
Why Should Constant Stimulation of Saccular Afferents Modify the Posture and Gait of Patients with Bilateral Vestibular Dysfunction? The Saccular Substitution Hypothesis


{kind=link}
Abstract
:1. Introduction
- Tests of peripheral vestibular function,
- Tests of postural stability and gait performance, and
- Patient subjective experience. Do they find constant ES to be an improvement?
2. The Steps in the Research Process
- Selection. Patients are selected according to carefully defined selection criteria defining bilateral vestibular dysfunction (BVD). It should be stressed that there is some residual peripheral vestibular function in these patients, but their level of function falls below the international accepted standard for bilateral vestibular dysfunction (Barany Society consensus statement [2]);
- Preoperative testing of peripheral vestibular function, postural stability and gait (e.g., using the DGI and CDP, etc.);
- The electrode and implantation. A modified cochlear implant electrode is implanted on a branch of the vestibular nerve in one ear. The exact location of the electrode depends on particular anatomical considerations at surgery. The aim is to implant the electrode very close to the inferior vestibular nerve containing afferents from the saccular macula and posterior semicircular canal. From the CT verified electrode locations, it appears that the electrode locations so far have been in the inferior vestibular nerve and so largely otolithic, mainly saccular afferents. It is likely there is some stimulation of the afferents from the posterior canal; however, it is necessary to emphasise that this is uncertain. We will refer to the data as being due to saccular stimulation, whereas in fact, it more likely should be referred to as vestibular stimulation (probably mainly saccular);
- Post-operative testing. Initially, it is necessary to verify that the stimulating electrode is functional and make adjustments of the stimulus parameters of the ES (relying on patient subjective reports of oscillopsia or subjective sensations);
- The stimulus. Then, the ES is switched on a constant train of pulses at high-frequency (900 or 1200 pulses/s), and it is kept on continuously. It is important to stress that with this implant, there is, unlike other vestibular implants [3], no modulation of the pulse rate or pulse amplitude in response to the movement of the patient. The ES is composed of a constant pulse train delivered by three electrodes of a modified cochlear implant electrode. Each pulse is biphasic (25 microseconds per phase) and delivered at a frequency of 900 or 1200 pulses per second. Three electrodes are activated, so the stimulus is simply a constant train of pulses, which will activate vestibular afferents at a very high rate, presumably duplicating the constant barrage of action potentials that healthy patients receive;
- Stimulation phase. The patient is then re-tested on repeated occasions in the postoperative phase (in some patients, this phase has lasted 1.5 years so far). On each testing occasion, this is done first to ensure the electrode is working. Fibrosis or changes in impedance may make the electrode non-functional, in which case vestibular and clinical tests would be rendered meaningless. Then, on each occasion, they are tested on the battery of tests of peripheral vestibular function, postural stability, gait performance, and acceptability. Their results on these tests are then compared to their preoperative measures and their performance in comparison to the electrode OFF condition and measures of patient satisfaction (usually relying on patient subjective reports or oscillopsia).
3. Preliminary Test Results
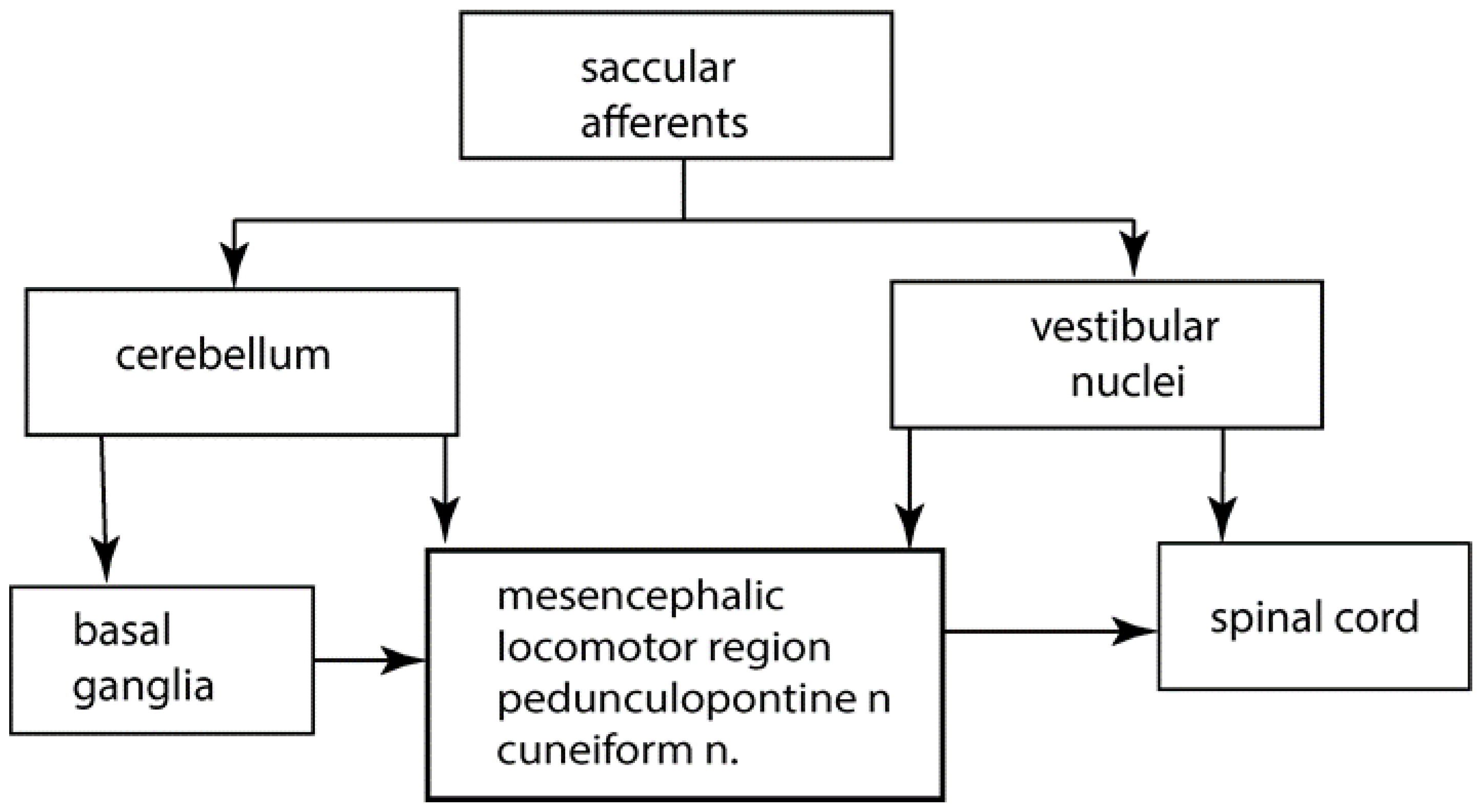
4. The Saccular Substitution Hypothesis
- Where do saccular afferents project to? and
- How could their sustained activation influence posture and gait performance?
5. Saccular Projections
6. The Role of the Cerebellum in Posture and Gait
7. Cerebellar Locomotor Region
8. Cerebellum to Basal Ganglia
9. The Pedunculopontine Nucleus (PPN)
10. Vestibular Nuclei to Basal Ganglia
11. Gait
12. Contribution of the Otoliths to Spatial Awareness
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramos Macias, A.; Ramos de Miguel, A.; Montesdeoca, I.R.; Barreiro, S.B.; Carlos, F.G.J. Chronic electrical stimulation of the otolith organ: Preliminary results in humans with bilateral vestibulopathy and sensorineural hearing loss. Audiol. Neurotol. 2020, 25, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Strupp, M.; Kim, J.S.; Murofushi, T.; Straumann, D.; Jen, J.C.; Rosengren, S.M.; Della Santina, C.C.; Kingma, H. Bilateral vestibulopathy: Diagnostic criteria Consensus document of the Classification Committee of the Barany Society. J. Vestib. Res.-Equilib. Orientat. 2017, 27, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Santina, C.C.; Migliaccio, A.A.; Patel, A.H. A multichannel semicircular canal neural prosthesis using electrical stimulation to restore 3-D vestibular sensation. IEEE Trans. Biomed. Eng. 2007, 54, 1016–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttner-Ennever, J.A. A review of otolith pathways to brainstem and cerebellum. In Otolith Function in Spatial Orientation and Movement; Cohen, B., Hess, B.J.M., Eds.; The New York Academy of Science: New York, NY, USA, 1999; Volume 871, pp. 51–64. [Google Scholar]
- Won, S.L.; Suarez, C.; Honrubia, V.; Gomez, J. Morphological aspects of the human vestibular nerve. Laryngoscope 1990, 100, 756–764. [Google Scholar]
- Goldberg, J.M. Afferent diversity and the organization of central vestibular pathways. Exp. Brain Res. 2000, 130, 277–297. [Google Scholar] [CrossRef] [PubMed]
- Maklad, A.; Fritzsch, B. Partial segregation of posterior crista and saccular fibers to the nodulus and uvula of the cerebellum in mice, and its development. Dev. Brain Res. 2003, 140, 223–236. [Google Scholar] [CrossRef]
- Maklad, A.; Kamel, S.; Wong, E.; Fritzsch, B. Development and organization of polarity-specific segregation of primary vestibular afferent fibers in mice. Cell Tissue Res. 2010, 340, 303–321. [Google Scholar] [CrossRef] [Green Version]
- Gacek, R.R. The course and central termination of first order neurons supplying vestibular endorgans in the cat. Acta Otolaryngol. Suppl. 1969, 254, 1–66. [Google Scholar]
- Barmack, N.H.; Baughman, R.W.; Errico, P.; Shojaku, H. Vestibular primary afferent projection to the cerebellum of the rabbit. J. Comp. Neurol. 1993, 327, 521–534. [Google Scholar] [CrossRef]
- Morton, S.M.; Bastian, A.J. Cerebellar control of balance and locomotion. Neuroscientist 2004, 10, 247–259. [Google Scholar] [CrossRef]
- Dijkstra, B.W.; Bekkers, E.M.J.; Gilat, M.; de Rond, V.; Hardwick, R.M.; Nieuwboer, A. Functional neuroimaging of human postural control: A systematic review with meta-analysis. Neurosci. Biobehav. Rev. 2020, 115, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Matsui, T.; Kuze, B.; Asanome, M.; Nakajima, K.; Matsuyama, K. Stimulation of a restricted region in the midline cerebellar white matter evokes coordinated quadrupedal locomotion in the decerebrate cat. J. Neurophysiol. 1999, 82, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Mauritz, K.H.; Dichgans, J.; Hufschmidt, A. Quantitative-analysis of stance in late cortical cerebellar atrophy of the anterior lobe and other forms of cerebellar-ataxia. Brain 1979, 102, 461–482. [Google Scholar] [CrossRef]
- Horak, F.B.; Diener, H.C. Cerebellar control of postural scaling and central set in stance. J. Neurophysiol. 1994, 72, 479–493. [Google Scholar] [CrossRef]
- Timmann, D.; Horak, F.B. Perturbed step initiation in cerebellar subjects: 2. Modification of anticipatory postural adjustments. Exp. Brain Res. 2001, 141, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Bent, L.R.; Inglis, J.T.; McFadyen, B.J. Vestibular contributions across the execution of a voluntary forward step. Exp. Brain Res. 2002, 143, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Bent, L.R.; McFadyen, B.J.; Inglis, J.T. Vestibular contributions during human locomotor tasks. Exerc. Sport Sci. Rev. 2005, 33, 107–113. [Google Scholar] [CrossRef]
- Jahn, K.; Deutschlader, A.; Stephan, T.; Kalla, R.; Wiesmann, M.; Strupp, M.; Brandt, T. Imaging human supraspinal locomotor centers in brainstem and cerebellum. Neuroimage 2008, 39, 786–792. [Google Scholar] [CrossRef]
- Imagawa, M.; Graf, W.; Sato, H.; Suwa, H.; Isu, N.; Izumi, R.; Uchino, Y. Morphology of single afferents of the saccular macula in cats. Neurosci. Lett. 1998, 240, 127–130. [Google Scholar] [CrossRef]
- Korte, G.E.; Mugnaini, E. Cerebellar projection of the vestibular nerve in the cat. J. Comp. Neurol. 1979, 184, 265–277. [Google Scholar] [CrossRef]
- Uchino, Y.; Sato, H.; Suwa, H. Excitatory and inhibitory inputs from saccular afferents to single vestibular neurons in the cat. J. Neurophysiol. 1997, 78, 2186–2192. [Google Scholar] [CrossRef] [PubMed]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. The basal ganglia communicate with the cerebellum. Proc. Natl. Acad. Sci. USA 2010, 107, 8452–8456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. Cerebellar networks with the cerebral cortex and basal ganglia. Trends Cogn. Sci. 2013, 17, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manto, M.; Argyropoulos, G.P.D.; Bocci, T.; Celnik, P.A.; Corben, L.A.; Guidetti, M.; Koch, G.; Priori, A.; Rothwell, J.C.; Sadnicka, A.; et al. Consensus Paper: Novel directions and next steps of non-invasive brain stimulation of the cerebellum in health and disease. Cerebellum 2021. [Google Scholar] [CrossRef]
- Miterko, L.N.; Baker, K.B.; Beckinghausen, J.; Bradnam, L.V.; Cheng, M.Y.; Cooperrider, J.; DeLong, M.R.; Gornati, S.V.; Hallett, M.; Heck, D.H.; et al. Consensus Paper: Experimental neurostimulation of the cerebellum. Cerebellum 2019, 18, 1064–1097. [Google Scholar] [CrossRef] [Green Version]
- Miterko, L.N.; Lin, T.; Zhou, J.; van der Heijden, M.E.; Beckinghausen, J.; White, J.J.; Sillitoe, R.V. Neuromodulation of the cerebellum rescues movement in a mouse model of ataxia. Nat. Commun. 2021, 12, 1295. [Google Scholar] [CrossRef]
- Ponce, G.V.; Klaus, J.; Schutter, D. A brief history of cerebellar neurostimulation. Cerebellum 2021. [Google Scholar] [CrossRef]
- Workman, C.D.; Fietsam, A.C.; Uc, E.Y.; Rudroff, T. Cerebellar transcranial direct current stimulation in people with Parkinson’s Disease: A pilot study. Brain Sci. 2020, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Hazrati, L.N.; Parent, A. Projection from the deep cerebellar nuclei to the pedunculopontine nucleus in the squirrel-monkey. Brain Res. 1992, 585, 267–271. [Google Scholar] [CrossRef]
- Raghu, A.L.B.; Parker, T.; Zand, A.P.D.; Payne, S.; Andersson, J.; Stein, J.; Aziz, T.Z.; Green, A.L. Tractography patterns of pedunculopontine nucleus deep brain stimulation. J. Neural Transm. 2021, 128, 659–670. [Google Scholar] [CrossRef]
- Tubert, C.; Galtieri, D.; Surmeier, D.J. The pedunclopontine nucleus and Parkinson’s disease. Neurobiol. Dis. 2019, 128, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Takakusaki, K.; Chiba, R.; Nozu, T.; Okumura, T. Brainstem control of locomotion and muscle tone with special reference to the role of the mesopontine tegmentum and medullary reticulospinal systems. J. Neural Transm. 2016, 123, 695–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, R.; Hass, C.J.; Sowalsky, K.; Schmitt, A.C.; Opri, E.; Roper, J.A.; Martinez-Ramirez, D.; Hess, C.W.; Foote, K.D.; Okun, M.S.; et al. Neurophysiological correlates of gait in the human basal ganglia and the PPN region in Parkinson’s Disease. Front. Hum. Neurosci. 2020, 14, 194. [Google Scholar] [CrossRef] [PubMed]
- Takakusaki, K. Functional neuroanatomy for posture and gait control. J. Mov. Disord. 2017, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Noga, B.R.; Guest, J.D. Combined neuromodulatory approaches in the central nervous system for treatment of spinal cord injury. Curr. Opin. Neurol. 2021, 34, 804–811. [Google Scholar] [CrossRef]
- Lai, H.; Tsumori, T.; Shiroyama, T.; Yokota, S.; Nakano, K.; Yasui, Y. Morphological evidence for a vestibulo-thalamo-striatal pathway via the parafascicular nucleus in the rat. Brain Res. 2000, 872, 208–214. [Google Scholar] [CrossRef]
- Stiles, L.; Smith, P.F. The vestibular-basal ganglia connection: Balancing motor control. Brain Res. 2015, 1597, 180–188. [Google Scholar] [CrossRef]
- Smith, P.F. The growing evidence for the importance of the otoliths in spatial memory. Front. Neural Circuits 2019, 13, 66. [Google Scholar] [CrossRef]
- Valjent, E.; Gangarossa, G. The tail of the striatum: From anatomy to connectivity and function. Trends Neurosci. 2021, 44, 203–214. [Google Scholar] [CrossRef]
- Cai, J.Y.; Lee, S.; Ba, F.; Garg, S.; Kim, L.J.; Liu, A.P.; Kim, D.; Wang, Z.J.; McKeown, M.J. Galvanic vestibular stimulation (GVS) augments deficient pedunculopontine nucleus (PPN) connectivity in mild Parkinson’s Disease: fMRI effects of different stimuli. Front. Neurosci. 2018, 12, 101. [Google Scholar] [CrossRef]
- Hitier, M.; Zhang, Y.F.; Sato, G.; Besnard, S.; Zheng, Y.W.; Smith, P.F. Stratification of hippocampal electrophysiological activation evoked by selective electrical stimulation of different angular and linear acceleration sensors in the rat peripheral vestibular system. Hear. Res. 2021, 403, 108173. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, A.; Hilber, P.; Chesneau, C.; Bulla, J.; Toulouse, J.; Machado, M.L.; Philoxene, B.; Smith, P.F.; Besnard, S. The critical role of vestibular graviception during cognitive-motor development. Behav. Brain Res. 2019, 372, 112040. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curthoys, I.S.; Smith, P.F.; de Miguel, A.R. Why Should Constant Stimulation of Saccular Afferents Modify the Posture and Gait of Patients with Bilateral Vestibular Dysfunction? The Saccular Substitution Hypothesis. J. Clin. Med. 2022, 11, 1132. https://doi.org/10.3390/jcm11041132
Curthoys IS, Smith PF, de Miguel AR. Why Should Constant Stimulation of Saccular Afferents Modify the Posture and Gait of Patients with Bilateral Vestibular Dysfunction? The Saccular Substitution Hypothesis. Journal of Clinical Medicine. 2022; 11(4):1132. https://doi.org/10.3390/jcm11041132
Chicago/Turabian StyleCurthoys, Ian S., Paul F. Smith, and Angel Ramos de Miguel. 2022. "Why Should Constant Stimulation of Saccular Afferents Modify the Posture and Gait of Patients with Bilateral Vestibular Dysfunction? The Saccular Substitution Hypothesis" Journal of Clinical Medicine 11, no. 4: 1132. https://doi.org/10.3390/jcm11041132