.; et al. Cognitive Impairment and CSF Proteome Modification after Oral Bacteriotherapy in HIV Patients. J. Neurovirol. 2020, 26, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Halaris, A. Inflammation-Associated Co-Morbidity Between Depression and Cardiovascular Disease. Curr. Top. Behav. Neurosci. 2017, 31, 45–70. [Google Scholar] [CrossRef]
- Chadarevian, R.; Bruckert, E.; Leenhardt, L.; Giral, P.; Ankri, A.; Turpin, G. Components of the Fibrinolytic System Are Differently Altered in Moderate and Severe Hypothyroidism. J. Clin. Endocrinol. Metab. 2001, 86, 732–737. [Google Scholar] [CrossRef]
- Jung, K.Y.; Ahn, H.Y.; Han, S.K.; Park, Y.J.; Cho, B.Y.; Moon, M.K. Association between Thyroid Function and Lipid Profiles, Apolipoproteins, and High-Density Lipoprotein Function. J. Clin. Lipidol. 2017, 11, 1347–1353. [Google Scholar] [CrossRef]
- O’Brien, T.; Katz, K.; Hodge, D.; Nguyen, T.T.; Kottke, B.A.; Hay, I.D. The Effect of the Treatment of Hypothyroidism and Hyperthyroidism on Plasma Lipids and Apolipoproteins AI, AII and E. Clin. Endocrinol. 1997, 46, 17–20. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, G.; Pang, H.; Liu, Q.; Liu, Y. The Effect of 131I-Induced Hypothyroidism on the Levels of Nitric Oxide (NO), Interleukin 6 (IL-6), Tumor Necrosis Factor Alpha (TNF-α), Total Nitric Oxide Synthase (NOS) Activity, and Expression of NOS Isoforms in Rats. Bosn. J. Basic Med. Sci. 2018, 18, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Di Segni, C.; Raimondo, S.; Olivieri, G.; Silvestrini, A.; Meucci, E.; Currò, D. Thyroid Hormones, Oxidative Stress, and Inflammation. Mediat. Inflamm. 2016, 2016, 6757154. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reduced FT3 | Stable FT3 | p-Value | |
|---|---|---|---|
| Age (years) | 0.2 | ||
| Mean ± SD | 46 ± 17.2 | 54 ± 7.4 | |
| Range | 24–73 | 44–65 | |
| Median | 44.5 | 54 | |
| Sex | >0.9 | ||
| Males (n, %) | 4 (50%) | 4 (50%) | |
| Females (n, %) | 4(50%) | 4 (50%) | |
| LT4 daily dose (mcg) | 0.6 | ||
| Mean ± SD | 131.2 ± 28.4 | 125 ± 31.3 | |
| Range | 82–175 | 87.5–175 | |
| Median | 131.2 | 118.7 | |
| Pro-Kg LT4 dose (mcg) | 0.01 | ||
| Mean ± SD | 1.8 ± 0.3 | 1.5 ± 0.1 | |
| Range | 1.4–2.3 | 1.3–1.6 | |
| Median | 1.7 | 1.5 | |
| Post-surgical FT3 (pg/mL) | 0.02 | ||
| Mean ± SD | 2.8 ± 0.1 | 3.2 ± 0.3 | |
| Range | 2.6–3.2 | 2.7–3.6 | |
| Median | 2.9 | 3.2 | |
| Normal range: 2.5–4.5 | |||
| Post-surgical FT4 (pg/mL) | 0.4 | ||
| Mean ± SD | 11.2 ± 1.6 | 10.6 ± 1.3 | |
| Range | 8.3–13.6 | 8.5–12.3 | |
| Median | 11.6 | 10.4 | |
| Normal range: 5.8–16.4 | |||
| Post-surgical TSH (mUI/L) | 0.1 | ||
| Mean ± SD | |||
| Range | 1 ± 0.5 | 0.6 ± 0.3 | |
| Median | 0.4–1.7 | 0.4–1.2 | |
| Normal range: 0.4–4.0 | 0.98 | 0.6 | |
| Post-surgical FT3/FT4 | 0.06 | ||
| Mean ± SD | 0.2 ± 0.04 | 0.3 ± 0.04 | |
| Range | 0.19–0.33 | 0.2–0.3 | |
| Median | 0.27 | 0.29 |
| Pre-Surgical | Post-Surgical | p-Value | ||
|---|---|---|---|---|
| “Stable FT3” | TSH (mUI/L) | 0.6 | ||
| Mean ± SD | 0.7 ± 0.3 | 0.6 ± 0.3 | ||
| Range | 0.4–1.6 | 0.4–1.2 | ||
| Median | 0.7 | 0.6 | ||
| FT3 pg/mL | 0.4 | |||
| Mean ± SD | 3.3 ± 0.3 | 3.2 ± 0.3 | ||
| Range | 2.9–3.9 | 2.7–3.6 | ||
| Median | 3.3 | 3.2 | ||
| FT4 pg/mL | 0.06 | |||
| Mean ± SD | 8.9 ± 1.1 | 10.6 ± 1.3 | ||
| Range | 7.3–10.7 | 8.5–12.3 | ||
| Median | 8.6 | 10.4 | ||
| “Reduced FT3” | TSH (mUI/L) | 0.4 | ||
| Mean ± SD | 0.9 ± 0.4 | 1 ± 0.5 | ||
| Range | 0.3–1.4 | 0.4–1.7 | ||
| Median | 0.8 | 0.98 | ||
| FT3 pg/mL | <0.001 | |||
| Mean ± SD | 3.7 ± 0.2 | 2.8 ± 0.1 | ||
| Range | 3.4–4.2 | 2.6–3.2 | ||
| Median | 3.7 | 2.9 | ||
| FT4 pg/mL | 0.9 | |||
| Mean ± SD | 11.1 ± 3.3 | 11.2 ± 1.6 | ||
| Range | 7.7–17.7 | 8.3–13.6 | ||
| Median | 9.8 | 11.6 |
| Spot n | Protein Name | UniProt Entry (HUMAN) | AC | Theoretical pI/Mr (kDa) | Mascot Search Results in Swissprot db | Means | Kruscal Wallis p-Value | Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Score | Matched Peptides | Coverage | Ft3red | Ft3 | CTRL | Red/Stable | Red/CTRL | Stable/CTRL | ||||||
| Stable | ||||||||||||||
| 2 | Complement Factor H | CFAH | P08603 | 6.21/143,680 | 158 | 17 | 15% | 0.0254 | 0.014 | 0.050 | 0.012 | 1.836 | −1.981 | −3.639 |
| 4 | Plasminogen | PLMN | P00747 | 7.04/93,247 | 231 | 23 | 31% | 0.0095 | 0.015 | 0.002 | 0.022 | −1.61 | 5.22 | 8.427 |
| 5 | Plasminogen | PLMN | P00747 | 7.04/93,247 | 264 | 22 | 28% | 0.0135 | 0.026 | 0.005 | 0.004 | −1.97 | 2.736 | 5.389 |
| 6 | Complement factor B | CFAB | P00751 | 6.67/86,847 | 83 | 7 | 13% | 0.0592 | 0.052 | 0.015 | 0.01 | 1.141 | 4.001 | 3.508 |
| 7 | Complement factor B | CFAB | P00751 | 6.67/86,847 | 233 | 18 | 29% | 0.0931 | 0.059 | 0.031 | 0.008 | 1.575 | 2.974 | 1.888 |
| 9 | Serotransferrin | TRFE | P02787 | 6.81/79,294 | 96 | 9 | 12% | 0.0297 | 0.025 | 0.007 | 0.028 | 1.181 | 4.378 | 3.706 |
| 10 | Albumin + Coagulation factor XIII B chain | ALBU F13B | P02768 P05160 | 5.92/71,317 6.01/77,742 | 236 = 146 + 111 | 12 15 | 22% 22% | 0.0244 | 0.007 | 0.002 | 0.033 | 3.437 | 12.337 | 3.589 |
| 11 | Albumin + alpha-1B-glycoprotein | ALBU A1BG | P02768 P04217 | 5.92/71,317 5.56/54,790 | 269 = 163 + 133 | 19 15 | 34% 42% | 0.0706 | 0.066 | 0.147 | 0.024 | 1.072 | −2.09 | −2.24 |
| 15 | Complement factor B frag. C-term | CFAB | P00751 | 6.67/86,847 | 104 | 11 | 17% | 0.0215 | 0.041 | 0.002 | 0.0001 | −1.91 | 11.445 | 21.831 |
| 16 | Vitamin D-binding protein + Alpha-1-antitrypsin | VTDB A1AT | P02774 P01009 | 5.32/54,480 5.37/46,878 | 375 = 230 + 158 | 21 17 | 43% 39% | 0.0506 | 0.028 | 0.083 | 0.04 | 1.822 | −1.634 | −2.978 |
| 17 | Alpha-1-antitrypsin + Apolipoprotein A-I | A1AT APOA1 | P01009 P02647 | 5.37/46,878 5.56/30,759 | 437 = 295 + 150 | 29 15 | 70% 49% | 0.1443 | 0.236 | 0.112 | 0.01 | −1.63 | 1.292 | 2.111 |
| 19 | Albumin fragment N-term | ALBU | P02768 | 5.92/71,317 | 244 | 23 | 38% | 0.0445 | 0.044 | 0.022 | 0.036 | 1.013 | 2.013 | 1.986 |
| 20 | Apolipoprotein A-IV + Haptoglobin | APOA4 HPT | P06727 P00738 | 5.528/45,344 6.13/45,861 | 443 = 352 + 116 | 31 14 | 58% 29% | 0.0755 | 0.035 | 0.206 | 0.003 | 2.131 | −2.73 | −5.82 |
| 21 | Albumin fragment C-term + Haptoglobin | ALBU HPT | P02768 P00738 | 5.92/71,317 6.13/45,861 | 235 = 139 + 119 | 16 14 | 27% 36% | 0.0496 | 0.037 | 0.016 | 0.007 | 1.327 | 3.194 | 2.408 |
| 22 | Histidine-rich glycoprotein | HRG | P04196 | 7.09/60,510 | 179 | 14 | 26% | 0.004 | 0.021 | 0.007 | 0.03 | −5.17 | −1.788 | 2.893 |
| 23 | Complement C3 fragment C-term + Histidine-rich glycoprotein | CO3 HRG | P01024 P04196 | 6.02/188,569 7.09/60,510 | 280 = 166 + 114 | 28 14 | 18% 29% | 0.056 | 0.049 | 0.140 | 0.007 | 1.152 | −2.501 | −2.88 |
| 26 | Albumin fragment N-term | ALBU | P02768 | 5.92/71,317 | 253 | 20 | 26% | 0.047 | 0.051 | 0.0033 | 0.002 | 0.924 | 14.432 | 15.622 |
| 30 | Complement C1q subcomponent subunit B | C1QB | P02746 | 8.83/26,933 | 157 | 12 | 44% | 0.0114 | 0.007 | 0.005 | 0.03 | 1.676 | 2.283 | 1.363 |
| 34 | Apolipoprotein A-I | APOA1 | P02647 | 5.56/30,759 | 334 | 28 | 70% | 0.0832 | 0.031 | 0.116 | 0.02 | 2.674 | 0.717 | −3.728 |
| 35 | Apolipoprotein A-I | APOA1 | P02647 | 5.56/30,759 | 321 | 26 | 70% | 0.0964 | 0.025 | 0.1020 | 0.05 | 3.851 | 0.944 | −4.079 |
| 37 | Hemoglobin | HBA | P69905 | 8.72/15,305 | 128 | 7 | 47% | 0.0985 | 0.04 | 0.044 | 0.04 | 2.757 | 2.257 | 0.819 |
| 39 | Apolipoprotein A-I fragment C-term | APOA1 | P02647 | 5.56/30,759 | 100 | 10 | 34% | 0.0141 | 0.04 | 0.0019 | 0.03 | −2.83 | 7.398 | 20.97 |
| 40 | Complement C1s subcomponent | C1S | P09871 | 4.86/78,174 | 250 | 25 | 32% | 0.0067 | 0 | 0.0188 | 0.02 | 0.007 | −2.798 | 0/0.018 |
| 41 | Complement C1s subcomponent | C1S | P09871 | 4.86/78,174 | 91 | 7 | 16% | 0.0112 | 0.012 | 0.0302 | 0.03 | 0.946 | −2.701 | −2.556 |
| 42 | Complement C1s subcomponent | C1S | P09871 | 4.86/78,174 | 150 | 14 | 28% | 0.007 | 0.001 | 0.009 | 0.02 | 7.793 | 0.746 | −10.447 |
| 43 | Apolipoprotein A-I + Alpha-1-antitrypsin | APOA1 A1AT | P02647 P01009 | 5.56/30,759 5.37/46,878 | 172 = 109 + 96 | 10 10 | 35% 23% | 0.0172 | 0.083 | 0.005 | 0.03 | −4.86 | 3.156 | 15.324 |
| 44 | Apolipoprotein A-I fragment N-term | APOA1 | P02647 | 5.56/30,759 | 199 | 16 | 41% | 0.0127 | 0.019 | 0.0914 | 0.01 | 0.661 | −7.204 | −4.759 |
| 45 | Alpha-1-antitrypsin + Apolipoprotein A-I | A1AT APOA1 | P01009 P02647 | 5.37/46,878 5.56/30,759 | 264 = 165 + 112 | 16 11 | 34% 38% | 0.7654 | 0.306 | 0.777 | 0.009 | 2.502 | 0.986 | −2.539 |
| 47 | Apolipoprotein E | APOE | P02649 | 5.44/122,983 | 325 | 26 | 67% | 0.2241 | 0.189 | 0.081 | 0.03 | 1.185 | 2.777 | 2.343 |
| 50 | Apolipoprotein A-I + Alpha-1-antitrypsin | APOA1 A1AT | P02647 P01009 | 5.56/30,759 5.37/46,878 | 329 = 202 + 123 | 19 14 | 58% 34% | 0.6651 | 0.367 | 1.157 | 0.02 | 1.812 | −1.74 | −3.154 |
| 51 | Complement Factor H | CFAH | P08603 | 6.21/143,680 | 269 | 25 | 23% | 0.0383 | 0.026 | 0.047 | 0.03 | 1.495 | 0.824 | −1.814 |
| 55 | Alpha-1B-glycoprotein | A1BG | P04217 | 5.56/54,790 | 207 | 16 | 36% | 0.0756 | 0.109 | 0.137 | 0.023 | 0.692 | −1.81 | 0.796 |
| 56 | Albumin fragment C-term | ALBU | P02768 | 5.92/71,317 | 253 | 22 | 37% | 0.0819 | 0.071 | 0.040 | 0.012 | 1.16 | 2.033 | 1.75 |
| 57 | Immunoglobulin heavy constant gamma 2 | IGHG2 | P01859 | 7.66/36,505 | 54 | 16 | 46% | 0.082 | 0.074 | 0.049 | 0.03 | 1.09 | 1.666 | 1.519 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landi, C.; Cantara, S.; Shaba, E.; Vantaggiato, L.; Marzocchi, C.; Maino, F.; Bombardieri, A.; Carleo, A.; Di Giuseppe, F.; Angelucci, S.; et al. Alteration of Serum Proteome in Levo-Thyroxine-Euthyroid Thyroidectomized Patients. J. Clin. Med. 2022, 11, 1676. https://doi.org/10.3390/jcm11061676
Landi C, Cantara S, Shaba E, Vantaggiato L, Marzocchi C, Maino F, Bombardieri A, Carleo A, Di Giuseppe F, Angelucci S, et al. Alteration of Serum Proteome in Levo-Thyroxine-Euthyroid Thyroidectomized Patients. Journal of Clinical Medicine. 2022; 11(6):1676. https://doi.org/10.3390/jcm11061676
Chicago/Turabian StyleLandi, Claudia, Silvia Cantara, Enxhi Shaba, Lorenza Vantaggiato, Carlotta Marzocchi, Fabio Maino, Alessio Bombardieri, Alfonso Carleo, Fabrizio Di Giuseppe, Stefania Angelucci, and et al. 2022. "Alteration of Serum Proteome in Levo-Thyroxine-Euthyroid Thyroidectomized Patients" Journal of Clinical Medicine 11, no. 6: 1676. https://doi.org/10.3390/jcm11061676