
Hematopoiesis, Inflammation and Aging—The Biological Background and Clinical Impact of Anemia and Increased C-Reactive Protein Levels on Elderly Individuals



Abstract
:1. Introduction
2. The Biological Context of Anemia in Elderly Individuals: Hallmarks of Aging in Normal Hematopoietic Cells and Their Associations with Signs of Inflammation
2.1. The Bone Marrow Microarchitecture and the Stem Cell Pool in Aging
2.2. Hematopoiesis and Hematopoietic Stem Cells in Elderly Individuals: Cell-Intrinsic Mechanisms Involved in Stem Cell Aging
2.2.1. Genetic Instability, Telomere Shortening, Altered Cell Communication and Inflammation
2.2.2. Epigenetic Abnormalities, Epigenetic Drift and Inflammation
2.2.3. Cellular Polarity and Epigenetic Asymmetry in Hematopoietic Stem Cell Division
2.2.4. Metabolic Characteristics and Regulation of Protein Homeostasis
2.2.5. Senescence and Intracellular Signaling
2.3. Stem-Cell-Extrinsic Mechanisms Involved in Hematopoietic Aging: Stem Cell Niches, Stromal Cell Subsets and Cellular Communications
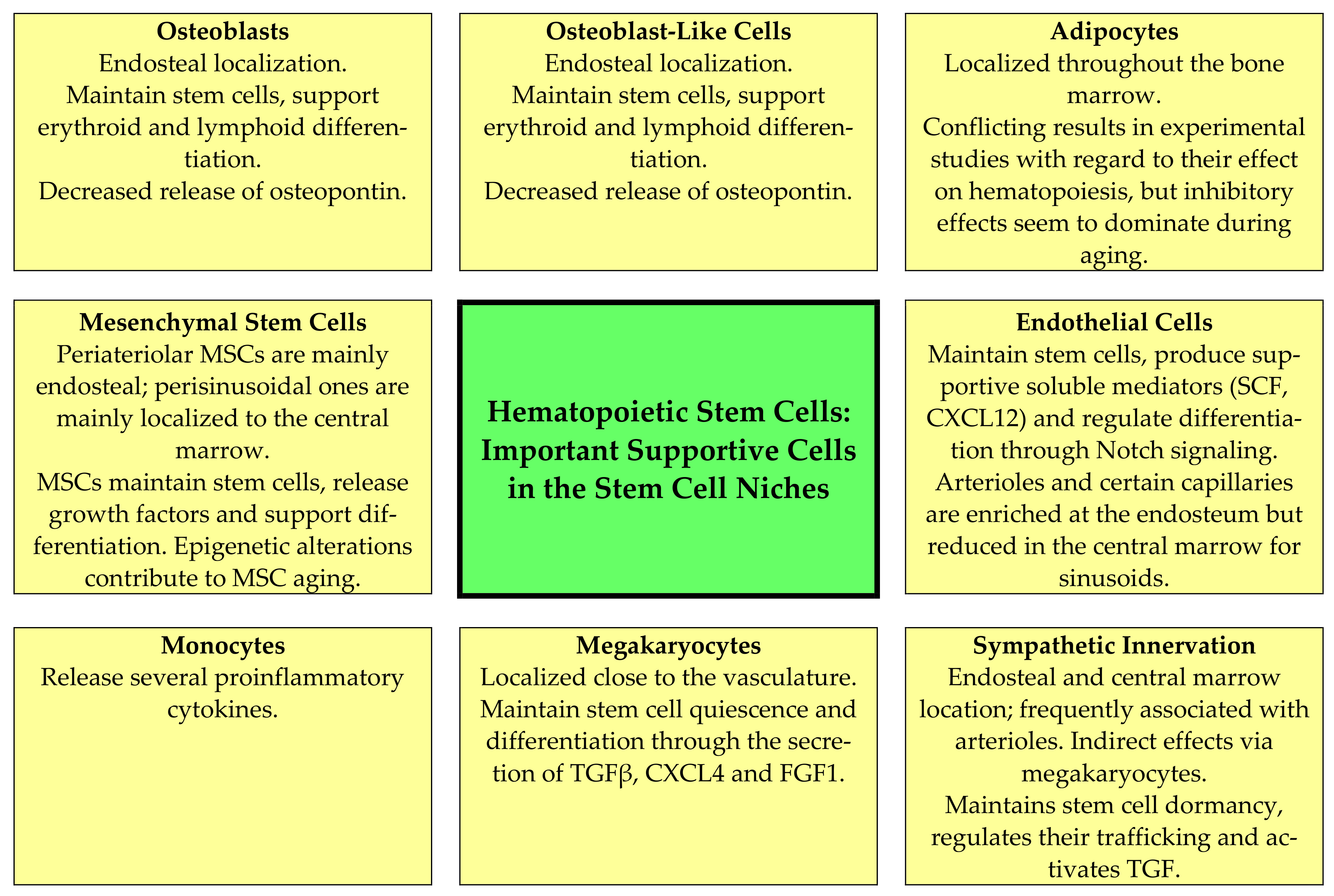
- Mesenchymal stem cells (MSCs). These cells are located close to arterioles and more loosely around sinusoidal vessels [17]; they are heterogeneous, and two main populations have been identified based on their expression of platelet-derived growth factor receptor (PDGFR)α and stem cell antigen 1 [17]. The MSCs support normal hematopoiesis through several mechanisms that are modulated by MSC aging, including their supportive function in stem cell niches (Figure 2, Table 2) [16,19,52,53,54,55,56,57,58,59,60,61]. Several epigenetic mechanisms are important for MSC aging and the alterations of their hematopoiesis-supporting mechanisms, including both altered DNA methylation and histone modification (e.g., acetylation) [61]; increased senescence is also observed [19,52,53,54,55,56,57,58,59,60,61]. First, MSCs produce several soluble mediators that are important both for myelopoiesis (e.g., CXCL12) and lymphopoiesis (e.g., IL7) [52]; the release of several growth factors is thus reduced. Second, MSCs and sinusoidal endothelial cells seem to form a complex network in close contact with the extracellular matrix and pervading the marrow tissue. The structural features of stromal components are maintained during aging [53], and although central perisinusoidal MSCs are increased or maintained there is a reduction in periarteriolar MSCs [16,19,56]. Third, the MSC functions in these networks seem to be altered, especially with regard to the regulation of cell cycle progression of the stem cells, stem cell trafficking in the microenvironment and the localization of progenitors close to different MSC subsets with different perivascular localizations [57]. Finally, the adipogenic differentiation of MSCs is preferred, and another consequence of this aging effect is reduced bone formation [59].
- Osteoblasts and other osteolineage cells. Osteoblasts are the predominant bone-lining endosteal cells [54], whereas osteolineage or osteoblastic lineage cells refer to the intermediate stages of differentiation in the direction from MSCs towards osteoblasts [57]. These cells and particularly mature osteoblasts seem to be the most important for the maintenance of more committed progenitors, especially lymphoid cells [62,63]. They stimulate/regulate hematopoiesis both through cell–cell contact (e.g., expression of the Notch ligand Jagged1) and through the release of soluble mediators (e.g., the growth factors CXCL12, stem cell factor and angiopoietin 1; osteopontin) [17]. Aging causes a decrease in the number of osteoblasts and in addition decreases osteopontin release via these aging cells (Figure 2, Table 2) [19,55,62,63]. As described above, aging MSCs favor adipogenic differentiation [59], and a reduction in osteoblasts is then caused by several mechanisms, including the induction of apoptosis, the increased release of reactive oxygen species, decreased glutathione reductase activity and the increased phosphorylation of p53 and p66 [64]. Finally, animal models suggest that a reduction in/lack of osteopontin causes decreased engraftment capacity but increases long-term stem cell frequency together with loss of stem cell polarity; as would then be expected from these observations, thrombin-activated osteopontin attenuates the aging stem cell phenotype [55].
- Adipocytes. Adipocytes release factors that seem to inhibit hematopoiesis [65,66,67]. Aging accelerates bone marrow adipogenesis [53]. This is apparent especially in the long bones where hematopoietic marrow is gradually replaced by adipocyte-rich marrow; although adipocytes release certain supportive mediators their overall effect is a reduction in hematopoiesis (Figure 2, Table 2) [19,66,68,69]. Animal studies suggest that this aging-associated adipocyte expansion can be further increased by dietary fat intake in aged animals [69]. The process of favored differentiation into adipocytes seems to be regulated at the transcriptional level and involves the transcriptional regulators Maf and Runx2 [58]. Furthermore, the release of adiponectin is a possible mechanism for the inhibition of hematopoiesis by adipocytes because this mediator has an antiproliferative and possibly also a proapoptotic effect, especially on myelomonocytic lineage cells [65]. The pharmacological inhibition of adipogenesis has therefore been suggested as a possible strategy to reduce the negative effects of adipogenesis on normal hematopoiesis [66,67]. However, the inhibitory effect of adipocytes may depend on the biological context, as a recent animal study has shown that adipocytes or a subset of adipocytes could release SCF and thereby promote regeneration after irradiation and myelotoxic chemotherapy [68].
- Endothelial cells. Endothelial cells and perivascular cells are intimately connected. Arteriolar and sinusoid endothelial cells seem to differ in their mechanisms with regard to supporting hematopoiesis (Figure 2) [54]; in particular, arteriolar cells release a wide range of hematopoietic growth factors [17,70,71]. The aging of endothelial cells has multiple effects on the stem cell niche and normal hematopoiesis (Table 2) [19,53,54,55,56,57,72,73]. First, the bone marrow endothelium shows aging-associated morphological and metabolic changes, including increased levels of reactive oxygen species that decrease their angiogenic and migratory potential, and the microvessels show a loss of integrity with augmented leakiness [70,71]. Second, the decreased release of prohematopoietic soluble mediators, including SCF and CXCL12, is one of the endothelial contributions to hematopoietic aging [70,72,74]. Third, the reduction in niche-forming vessels is likely to induce metabolic changes in the bone marrow microenvironment [70]. Finally, the niche-forming vessels in aging mice can be restored either by endothelial transplantation [72] or by the activation of endothelial Notch signaling, which seems to be altered in the aging bone marrow endothelium [56].
- Perivascular cells. This cell population is heterogeneous and includes cells expressing both pericyte and smooth muscle markers [17,70]. Aging reduces the abundance of pericytes and thereby the release of several soluble mediators that are important for the induction of quiescence of hematopoietic stem cells (e.g., SCF, bone morphogenic proteins 4 and 6) [70].
- Neural regulation. Sympathetic and sensory nerves innervate both the bone and the bone marrow [27]. Furthermore, human CD34+ cells express both dopaminergic and β2 adrenergic receptors; the receptors are expressed especially by immature CD34+CD38low cells and can be upregulated by G-CSF and GM-CSF [74]. Thus, adrenergic signals act directly on human hematopoietic progenitors and can increase their migration, proliferation, polarity as well as extracellular protease release, and Wnt-initiated signaling is involved in this stem cell modulation [74]. The perivascular arteriolar niche consists of specialized MSCs together with adrenergic nerves and megakaryocytes, and these cells are closely associated with quiescent stem cells [62,75,76,77]. Finally, nonmyelinating Schwann cells (i.e., glial fibrillary acidic protein-expressing cells) ensheath autonomic nerves, express genes that are important for the support of hematopoietic stem cells and can activate the latent form of TGFβ [76]. Thus, autonomic nerves are not only important through the direct effects of neurotransmitters on hematopoietic cells but also through their modulation of the niche cytokine network [76] and indirectly through the modulation of adrenoreceptor-expressing MSCs [77]. There is an aging-associated sympathetic denervation of the niche, and targeting this denervation with adrenoreceptor β3 agonists improves the function of aged stem cells in animal models [19,20,78]. Another effect of the denervation is the expansion of MSCs with decreased stem cell supporting capacity, a reduction in arterioles and increased stem cell numbers [16,19,54,57,74].
- Megakaryocytes. The role of megakaryocytes in the regulation of normal hematopoiesis can be regarded as a feedback mechanism. Megakaryocyte precursors migrate from the endosteal microenvironment to sinusoids for maturation, and noradrenergic bone marrow innervation promotes β2-adrenergic/IL6-dependent megakaryopoiesis [20]. A subset of hematopoietic stem cells is then associated with megakaryocytes that regulate stem cell quiescence through the release of soluble mediators (especially CXCL4 and TGFβ) as well as CD41 expression (Figure 2, Table 2) [18,19,37,54,57,79,80,81,82,83]. Age-dependent epigenetic alterations in hematopoietic stem cells and possibly also megakaryocytes seem to modify these interactions between megakaryocytes and neighboring hematopoietic stem cells [37]. Thus, there seems to be an interaction between aging, sympathetic innervation/denervation, epigenetic modulation and megakaryopoiesis with regard to the effects of aging on hematopoiesis [37]. The effects of megakaryocyte are partly mediated through the local release of TGFβ, which is important for the regulation of quiescence and initiates SMAD signaling in stem cells [79]; additionally, megakaryocytes release thrombopoietin [80] and CXCR4 [81], which act directly on immature hematopoietic cells. The release of thrombopoietin and possibly also other mediators can be stimulated by the ligation of C-type lectin-like receptor 2 (CLEC-2), and megakaryocyte expression of this receptor thereby becomes important for the regulation of stem cell quiescence [83]. Finally, the peripheral blood platelet count will possibly modulate these megakaryocyte effects through its effects on systemic thrombopoietin levels [37].
- Neutrophils. Neutrophils also seem to have regulatory functions in normal hematopoiesis; the mechanisms involve the neutrophil-mediated augmentation of sympathetic nervous system effects with the release of prostaglandin E2 [63]. These observations show that neutrophils can function as a link between the sympathetic nervous system and the stem cell niches.
- Monocytes, macrophages and osteoclasts. Both monocytes and other immunocompetent cells can contribute to the aging of hematopoiesis [17], possibly through their modulation of local levels of various proinflammatory cytokines [60]. Furthermore, aging seems to be associated with a shift from the anti-inflammatory M2 phenotype to the proinflammatory M1 phenotype; this shift is associated with the increased release of proinflammatory cytokines and seems to depend on monocyte/macrophage expression of the Foxo3 transcription factor (i.e., it is probably caused by an intrinsic mechanism) [11]. This shift is associated with local inflammation in the gastrointestinal tract, and in our opinion one should further investigate whether this shift is also important for inflammaging and/or the regulation of aging normal hematopoiesis. Finally, bone-marrow-associated macrophages are also important to maintain many stem-cell-supporting characteristics of MSCs, including their release of CXCL12 and SCF [57]. Finally, osteoclasts support hematopoiesis and lymphopoiesis indirectly by increasing the osteoblast secretion of CXCL12 and IL7 [57].
- T cells. As described in a recent review, activated T cells release several cytokines involved in the regulation of normal hematopoiesis [63]. CD4+ T cells thereby stimulate hematopoiesis, whereas CD4+CD25+ regulatory T cells seem to inhibit it. Furthermore, the clinical experience from allogeneic stem cell transplantation suggests that T cells facilitate engraftment, and even regulatory T cells (including CD150+ Treg cells) seem to facilitate engraftment and promote stem cell quiescence [63]. Finally, animal models suggest that CD8+ T cells also contribute to the regulation of hematopoiesis because IL6 and IFNγ released by CD8+ T cells can trigger emergency myelopoiesis [63].
2.4. Myeloid Skewing: An Intrinsic or Extrinsic Effect?
2.5. Inflammation and Hematopoiesis in Aging: The Contributions of Individual Cytokines and A Focus on the Myeloid Skewing
- IL1. IL1α/β exposure can induce myeloid skewing of normal hematopoietic stem cells at the expense of self-renewal [87]. Aging macrophages seem to stimulate megakaryocytic differentiation and myeloid skewing through IL1β-induced signaling [60,88]. IL1 also blocks the lymphoid differentiation of stem cells [89].
- IL6. This cytokine seems to promote thrombopoiesis (i.e., megakaryocyte modulation) [60].
- TNFα. This cytokine seems to stimulate myelopoiesis in aging [90].
- CCL5. This proinflammatory chemokine increases with age and seems to stimulate myeloid-biased differentiation [84].
- A certain subset of hematopoietic stem cells seems to respond to proinflammatory stimuli and thereby becomes particularly important for the myeloid skewing of hematopoiesis [90]. This study also suggests that young and aged long-term hematopoietic stem cells respond differently to inflammatory stress, such that the aged cells show a myeloid-biased gene expression initiated by several transcription factors, including Klf5, Ikzf1 and Stat3 [90].
2.6. Aging and Leukemic Hematopoiesis: Acute Myeloid Leukemia as an Example
3. Anemia in Elderly Individuals
3.1. Definition of Anemi
3.2. Anemia Is Common but Severe Anemia Is Uncommon in Elderly Individuals
3.3. Causes of Anemia in Elderly Individuals
- Six percent of the patients were diagnosed with a hematological malignancy, the most common being myelodysplastic syndrome (MDS), which was the suspected cause for 16% of the patients.
- Eleven percent had a non-hematological malignancy.
- Twelve percent had iron deficiency, but only a minority of these patients had microcytic anemia and for many patients the hemoglobin level did not normalize in response to iron supplementation. Iron deficiency was thus a possible contributing cause of anemia for many of these patients
- Renal failure was the cause of anemia for 4% of the patients.
- Long-lasting inflammation was the cause for 6% of the patients.
- Anemic patients and controls did not differ with regard to pharmacotherapy.
- For 35% of the individuals the cause of their anemia was not found. None of these individuals had hemoglobin levels below 9 g/dL (i.e., they probably did not require regular erythrocyte transfusions) and there was no association with ethnicity, age or sex. However, many of these patients had increased erythrocyte sedimentation rates and ferritin levels, i.e., they had systemic signs of an acute-phase reaction.
3.4. The Diagnostic Evaluation of Anemia in Elderly Patients
3.5. Anemia as a Prognostic Parameter in Community-Living Elderly Individuals
3.6. Anemia as a Prognostic Parameter in Nursing Home Residents
4. Causes of Mortality in Elderly Individuals with Anemia
4.1. Increased Mortality from Stroke
4.2. Increased Mortality from Heart Disease: Studies in Patients with Chronic Heart Failure
- One study included 6159 outpatients with stable chronic heart failure [141]. The prevalence of anemia was 17.2% (median age: 69 years for anemic versus 65 years for nonanemic); after six months 43% of these anemic patients at baseline had normalized Hb levels, whereas 16% of the nonanemic patients had developed anemia. After a mean follow-up of 3.9 years the mortality was higher both for patients with persistent anemia (58% vs. 31%, p < 0.0001) and incident anemia (45% vs. 31%, p < 0.0001) compared with nonanemic individuals at six months.
- A meta-analysis based on 153,180 heart failure patients included 37.2% anemic patients [142]; after a follow-up of at least six months the mortality was 46.8% for anemic and 29.5% for nonanemic patients. Lower baseline Hb was associated with higher mortality. These observations were also supported by another meta-analysis [143]: the patients in 10 of the 20 included reports had a mean age above 60 years, and an association between anemia and more severe heart failure was observed.
- The study by Kosiborod et al. [144] included 2281 patients aged 65 years or older with heart failure. This study showed that elderly patients with heart failure and anemia had higher one-year mortality.
4.3. Nutritional Defects and the Role of Iron Deficiency in Patients with Heart Failure
4.4. Anemia in Patients with Cancer
4.5. Increased Mortality of Anemic Patients after Surgery
4.6. Summarizing Comments: Anemia, Inflammation and Mortality
5. Inflammation in Aging
5.1. CRP as a Marker of Inflammation: Its Structure and Function
- Monocytes/macrophages: These cells can be polarized by CRP towards the proinflammatory M1 phenotype with increased phagocytosis and cytokine release, inhibited chemotaxis and altered metabolism with increased LDL uptake.
- Neutrophils: The functional CRP effects depend on the biological context and can be decreased activation, inhibition ofchemotaxis and/or stimulated phagocytosis [181].
- Endothelial cell activation.
- Thrombocytes: Inhibition of activation, trafficking and aggregation.
- Complement activation.
5.2. Inflammaging and Systemic CRP Levels in Elderly Patients
5.3. Inflammation-Induced Modulation of the Hematopoietic Stem Cell Niche
5.4. Inflammation, Aging, Disease and Mortality
6. Summarizing Discussion
6.1. Anemia and Aging
6.2. Carcinogenesis and Cancer Treatment in Elderly Individuals
6.3. Inflammation and Anemia
6.4. Anemia of an Unknown Cause
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudnicka, E.; Napierała, P.; Podfigurna, A.; Męczekalski, B.; Smolarczyk, R.; Grymowicz, M. The World Health Organization (WHO) approach to healthy ageing. Maturitas 2020, 139, 6–11. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijg, J.; Dong, X.; Milholland, B.; Zhang, L. Genome instability: A conserved mechanism of ageing? Essays Biochem. 2017, 61, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Morris, B.J.; Willcox, B.J.; Donlon, T.A. Genetic and epigenetic regulation of human aging and longevity. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1718–1744. [Google Scholar] [CrossRef]
- Micó, V.; Berninches, L.; Tapia, J.; Daimiel, L. NutrimiRAging: Micromanaging Nutrient Sensing Pathways through Nutrition to Promote Healthy Aging. Int. J. Mol. Sci. 2017, 18, 915. [Google Scholar] [CrossRef] [Green Version]
- Fakouri, N.B.; Hou, Y.; Demarest, T.G.; Christiansen, L.S.; Okur, M.N.; Mohanty, J.G.; Croteau, D.L.; Bohr, V.A. Toward understanding genomic instability, mitochondrial dysfunction and aging. FEBS J. 2019, 286, 1058–1073. [Google Scholar] [CrossRef]
- Klaips, C.L.; Jayaraj, G.G.; Hartl, F.U. Pathways of cellular proteostasis in aging and disease. J. Cell Biol. 2018, 217, 51–63. [Google Scholar] [CrossRef]
- Schmeer, C.; Kretz, A.; Wengerodt, D.; Stojiljkovic, M.; Witte, O.W. Dissecting Aging and Senescence-Current Concepts and Open Lessons. Cells 2019, 8, 1446. [Google Scholar] [CrossRef] [Green Version]
- Sameri, S.; Samadi, P.; Dehghan, R.; Salem, E.; Fayazi, N.; Amini, R. Stem Cell Aging in Lifespan and Disease: A State-of-the-Art Review. Curr. Stem Cell Res. Ther. 2020, 15, 362–378. [Google Scholar] [CrossRef]
- Becker, L.; Nguyen, L.; Gill, J.; Kulkarni, S.; Pasricha, P.J.; Habtezion, A. Age-dependent shift in macrophage polarisation causes inflammation-mediated degeneration of enteric nervous system. Gut 2018, 67, 827–836. [Google Scholar] [CrossRef] [Green Version]
- de Haan, G.; Lazare, S.S. Aging of hematopoietic stem cells. Blood 2018, 131, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akunuru, S.; Geiger, H. Aging, Clonality, and Rejuvenation of Hematopoietic Stem Cells. Trends Mol. Med. 2016, 22, 701–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejia-Ramirez, E.; Florian, M.C. Understanding intrinsic hematopoietic stem cell aging. Haematologica 2020, 105, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.J.; Bryder, D.; Zahn, J.M.; Ahlenius, H.; Sonu, R.; Wagers, A.J.; Weissman, I.L. Cell intrinsic alterations underlie hematopoietic stem cell aging. Proc. Natl. Acad. Sci. USA 2005, 102, 9194–9199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verovskaya, E.V.; Dellorusso, P.V.; Passegué, E. Losing Sense of Self and Surroundings: Hematopoietic Stem Cell Aging and Leukemic Transformation. Trends Mol. Med. 2019, 25, 494–515. [Google Scholar] [CrossRef]
- Pinho, S.; Frenette, P.S. Haematopoietic stem cell activity and interactions with the niche. Nat. Rev. Mol. Cell Biol. 2019, 20, 303–320. [Google Scholar] [CrossRef]
- Florian, M.C.; Dörr, K.; Niebel, A.; Daria, D.; Schrezenmeier, H.; Rojewski, M.; Filippi, M.D.; Hasenberg, A.; Gunzer, M.; Scharffetter-Kochanek, K.; et al. Cdc42 activity regulates hematopoietic stem cell aging and rejuvenation. Cell Stem Cell 2012, 10, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Maryanovich, M.; Zahalka, A.H.; Pierce, H.; Pinho, S.; Nakahara, F.; Asada, N.; Wei, Q.; Wang, X.; Ciero, P.; Xu, J.; et al. Adrenergic nerve degeneration in bone marrow drives aging of the hematopoietic stem cell niche. Nat. Med. 2018, 24, 782–791. [Google Scholar] [CrossRef]
- Ho, Y.H.; Del Toro, R.; Rivera-Torres, J.; Rak, J.; Korn, C.; García-García, A.; Macías, D.; González-Gómez, C.; Del Monte, A.; Wittner, M.; et al. Remodeling of Bone Marrow Hematopoietic Stem Cell Niches Promotes Myeloid Cell Expansion during Premature or Physiological Aging. Cell Stem Cell 2019, 25, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Saçma, M.; Pospiech, J.; Bogeska, R.; de Back, W.; Mallm, J.P.; Sakk, V.; Soller, K.; Marka, G.; Vollmer, A.; Karns, R.; et al. Haematopoietic stem cells in perisinusoidal niches are protected from ageing. Nat. Cell Biol. 2019, 21, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Wahlestedt, M.; Bryder, D. The slippery slope of hematopoietic stem cell aging. Exp. Hematol. 2017, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Beerman, I.; Bock, C.; Garrison, B.S.; Smith, Z.D.; Gu, H.; Meissner, A.; Rossi, D.J. Proliferation-dependent alterations of the DNA methylation landscape underlie hematopoietic stem cell aging. Cell Stem Cell 2013, 12, 413–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, S.M.; Shaw, C.A.; Gatza, C.; Fisk, C.J.; Donehower, L.A.; Goodell, M.A. Aging hematopoietic stem cells decline in function and exhibit epigenetic dysregulation. PLoS Biol. 2007, 5, e201. [Google Scholar] [CrossRef]
- Geiger, H.; Van Zant, G. The aging of lympho-hematopoietic stem cells. Nat. Immunol. 2002, 3, 329–333. [Google Scholar] [CrossRef]
- Lazzari, E.; Butler, J.M. The Instructive Role of the Bone Marrow Niche in Aging and Leukemia. Curr. Stem Cell Rep. 2018, 4, 291–298. [Google Scholar] [CrossRef]
- Rossi, D.J.; Bryder, D.; Seita, J.; Nussenzweig, A.; Hoeijmakers, J.; Weissman, I.L. Deficiencies in DNA damage repair limit the function of haematopoietic stem cells with age. Nature 2007, 447, 725–729. [Google Scholar] [CrossRef]
- Rudolph, K.L.; Chang, S.; Lee, H.W.; Blasco, M.; Gottlieb, G.J.; Greider, C.; DePinho, R.A. Longevity, stress response, and cancer in aging telomerase-deficient mice. Cell 1999, 96, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Kao, Y.R.; Sun, D.; Todorova, T.I.; Reynolds, D.; Narayanagari, S.R.; Montagna, C.; Will, B.; Verma, A.; Steidl, U. Myelodysplastic syndrome progression to acute myeloid leukemia at the stem cell level. Nat. Med. 2019, 25, 103–110. [Google Scholar] [CrossRef]
- Fuster, J.J.; MacLauchlan, S.; Zuriaga, M.A.; Polackal, M.N.; Ostriker, A.C.; Chakraborty, R.; Wu, C.L.; Sano, S.; Muralidharan, S.; Rius, C.; et al. Clonal hematopoiesis associated with TET2 deficiency accelerates atherosclerosis development in mice. Science 2017, 355, 842–847. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Natarajan, P.; Silver, A.J.; Gibson, C.J.; Bick, A.G.; Shvartz, E.; McConkey, M.; Gupta, N.; Gabriel, S.; Ardissino, D.; et al. Clonal Hematopoiesis and Risk of Atherosclerotic Cardiovascular Disease. N. Engl. J. Med. 2017, 377, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Busque, L.; Sun, M.; Buscarlet, M.; Ayachi, S.; Feroz Zada, Y.; Provost, S.; Bourgoin, V.; Mollica, L.; Meisel, M.; Hinterleitner, R.; et al. High-sensitivity C-reactive protein is associated with clonal hematopoiesis of indeterminate potential. Blood Adv. 2020, 4, 2430–2438. [Google Scholar] [CrossRef] [PubMed]
- Yura, Y.; Sano, S.; Walsh, K. Clonal Hematopoiesis: A New Step Linking Inflammation to Heart Failure. JACC Basic Transl. Sci. 2020, 5, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, R.S.; Jernigan, P.L.; Chang, A.L.; Edwards, M.J.; Pritts, T.A. Molecular mechanisms of erythrocyte aging. Biol. Chem. 2015, 396, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beerman, I.; Rossi, D.J. Epigenetic regulation of hematopoietic stem cell aging. Exp. Cell Res. 2014, 329, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Luo, M.; Jeong, M.; Rodriguez, B.; Xia, Z.; Hannah, R.; Wang, H.; Le, T.; Faull, K.F.; Chen, R.; et al. Epigenomic profiling of young and aged HSCs reveals concerted changes during aging that reinforce self-renewal. Cell Stem Cell 2014, 14, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Choudry, F.A.; Frontini, M. Epigenetic Control of Haematopoietic Stem Cell Aging and Its Clinical Implications. Stem Cells Int. 2016, 2016, 5797521. [Google Scholar] [CrossRef] [Green Version]
- Benayoun, B.A.; Pollina, E.A.; Singh, P.P.; Mahmoudi, S.; Harel, I.; Casey, K.M.; Dulken, B.W.; Kundaje, A.; Brunet, A. Remodeling of epigenome and transcriptome landscapes with aging in mice reveals widespread induction of inflammatory responses. Genome Res. 2019, 29, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Cheung, P.; Vallania, F.; Warsinske, H.C.; Donato, M.; Schaffert, S.; Chang, S.E.; Dvorak, M.; Dekker, C.L.; Davis, M.M.; Utz, P.J.; et al. Single-Cell Chromatin Modification Profiling Reveals Increased Epigenetic Variations with Aging. Cell 2018, 173, 1385–1397. [Google Scholar] [CrossRef] [Green Version]
- Grigoryan, A.; Guidi, N.; Senger, K.; Liehr, T.; Soller, K.; Marka, G.; Vollmer, A.; Markaki, Y.; Leonhardt, H.; Buske, C.; et al. LaminA/C regulates epigenetic and chromatin architecture changes upon aging of hematopoietic stem cells. Genome Biol. 2018, 19, 189. [Google Scholar] [CrossRef]
- Florian, M.C.; Klose, M.; Sacma, M.; Jablanovic, J.; Knudson, L.; Nattamai, K.J.; Marka, G.; Vollmer, A.; Soller, K.; Sakk, V.; et al. Aging alters the epigenetic asymmetry of HSC division. PLoS Biol. 2018, 16, e2003389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura-Ishizu, A.; Ito, K.; Suda, T. Hematopoietic Stem Cell Metabolism during Development and Aging. Dev. Cell 2020, 54, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, D.; Simic, M.S.; Dillin, A. Proteostasis and aging of stem cells. Trends. Cell Biol. 2014, 24, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Chapple, R.H.; Hu, T.; Tseng, Y.J.; Liu, L.; Kitano, A.; Luu, V.; Hoegenauer, K.A.; Iwawaki, T.; Li, Q.; Nakada, D. ERα promotes murine hematopoietic regeneration through the Ire1α-mediated unfolded protein response. Elife 2018, 7, e31159. [Google Scholar] [CrossRef]
- Moran-Crusio, K.; Reavie, L.B.; Aifantis, I. Regulation of hematopoietic stem cell fate by the ubiquitin proteasome system. Trends Immunol. 2012, 33, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Wang, Y.; Shao, L.; Laberge, R.M.; Demaria, M.; Campisi, J.; Janakiraman, K.; Sharpless, N.E.; Ding, S.; Feng, W.; et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat. Med. 2016, 22, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Quéré, R.; Saint-Paul, L.; Carmignac, V.; Martin, R.Z.; Chrétien, M.L.; Largeot, A.; Hammann, A.; Pais de Barros, J.P.; Bastie, J.N.; Delva, L. Tif1γ regulates the TGF-β1 receptor and promotes physiological aging of hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 10592–10597. [Google Scholar] [CrossRef] [Green Version]
- Mastelaro de Rezende, M.; Zenker Justo, G.; Julian Paredes-Gamero, E.; Gosens, R. Wnt-5A/B Signaling in Hematopoiesis throughout Life. Cells 2020, 9, 1801. [Google Scholar] [CrossRef]
- King, A.M.; Van der Put, E.; Blomberg, B.B.; Riley, R.L. Accelerated Notch-dependent degradation of E47 proteins in aged B cell precursors is associated with increased ERK MAPK activation. J. Immunol. 2007, 178, 3521–3529. [Google Scholar] [CrossRef]
- Xiao, N.; Jani, K.; Morgan, K.; Okabe, R.; Cullen, D.E.; Jesneck, J.L.; Raffel, G.D. Hematopoietic stem cells lacking Ott1 display aspects associated with aging and are unable to maintain quiescence during proliferative stress. Blood 2012, 119, 4898–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fistonich, C.; Zehentmeier, S.; Bednarski, J.J.; Miao, R.; Schjerven, H.; Sleckman, B.P.; Pereira, J.P. Cell circuits between B cell progenitors and IL-7+ mesenchymal progenitor cells control B cell development. J. Exp. Med. 2018, 215, 2586–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomariz, A.; Helbling, P.M.; Isringhausen, S.; Suessbier, U.; Becker, A.; Boss, A.; Nagasawa, T.; Paul, G.; Goksel, O.; Székely, G.; et al. Quantitative spatial analysis of haematopoiesis-regulating stromal cells in the bone marrow microenvironment by 3D microscopy. Nat. Commun. 2018, 9, 2532. [Google Scholar] [CrossRef] [PubMed]
- Crane, G.M.; Jeffery, E.; Morrison, S.J. Adult haematopoietic stem cell niches. Nat. Rev. Immunol. 2017, 17, 573–590. [Google Scholar] [CrossRef]
- Guidi, N.; Sacma, M.; Ständker, L.; Soller, K.; Marka, G.; Eiwen, K.; Weiss, J.M.; Kirchhoff, F.; Weil, T.; Cancelas, J.A.; et al. Osteopontin attenuates aging-associated phenotypes of hematopoietic stem cells. EMBO J. 2017, 36, 840–853. [Google Scholar] [CrossRef] [Green Version]
- Kusumbe, A.P.; Ramasamy, S.K.; Itkin, T.; Mäe, M.A.; Langen, U.H.; Betsholtz, C.; Lapidot, T.; Adams, R.H. Age-dependent modulation of vascular niches for haematopoietic stem cells. Nature 2016, 532, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Schepers, K.; Campbell, T.B.; Passegué, E. Normal and leukemic stem cell niches: Insights and therapeutic opportunities. Cell Stem Cell 2015, 16, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, K.; Nakashima, T.; Takeda, S.; Isogai, M.; Hamada, M.; Kimura, A.; Kodama, T.; Yamaguchi, A.; Owen, M.J.; Takahashi, S.; et al. Maf promotes osteoblast differentiation in mice by mediating the age-related switch in mesenchymal cell differentiation. J. Clin. Investig. 2010, 120, 3455–3465. [Google Scholar] [CrossRef] [Green Version]
- Singh, L.; Brennan, T.A.; Russell, E.; Kim, J.H.; Chen, Q.; Brad Johnson, F.; Pignolo, R.J. Aging alters bone-fat reciprocity by shifting in vivo mesenchymal precursor cell fate towards an adipogenic lineage. Bone 2016, 85, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.H.; Méndez-Ferrer, S. Microenvironmental contributions to hematopoietic stem cell aging. Haematologica 2020, 105, 38–46. [Google Scholar] [CrossRef]
- Cakouros, D.; Gronthos, S. Epigenetic Regulation of Bone Marrow Stem Cell Aging: Revealing Epigenetic Signatures associated with Hematopoietic and Mesenchymal Stem Cell Aging. Aging Dis. 2019, 10, 174–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Frenette, P.S. Niches for Hematopoietic Stem Cells and Their Progeny. Immunity 2018, 48, 632–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef] [Green Version]
- Yokota, T.; Oritani, K.; Takahashi, I.; Ishikawa, J.; Matsuyama, A.; Ouchi, N.; Kihara, S.; Funahashi, T.; Tenner, A.J.; Tomiyama, Y.; et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000, 96, 1723–1732. [Google Scholar] [CrossRef]
- Naveiras, O.; Nardi, V.; Wenzel, P.L.; Hauschka, P.V.; Fahey, F.; Daley, G.Q. Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment. Nature 2009, 460, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.J.; Wu, M.Q.; Li, Z.J.; Zhang, Y.; Liu, K.Y. Hematopoietic recovery following chemotherapy is improved by BADGE-induced inhibition of adipogenesis. Int. J. Hematol. 2013, 97, 58–72. [Google Scholar] [CrossRef]
- Zhou, B.O.; Yu, H.; Yue, R.; Zhao, Z.; Rios, J.J.; Naveiras, O.; Morrison, S.J. Bone marrow adipocytes promote the regeneration of stem cells and haematopoiesis by secreting SCF. Nat. Cell Biol. 2017, 19, 891–903. [Google Scholar] [CrossRef]
- Ambrosi, T.H.; Scialdone, A.; Graja, A.; Gohlke, S.; Jank, A.M.; Bocian, C.; Woelk, L.; Fan, H.; Logan, D.W.; Schürmann, A.; et al. Adipocyte Accumulation in the Bone Marrow during Obesity and Aging Impairs Stem Cell-Based Hematopoietic and Bone Regeneration. Cell Stem Cell 2017, 20, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Stucker, S.; Chen, J.; Watt, F.E.; Kusumbe, A.P. Bone Angiogenesis and Vascular Niche Remodeling in Stress, Aging, and Diseases. Front. Cell Dev. Biol. 2020, 8, 602269. [Google Scholar] [CrossRef]
- Ramasamy, S.K. Structure and Functions of Blood Vessels and Vascular Niches in Bone. Stem Cells Int. 2017, 2017, 5046953. [Google Scholar] [CrossRef] [Green Version]
- Poulos, M.G.; Ramalingam, P.; Gutkin, M.C.; Llanos, P.; Gilleran, K.; Rabbany, S.Y.; Butler, J.M. Endothelial transplantation rejuvenates aged hematopoietic stem cell function. J. Clin. Investig. 2017, 127, 4163–4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, A.; Shivtiel, S.; Kalinkovich, A.; Ludin, A.; Netzer, N.; Goichberg, P.; Azaria, Y.; Resnick, I.; Hardan, I.; Ben-Hur, H.; et al. Catecholaminergic neurotransmitters regulate migration and repopulation of immature human CD34+ cells through Wnt signaling. Nat. Immunol. 2007, 8, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Kunisaki, Y.; Frenette, P.S. Influences of vascular niches on hematopoietic stem cell fate. Int. J. Hematol. 2014, 99, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Ema, H.; Karlsson, G.; Yamaguchi, T.; Miyoshi, H.; Shioda, S.; Taketo, M.M.; Karlsson, S.; Iwama, A.; Nakauchi, H. Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 2011, 147, 1146–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Forte, D.; Krause, D.S.; Andreeff, M.; Bonnet, D.; Méndez-Ferrer, S. Updates on the hematologic tumor microenvironment and its therapeutic targeting. Haematologica 2019, 104, 1928–1934. [Google Scholar] [CrossRef]
- Jiang, L.; Han, X.; Wang, J.; Wang, C.; Sun, X.; Xie, J.; Wu, G.; Phan, H.; Liu, Z.; Yeh, E.T.H.; et al. SHP-1 regulates hematopoietic stem cell quiescence by coordinating TGF-β signaling. J. Exp. Med. 2018, 215, 1337–1347. [Google Scholar] [CrossRef]
- Nakamura-Ishizu, A.; Takubo, K.; Fujioka, M.; Suda, T. Megakaryocytes are essential for HSC quiescence through the production of thrombopoietin. Biochem. Biophys. Res. Commun. 2014, 454, 353–357. [Google Scholar] [CrossRef]
- Bruns, I.; Lucas, D.; Pinho, S.; Ahmed, J.; Lambert, M.P.; Kunisaki, Y.; Scheiermann, C.; Schiff, L.; Poncz, M.; Bergman, A.; et al. Megakaryocytes regulate hematopoietic stem cell quiescence through CXCL4 secretion. Nat. Med. 2014, 20, 1315–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Perry, J.M.; Marshall, H.; Venkatraman, A.; Qian, P.; He, X.C.; Ahamed, J.; Li, L. Megakaryocytes maintain homeostatic quiescence and promote post-injury regeneration of hematopoietic stem cells. Nat. Med. 2014, 20, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Ishizu, A.; Takubo, K.; Kobayashi, H.; Suzuki-Inoue, K.; Suda, T. CLEC-2 in megakaryocytes is critical for maintenance of hematopoietic stem cells in the bone marrow. J. Exp. Med. 2015, 212, 2133–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergen, A.V.; Boles, N.C.; Goodell, M.A. Rantes/Ccl5 influences hematopoietic stem cell subtypes and causes myeloid skewing. Blood 2012, 119, 2500–2509. [Google Scholar] [CrossRef] [Green Version]
- Pietras, E.M. Inflammation: A key regulator of hematopoietic stem cell fate in health and disease. Blood 2017, 130, 1693–1698. [Google Scholar] [CrossRef] [Green Version]
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef]
- Pietras, E.M.; Mirantes-Barbeito, C.; Fong, S.; Loeffler, D.; Kovtonyuk, L.V.; Zhang, S.; Lakshminarasimhan, R.; Chin, C.P.; Techner, J.M.; Will, B.; et al. Chronic interleukin-1 exposure drives haematopoietic stem cells towards precocious myeloid differentiation at the expense of self-renewal. Nat. Cell Biol. 2016, 18, 607–618. [Google Scholar] [CrossRef]
- Frisch, B.J.; Hoffman, C.M.; Latchney, S.E.; LaMere, M.W.; Myers, J.; Ashton, J.; Li, A.J.; Saunders, J., 2nd; Palis, J.; Perkins, A.S.; et al. Aged marrow macrophages expand platelet-biased hematopoietic stem cells via Interleukin1B. JCI Insight 2019, 5, e124213. [Google Scholar] [CrossRef]
- Montecino-Rodriguez, E.; Kong, Y.; Casero, D.; Rouault, A.; Dorshkind, K.; Pioli, P.D. Lymphoid-Biased Hematopoietic Stem Cells Are Maintained with Age and Efficiently Generate Lymphoid Progeny. Stem Cell Rep. 2019, 12, 584–596. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.; Mehta, A.; de Boer, C.G.; Kowalczyk, M.S.; Lee, K.; Haldeman, P.; Rogel, N.; Knecht, A.R.; Farouq, D.; Regev, A.; et al. Heterogeneous Responses of Hematopoietic Stem Cells to Inflammatory Stimuli Are Altered with Age. Cell Rep. 2018, 25, 2992–3005. [Google Scholar] [CrossRef] [Green Version]
- Sakamaki, S.; Hirayama, Y.; Matsunaga, T.; Kuroda, H.; Kusakabe, T.; Akiyama, T.; Konuma, Y.; Sasaki, K.; Tsuji, N.; Okamoto, T.; et al. Transforming growth factor-beta1 (TGF-beta1) induces thrombopoietin from bone marrow stromal cells, which stimulates the expression of TGF-beta receptor on megakaryocytes and, in turn, renders them susceptible to suppression by TGF-beta itself with high specificity. Blood 1999, 94, 1961–1970. [Google Scholar] [PubMed]
- Nagel, G.; Weber, D.; Fromm, E.; Erhardt, S.; Lübbert, M.; Fiedler, W.; Kindler, T.; Krauter, J.; Brossart, P.; Kündgen, A.; et al. Epidemiological, genetic, and clinical characterization by age of newly diagnosed acute myeloid leukemia based on an academic population-based registry study (AMLSG BiO). Ann. Hematol. 2017, 96, 1993–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Valladares, M.; Aasebø, E.; Berven, F.; Selheim, F.; Bruserud, Ø. Biological characteristics of aging in human acute myeloid leukemia cells: The possible importance of aldehyde dehydrogenase, the cytoskeleton and altered transcriptional regulation. Aging 2020, 12, 24734–24777. [Google Scholar] [CrossRef] [PubMed]
- Cappellini, M.D.; Motta, I. Anemia in Clinical Practice-Definition and Classification: Does Hemoglobin Change With Aging? Semin. Hematol. 2015, 52, 261–269. [Google Scholar] [CrossRef]
- Cappellini, M.D.; Beris, P. Anemia in Clinical Practice: Introduction. Semin. Hematol. 2015, 52, 259–260. [Google Scholar] [CrossRef]
- Martinsson, A.; Andersson, C.; Andell, P.; Koul, S.; Engström, G.; Smith, J.G. Anemia in the general population: Prevalence, clinical correlates and prognostic impact. Eur. J. Epidemiol. 2014, 29, 489–498. [Google Scholar] [CrossRef]
- Schop, A.; Stouten, K.; Riedl, J.A.; van Houten, R.J.; Leening, M.J.G.; van Rosmalen, J.; Bindels, P.J.E.; Levin, M.D. A new diagnostic work-up for defining anemia etiologies: A cohort study in patients ≥ 50 years in general practices. BMC Fam. Pract. 2020, 21, 167. [Google Scholar] [CrossRef]
- Guralnik, J.M.; Eisenstaedt, R.S.; Ferrucci, L.; Klein, H.G.; Woodman, R.C. Prevalence of anemia in persons 65 years and older in the United States: Evidence for a high rate of unexplained anemia. Blood 2004, 104, 2263–2268. [Google Scholar] [CrossRef] [Green Version]
- Westerlind, B.; Östgren, C.J.; Mölstad, S.; Midlöv, P. Prevalence and predictive importance of anemia in Swedish nursing home residents—A longitudinal study. BMC Geriatr. 2016, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Gaskell, H.; Derry, S.; Andrew Moore, R.; McQuay, H.J. Prevalence of anaemia in older persons: Systematic review. BMC Geriatr. 2008, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, E.A.; Mehra, R.; Holmes, T.H.; Schrier, S.L. Anemia in older persons: Etiology and evaluation. Blood Cells Mol. Dis. 2011, 46, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.; Ferrucci, L.; Guralnik, J. Anemia in older adults. Curr. Opin. Hematol. 2005, 12, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Cheungpasitporn, W.; Thongprayoon, C.; Chiasakul, T.; Korpaisarn, S.; Erickson, S.B. Renin-angiotensin system inhibitors linked to anemia: A systematic review and meta-analysis. QJM 2015, 108, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Guralnik, J.M.; Woodman, R.C.; Bandinelli, S.; Lauretani, F.; Corsi, A.M.; Chaves, P.H.; Ershler, W.B.; Longo, D.L. Proinflammatory state and circulating erythropoietin in persons with and without anemia. Am. J. Med. 2005, 118, 1288. [Google Scholar] [CrossRef] [PubMed]
- Stauder, R.; Valent, P.; Theurl, I. Anemia at older age: Etiologies, clinical implications, and management. Blood 2018, 131, 505–514. [Google Scholar] [CrossRef]
- Ferrucci, L.; Maggio, M.; Bandinelli, S.; Basaria, S.; Lauretani, F.; Ble, A.; Valenti, G.; Ershler, W.B.; Guralnik, J.M.; Longo, D.L. Low testosterone levels and the risk of anemia in older men and women. Arch. Intern. Med. 2006, 166, 1380–1388. [Google Scholar] [CrossRef] [Green Version]
- Waalen, J.; von Löhneysen, K.; Lee, P.; Xu, X.; Friedman, J.S. Erythropoietin, GDF15, IL6, hepcidin and testosterone levels in a large cohort of elderly individuals with anaemia of known and unknown cause. Eur. J. Haematol. 2011, 87, 107–116. [Google Scholar] [CrossRef]
- Monlezun, D.J.; Camargo, C.A., Jr.; Mullen, J.T.; Quraishi, S.A. Vitamin D Status and the Risk of Anemia in Community-Dwelling Adults: Results from the National Health and Nutrition Examination Survey 2001–2006. Medicine 2015, 94, e1799. [Google Scholar] [CrossRef]
- Ershler, W.B.; Sheng, S.; McKelvey, J.; Artz, A.S.; Denduluri, N.; Tecson, J.; Taub, D.D.; Brant, L.J.; Ferrucci, L.; Longo, D.L. Serum erythropoietin and aging: A longitudinal analysis. J. Am. Geriatr. Soc. 2005, 53, 1360–1365. [Google Scholar] [CrossRef]
- Beloosesky, Y.; Cohen, A.M.; Grosman, B.; Grinblat, J. Prevalence and survival of myelodysplastic syndrome of the refractory anemia type in hospitalized cognitively different geriatric patients. Gerontology 2000, 46, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Kwok, B.; Hall, J.M.; Witte, J.S.; Xu, Y.; Reddy, P.; Lin, K.; Flamholz, R.; Dabbas, B.; Yung, A.; Al-Hafidh, J.; et al. MDS-associated somatic mutations and clonal hematopoiesis are common in idiopathic cytopenias of undetermined significance. Blood 2015, 126, 2355–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cargo, C.A.; Rowbotham, N.; Evans, P.A.; Barrans, S.L.; Bowen, D.T.; Crouch, S.; Jack, A.S. Targeted sequencing identifies patients with preclinical MDS at high risk of disease progression. Blood 2015, 126, 2362–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steensma, D.P. New challenges in evaluating anemia in older persons in the era of molecular testing. Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Schop, A.; Stouten, K.; van Houten, R.; Riedl, J.; van Rosmalen, J.; Bindels, P.J.; Levin, M.D. Diagnostics in anaemia of chronic disease in general practice: A real-world retrospective cohort study. BJGP Open 2018, 2, bjgpopen18X101597. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.V.; Longo, D.L.; Ershler, W.B.; Yu, B.; Semba, R.D.; Ferrucci, L.; Guralnik, J.M. Haemoglobin concentration and the risk of death in older adults: Differences by race/ethnicity in the NHANES III follow-up. Br. J. Haematol. 2009, 145, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Penninx, B.W.; Pahor, M.; Woodman, R.C.; Guralnik, J.M. Anemia in old age is associated with increased mortality and hospitalization. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 474–479. [Google Scholar] [CrossRef] [Green Version]
- Shavelle, R.M.; MacKenzie, R.; Paculdo, D.R. Anemia and mortality in older persons: Does the type of anemia affect survival? Int. J. Hematol. 2012, 95, 248–256. [Google Scholar] [CrossRef]
- Zakai, N.A.; French, B.; Arnold, A.M.; Newman, A.B.; Fried, L.F.; Robbins, J.; Chaves, P.; Cushman, M. Hemoglobin decline, function, and mortality in the elderly: The cardiovascular health study. Am. J. Hematol. 2013, 88, 5–9. [Google Scholar] [CrossRef] [Green Version]
- den Elzen, W.P.; Willems, J.M.; Westendorp, R.G.; de Craen, A.J.; Assendelft, W.J.; Gussekloo, J. Effect of anemia and comorbidity on functional status and mortality in old age: Results from the Leiden 85-plus Study. CMAJ 2009, 181, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, T.C.; Yap, D.Y.; Shea, Y.F.; Luk, J.K.; Chan, F.H.; Chu, L.W. Prevalence of anemia in Chinese nursing home older adults: Implication of age and renal impairment. Geriatr. Gerontol. Int. 2013, 13, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Resnick, B.; Sabol, V.; Galik, E.; Gruber-Baldini, A.L. The impact of anemia on nursing home residents. Clin. Nurs. Res. 2010, 19, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Abid, S.A.; Gravenstein, S.; Nanda, A. Anemia in the Long-Term Care Setting. Clin. Geriatr. Med. 2019, 35, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Pandya, N.; Bookhart, B.; Mody, S.H.; Funk Orsini, P.A.; Reardon, G. Study of anemia in long-term care (SALT): Prevalence of anemia and its relationship with the risk of falls in nursing home residents. Curr. Med. Res. Opin. 2008, 24, 2139–2149. [Google Scholar] [CrossRef] [PubMed]
- Landi, F.; Russo, A.; Danese, P.; Liperoti, R.; Barillaro, C.; Bernabei, R.; Onder, G. Anemia status, hemoglobin concentration, and mortality in nursing home older residents. J. Am. Med. Dir. Assoc. 2007, 8, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.; Artz, A.S.; Culleton, B.; Critchlow, C.; Sciarra, A.; Audhya, P. Prevalence of anemia in the nursing home: Contribution of chronic kidney disease. J. Am. Geriatr. Soc. 2007, 55, 1566–1570. [Google Scholar] [CrossRef]
- Lucca, U.; Tettamanti, M.; Mosconi, P.; Apolone, G.; Gandini, F.; Nobili, A.; Tallone, M.V.; Detoma, P.; Giacomin, A.; Clerico, M.; et al. Association of mild anemia with cognitive, functional, mood and quality of life outcomes in the elderly: The “Health and Anemia” study. PLoS ONE 2008, 3, e1920. [Google Scholar] [CrossRef]
- Onder, G.; Penninx, B.W.; Cesari, M.; Bandinelli, S.; Lauretani, F.; Bartali, B.; Gori, A.M.; Pahor, M.; Ferrucci, L. Anemia is associated with depression in older adults: Results from the InCHIANTI study. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1168–1172. [Google Scholar] [CrossRef] [Green Version]
- Sabol, V.K.; Resnick, B.; Galik, E.; Ruber-Baldini, A.; Morton, P.G.; Hicks, G.E. Anemia and its impact on function in nursing home residents: What do we know? J. Am. Acad. Nurse Pract. 2010, 22, 3–16. [Google Scholar] [CrossRef]
- Bailey, R.A.; Reardon, G.; Wasserman, M.R.; McKenzie, R.S.; Hord, R.S. Association of anemia with worsened activities of daily living and health-related quality of life scores derived from the Minimum Data Set in long-term care residents. Health Qual Life Outcomes 2012, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Hopstock, L.A.; Utne, E.B.; Horsch, A.; Skjelbakken, T. The association between anemia and falls in community-living women and men aged 65 years and older from the fifth Tromsø Study 2001-02: A replication study. BMC Geriatr. 2017, 17, 292. [Google Scholar] [CrossRef] [PubMed]
- Dharmarajan, T.S.; Avula, S.; Norkus, E.P. Anemia increases risk for falls in hospitalized older adults: An evaluation of falls in 362 hospitalized, ambulatory, long-term care, and community patients. J. Am. Med. Dir. Assoc. 2007, 8, e9–e15. [Google Scholar] [CrossRef] [PubMed]
- Reardon, G.; Pandya, N.; Bailey, R.A. Falls in nursing home residents receiving pharmacotherapy for anemia. Clin. Interv. Aging 2012, 7, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, G. Frailty as a Predictor of Nursing Home Placement Among Community-Dwelling Older Adults: A Systematic Review and Meta-analysis. J. Geriatr. Phys. Ther. 2018, 41, 42–48. [Google Scholar] [CrossRef]
- Veronese, N.; Cereda, E.; Solmi, M.; Fowler, S.A.; Manzato, E.; Maggi, S.; Manu, P.; Abe, E.; Hayashi, K.; Allard, J.P.; et al. Inverse relationship between body mass index and mortality in older nursing home residents: A meta-analysis of 19,538 elderly subjects. Obes. Rev. 2015, 16, 1001–1015. [Google Scholar] [CrossRef] [Green Version]
- Doni, L.; Perin, A.; Manzione, L.; Gebbia, V.; Mattioli, R.; Speranza, G.B.; Latini, L.; Iop, A.; Bertetto, O.; Ferraù, F.; et al. The impact of anemia on quality of life and hospitalisation in elderly cancer patients undergoing chemotherapy. Crit. Rev. Oncol. Hematol. 2011, 77, 70–77. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, T.; Li, Y.; Chen, P.; Chen, L. Anemia increases the mortality risk in patients with stroke: A meta-analysis of cohort studies. Sci. Rep. 2016, 6, 26636. [Google Scholar] [CrossRef]
- Sico, J.J.; Concato, J.; Wells, C.K.; Lo, A.C.; Nadeau, S.E.; Williams, L.S.; Peixoto, A.J.; Gorman, M.; Boice, J.L.; Bravata, D.M. Anemia is associated with poor outcomes in patients with less severe ischemic stroke. J. Stroke Cerebrovasc. Dis. 2013, 22, 271–278. [Google Scholar] [CrossRef]
- Barlas, R.S.; Honney, K.; Loke, Y.K.; McCall, S.J.; Bettencourt-Silva, J.H.; Clark, A.B.; Bowles, K.M.; Metcalf, A.K.; Mamas, M.A.; Potter, J.F.; et al. Impact of Hemoglobin Levels and Anemia on Mortality in Acute Stroke: Analysis of UK Regional Registry Data, Systematic Review, and Meta-Analysis. J. Am. Heart Assoc. 2016, 5, e003019. [Google Scholar] [CrossRef] [Green Version]
- Barlas, R.S.; McCall, S.J.; Bettencourt-Silva, J.H.; Clark, A.B.; Bowles, K.M.; Metcalf, A.K.; Mamas, M.A.; Potter, J.F.; Myint, P.K. Impact of anaemia on acute stroke outcomes depends on the type of anaemia: Evidence from a UK stroke register. J. Neurol. Sci. 2017, 383, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.; Tong, W.; Jain, A.; Francis, G.S.; Harris, C.M.; Young, J.B. Evaluation and long-term prognosis of new-onset, transient, and persistent anemia in ambulatory patients with chronic heart failure. J. Am. Coll Cardiol. 2008, 51, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenveld, H.F.; Januzzi, J.L.; Damman, K.; van Wijngaarden, J.; Hillege, H.L.; van Veldhuisen, D.J.; van der Meer, P. Anemia and mortality in heart failure patients a systematic review and meta-analysis. J. Am. Coll Cardiol. 2008, 52, 818–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.W.; Wang, L.X. The impact of anemia on the prognosis of chronic heart failure: A meta-analysis and systemic review. Congest Heart Fail. 2009, 15, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosiborod, M.; Smith, G.L.; Radford, M.J.; Foody, J.M.; Krumholz, H.M. The prognostic importance of anemia in patients with heart failure. Am. J. Med. 2003, 114, 112–119. [Google Scholar] [CrossRef]
- Arora, N.P.; Ghali, J.K. Anemia and iron deficiency in heart failure. Heart Fail. Clin. 2014, 10, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Klip, I.T.; Comin-Colet, J.; Voors, A.A.; Ponikowski, P.; Enjuanes, C.; Banasiak, W.; Lok, D.J.; Rosentryt, P.; Torrens, A.; Polonski, L.; et al. Iron deficiency in chronic heart failure: An international pooled analysis. Am. Heart J. 2013, 165, 575–582.e3. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.; Kirwan, B.A.; Anker, S.D.; McDonagh, T.; Dorobantu, M.; Drozdz, J.; Fabien, V.; Filippatos, G.; Göhring, U.M.; Keren, A.; et al. Ferric carboxymaltose for iron deficiency at discharge after acute heart failure: A multicentre, double-blind, randomised, controlled trial. Lancet 2020, 396, 1895–1904. [Google Scholar] [CrossRef]
- Anker, S.D.; Kirwan, B.A.; van Veldhuisen, D.J.; Filippatos, G.; Comin-Colet, J.; Ruschitzka, F.; Lüscher, T.F.; Arutyunov, G.P.; Motro, M.; Mori, C.; et al. Effects of ferric carboxymaltose on hospitalisations and mortality rates in iron-deficient heart failure patients: An individual patient data meta-analysis. Eur. J. Heart Fail. 2018, 20, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.D.; Malhotra, R.; Hernandez, A.F.; McNulty, S.E.; Smith, A.; Felker, G.M.; Tang, W.H.W.; LaRue, S.J.; Redfield, M.M.; Semigran, M.J.; et al. Effect of Oral Iron Repletion on Exercise Capacity in Patients with Heart Failure with Reduced Ejection Fraction and Iron Deficiency: The IRONOUT HF Randomized Clinical Trial. JAMA 2017, 317, 1958–1966. [Google Scholar] [CrossRef]
- van der Wal, H.H.; Comin-Colet, J.; Klip, I.T.; Enjuanes, C.; Grote Beverborg, N.; Voors, A.A.; Banasiak, W.; van Veldhuisen, D.J.; Bruguera, J.; Ponikowski, P.; et al. Vitamin B12 and folate deficiency in chronic heart failure. Heart 2015, 101, 302–310. [Google Scholar] [CrossRef]
- Anand, S.; Burkenroad, A.; Glaspy, J. Workup of anemia in cancer. Clin. Adv. Hematol. Oncol. 2020, 18, 640–646. [Google Scholar] [PubMed]
- Gilreath, J.A.; Stenehjem, D.D.; Rodgers, G.M. Diagnosis and treatment of cancer-related anemia. Am. J. Hematol. 2014, 89, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Han, S.V.; Park, M.; Kwon, Y.M.; Yoon, H.J.; Chang, Y.; Kim, H.; Lim, Y.H.; Kim, S.G.; Ko, A. Mild Anemia and Risk for All-Cause, Cardiovascular and Cancer Deaths in Apparently Healthy Elderly Koreans. Korean J. Fam. Med. 2019, 40, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Wouters, H.J.C.M.; van der Klauw, M.M.; de Witte, T.; Stauder, R.; Swinkels, D.W.; Wolffenbuttel, B.H.R.; Huls, G. Association of anemia with health-related quality of life and survival: A large population-based cohort study. Haematologica 2019, 104, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Girelli, D.; Marchi, G.; Camaschella, C. Anemia in the Elderly. Hemasphere 2018, 2, e40. [Google Scholar] [CrossRef]
- Goddard, A.F.; James, M.W.; McIntyre, A.S.; Scott, B.B.; British Society of Gastroenterology. Guidelines for the management of iron deficiency anaemia. Gut 2011, 60, 1309–1316. [Google Scholar] [CrossRef] [Green Version]
- Nahon, S.; Lahmek, P.; Barclay, F.; Macaigne, G.; Poupardin, C.; Jounnaud, V.; Delas, N.; Lesgourgues, B. Long-term follow-up and predictive factors of recurrence of anemia in a cohort of 102 very elderly patients explored for iron-deficiency anemia. J. Clin. Gastroenterol. 2008, 42, 984–990. [Google Scholar] [CrossRef]
- Nahon, S.; Lahmek, P.; Aras, N.; Poupardin, C.; Lesgourgues, B.; Macaigne, G.; Delas, N. Management and predictors of early mortality in elderly patients with iron deficiency anemia: A prospective study of 111 patients. Gastroenterol. Clin. Biol. 2007, 31, 169–174. [Google Scholar] [CrossRef]
- Dicato, M.; Plawny, L.; Diederich, M. Anemia in cancer. Ann. Oncol. 2010, 21, vii167–vii172. [Google Scholar] [CrossRef]
- Endres, H.G.; Wedding, U.; Pittrow, D.; Thiem, U.; Trampisch, H.J.; Diehm, C. Prevalence of anemia in elderly patients in primary care: Impact on 5-year mortality risk and differences between men and women. Curr. Med. Res. Opin. 2009, 25, 1143–1158. [Google Scholar] [CrossRef]
- Lee, J.; Chin, J.H.; Kim, J.I.; Lee, E.H.; Choi, I.C. Association between red blood cell transfusion and long-term mortality in patients with cancer of the esophagus after esophagectomy. Dis. Esophagus. 2018, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Chen, D.; Jiang, Y.; Huang, W.; Ouyang, H.; Xing, W.; Zeng, M.; Xie, X.; Zeng, W. Impact of preoperative anemia on relapse and survival in breast cancer patients. BMC Cancer 2014, 14, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilreath, J.A.; Rodgers, G.M. How I treat cancer-associated anemia. Blood 2020, 136, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Fowler, A.J.; Ahmad, T.; Phull, M.K.; Allard, S.; Gillies, M.A.; Pearse, R.M. Meta-analysis of the association between preoperative anaemia and mortality after surgery. Br. J. Surg. 2015, 102, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Sundaram, A.; Mactaggart, J.N.; Johanning, J.M.; Gupta, H.; Fang, X.; Forse, R.A.; Balters, M.; Longo, G.M.; Sugimoto, J.T.; et al. Preoperative anemia is an independent predictor of postoperative mortality and adverse cardiac events in elderly patients undergoing elective vascular operations. Ann. Surg. 2013, 258, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.C.; Schifftner, T.L.; Henderson, W.G.; Eaton, C.B.; Poses, R.M.; Uttley, G.; Sharma, S.C.; Vezeridis, M.; Khuri, S.F.; Friedmann, P.D. Preoperative hematocrit levels and postoperative outcomes in older patients undergoing noncardiac surgery. JAMA 2007, 297, 2481–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, G.; Nowak, L.; Melo, L.; Ward, S.; Atrey, A.; Schemitsch, E.H.; Nauth, A.; Khoshbin, A. Anemia at Presentation Predicts Acute Mortality and Need for Readmission Following Geriatric Hip Fracture. JB JS Open Access 2020, 5, e20:00048. [Google Scholar] [CrossRef]
- Dubljanin-Raspopović, E.; Marković-Denić, L.; Nikolić, D.; Tulic, G.; Kadija, M.; Bumbasirevic, M. Is Anemia at admission related to short-term outcomes of elderly hip fracture patients? Cent. Eur. J. Med. 2011, 6, 483–489. [Google Scholar] [CrossRef]
- Bruserud, Ø.; Aarstad, H.H.; Tvedt, T.H.A. Combined C-Reactive Protein and Novel Inflammatory Parameters as a Predictor in Cancer-What Can We Learn from the Hematological Experience? Cancers 2020, 12, 1966. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Boncler, M.; Wu, Y.; Watala, C. The multiple faces of C-Reactive Protein-Physiological and pathophysiological implications in cardiovascular disease. Molecules 2019, 24, 2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Z.; Zhang, Y.; Wu, H. Regulation of C-reactive protein conformation in inflammation. Inflamm. Res. 2019, 68, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Potempa, L.A.; El Kebir, D.; Filep, J.G. C-reactive protein and inflammationEvenn: Conformational changes affect function. Biol. Chem. 2015, 396, 1181–1197. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Wu, Y.; Chen, Y.; Sui, S.F. Polymorphism of structural forms of C-reactive protein. Int. J. Mol. Med. 2002, 9, 665–671. [Google Scholar] [CrossRef]
- Li, Q.; Xu, W.; Xue, X.; Wang, Q.; Han, L.; Li, W.; Lv, S.; Liu, D.; Richards, J.; Shen, Z.; et al. Presence of multimeric isoforms of human C-reactive protein in tissues and blood. Mol. Med. Rep. 2016, 14, 5461–5466. [Google Scholar] [CrossRef] [Green Version]
- Okemefuna, A.I.; Stach, L.; Rana, S.; Buetas, A.J.; Gor, J.; Perkins, S.J. C-reactive protein exists in an NaCl concentration-dependent pentamer-decamer equilibrium in physiological buffer. J. Biol. Chem. 2010, 285, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- El Kebir, D.; Zhang, Y.; Potempa, L.A.; Wu, Y.; Fournier, A.; Filep, J.G. C-reactive protein-derived peptide 201-206 inhibits neutrophil adhesion to endothelial cells and platelets through CD32. J. Leukoc. Biol. 2011, 90, 1167–1175. [Google Scholar] [CrossRef]
- Van Vré, E.A.; Bult, H.; Hoymans, V.Y.; Van Tendeloo, V.F.; Vrints, C.J.; Bosmans, J.M. Human C-reactive protein activates monocyte-derived dendritic cells and induces dendritic cell-mediated T-cell activation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, R.V.; Wright, T.T.; Jones, N.R.; Wu, J.; Gibson, A.W.; Szalai, A.J. C-Reactive protein impairs dendritic cell development, maturation, and function: Implications for peripheral tolerance. Front. Immunol. 2018, 9, 372. [Google Scholar] [CrossRef]
- He, W.; Ren, Y.; Wang, X.; Chen, Q.; Ding, S. C reactive protein and enzymatically modified LDL cooperatively promote dendritic cell-mediated T cell activation. Cardiovasc. Pathol. 2017, 29, 1–6. [Google Scholar] [CrossRef]
- Bach, M.; Moon, J.; Moore, R.; Pan, T.; Nelson, J.L.; Lood, C. A Neutrophil Activation Biomarker Panel in Prognosis and Monitoring of Patients with Rheumatoid Arthritis. Arthritis Rheumatol. 2020, 72, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a promising target in slowing down the ageing process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perner, F.; Perner, C.; Ernst, T.; Heidel, F.H. Roles of JAK2 in Aging, Inflammation, Hematopoiesis and Malignant Transformation. Cells 2019, 8, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovtonyuk, L.V.; Fritsch, K.; Feng, X.; Manz, M.G.; Takizawa, H. Inflamm-Aging of Hematopoiesis, Hematopoietic Stem Cells, and the Bone Marrow Microenvironment. Front. Immunol. 2016, 7, 502. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Müller, L.; Di Benedetto, S.; Pawelec, G. The Immune System and Its Dysregulation with Aging. Subcell. Biochem. 2019, 91, 21–43. [Google Scholar]
- Fülöp, T.; Larbi, A.; Witkowski, J.M. Human Inflammaging. Gerontology 2019, 65, 495–504. [Google Scholar] [CrossRef]
- Cook, E.K.; Luo, M.; Rauh, M.J. Clonal hematopoiesis and inflammation: Partners in leukemogenesis and comorbidity. Exp. Hematol. 2020, 83, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Tvedt, T.H.A.; Melve, G.K.; Tsykunova, G.; Ahmed, A.B.; Brenner, A.K.; Bruserud, Ø. Immunological Heterogeneity of Healthy Peripheral Blood Stem Cell Donors-Effects of Granulocyte Colony-Stimulating Factor on Inflammatory Responses. Int. J. Mol. Sci. 2018, 19, 2886. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, H.; Manz, M.G. Impact of inflammation on early hematopoiesis and the microenvironment. Int. J. Hematol. 2017, 106, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sun, Q.; Morita, Y.; Jiang, H.; Gross, A.; Lechel, A.; Hildner, K.; Guachalla, L.M.; Gompf, A.; Hartmann, D.; et al. A differentiation checkpoint limits hematopoietic stem cell self-renewal in response to DNA damage. Cell 2012, 148, 1001–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.; Jia, T.; Mendez-Ferrer, S.; Hohl, T.M.; Serbina, N.V.; Lipuma, L.; Leiner, I.; Li, M.O.; Frenette, P.S.; Pamer, E.G. Bone marrow mesenchymal stem and progenitor cells induce monocyte emigration in response to circulating toll-like receptor ligands. Immunity 2011, 34, 590–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, N.T.; McGettrick, H.M.; Buckley, C.D.; Newsome, P.N.; Rainger, G.E.; Frampton, J.; Nash, G.B. Crosstalk between mesenchymal stem cells and endothelial cells leads to downregulation of cytokine-induced leukocyte recruitment. Stem Cells 2013, 31, 2690–2702. [Google Scholar]
- Fernandez, L.; Rodriguez, S.; Huang, H.; Chora, A.; Fernandes, J.; Mumaw, C.; Cruz, E.; Pollok, K.; Cristina, F.; Price, J.E.; et al. Tumor necrosis factor-alpha and endothelial cells modulate Notch signaling in the bone marrow microenvironment during inflammation. Exp. Hematol. 2008, 36, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Boettcher, S.; Gerosa, R.C.; Radpour, R.; Bauer, J.; Ampenberger, F.; Heikenwalder, M.; Kopf, M.; Manz, M.G. Endothelial cells translate pathogen signals into G-CSF-driven emergency granulopoiesis. Blood 2014, 124, 1393–1403. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Li, Y.; Jin, J.; Zhang, X.; Lopes-Virella, M.F.; Huang, Y. Toll-like receptor 4 activation in microvascular endothelial cells triggers a robust inflammatory response and cross talk with mononuclear cells via interleukin-6. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1696–1706. [Google Scholar] [CrossRef] [Green Version]
- Tracy, R.P.; Lemaitre, R.N.; Psaty, B.M.; Ives, D.G.; Evans, R.W.; Cushman, M.; Meilahn, E.N.; Kuller, L.H. Relationship of C-reactive protein to risk of cardiovascular disease in the elderly. Results from the Cardiovascular Health Study and the Rural Health Promotion Project. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1121–1127. [Google Scholar] [CrossRef]
- Cesari, M.; Penninx, B.W.; Newman, A.B.; Kritchevsky, S.B.; Nicklas, B.J.; Sutton-Tyrrell, K.; Tracy, R.P.; Rubin, S.M.; Harris, T.B.; Pahor, M. Inflammatory markers and cardiovascular disease (The Health, Aging and Body Composition [Health ABC] Study). Am. J. Cardiol. 2003, 92, 522–528. [Google Scholar] [CrossRef]
- Cesari, M.; Penninx, B.W.; Newman, A.B.; Kritchevsky, S.B.; Nicklas, B.J.; Sutton-Tyrrell, K.; Rubin, S.M.; Ding, J.; Simonsick, E.M.; Harris, T.B.; et al. Inflammatory markers and onset of cardiovascular events: Results from the Health ABC study. Circulation 2003, 108, 2317–2322. [Google Scholar] [CrossRef] [Green Version]
- Makita, S.; Nakamura, M.; Hiramori, K. The association of C-reactive protein levels with carotid intima-media complex thickness and plaque formation in the general population. Stroke 2005, 36, 2138–2142. [Google Scholar] [CrossRef] [Green Version]
- Hosford-Donovan, A.; Nilsson, A.; Wåhlin-Larsson, B.; Kadi, F. Observational and mechanistic links between C-reactive protein and blood pressure in elderly women. Maturitas 2016, 89, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Labonté, M.E.; Dewailly, E.; Chateau-Degat, M.L.; Couture, P.; Lamarche, B. Population-based study of high plasma C-reactive protein concentrations among the Inuit of Nunavik. Int. J. Circumpolar. Health 2012, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kritchevsky, S.B.; Cesari, M.; Pahor, M. Inflammatory markers and cardiovascular health in older adults. Cardiovasc. Res. 2005, 66, 265–275. [Google Scholar] [CrossRef]
- Nadrowski, P.; Chudek, J.; Skrzypek, M.; Puzianowska-Kuźnicka, M.; Mossakowska, M.; Więcek, A.; Zdrojewski, T.; Grodzicki, T.; Kozakiewicz, K. Associations between cardiovascular disease risk factors and IL-6 and hsCRP levels in the elderly. Exp. Gerontol. 2016, 85, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Shivappa, N.; Wirth, M.D.; Hurley, T.G.; Hébert, J.R. Association between the dietary inflammatory index (DII) and telomere length and C-reactive protein from the National Health and Nutrition Examination Survey-1999–2002. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Varadhan, R.; Yao, W.; Matteini, A.; Beamer, B.A.; Xue, Q.L.; Yang, H.; Manwani, B.; Reiner, A.; Jenny, N.; Parekh, N.; et al. Simple biologically informed inflammatory index of two serum cytokines predicts 10 year all-cause mortality in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 165–173. [Google Scholar] [CrossRef]
- McCabe, E.L.; Larson, M.G.; Lunetta, K.L.; Newman, A.B.; Cheng, S.; Murabito, J.M. Association of an Index of Healthy Aging With Incident Cardiovascular Disease and Mortality in a Community-Based Sample of Older Adults. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Hamann, L.; Bustami, J.; Iakoubov, L.; Szwed, M.; Mossakowska, M.; Schumann, R.R.; Puzianowska-Kuznicka, M. TLR-6 SNP P249S is associated with healthy aging in nonsmoking Eastern European Caucasians—A cohort study. Immun. Ageing 2016, 17, 13:7. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Fung, E.; Xu, A.; Lan, H.Y. C-reactive protein and ageing. Clin. Exp. Pharmacol. Physiol. 2017, 44, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Kupelian, V.; Chiu, G.R.; Araujo, A.B.; Williams, R.E.; Clark, R.V.; McKinlay, J.B. Association of sex hormones and C-reactive protein levels in men. Clin. Endocrinol. 2010, 72, 527–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacho, J.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. C-reactive protein and all-cause mortality—the Copenhagen City Heart Study. Eur. Heart J. 2010, 31, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Barron, E.; Lara, J.; White, M.; Mathers, J.C. Blood-borne biomarkers of mortality risk: Systematic review of cohort studies. PLoS ONE 2015, 10, e0127550. [Google Scholar] [CrossRef] [PubMed]
- Abel, G.A.; Klepin, H.D. Frailty and the management of hematologic malignancies. Blood 2018, 131, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Gadalla, S.M.; Wang, T.; Dagnall, C.; Haagenson, M.; Spellman, S.R.; Hicks, B.; Jones, K.; Katki, H.A.; Lee, S.J.; Savage, S.A. Effect of Recipient Age and Stem Cell Source on the Association between Donor Telomere Length and Survival after Allogeneic Unrelated Hematopoietic Cell Transplantation for Severe Aplastic Anemia. Biol. Blood Marrow. Transplant. 2016, 22, 2276–2282. [Google Scholar] [CrossRef]
- Spólnicka, M.; Piekarska, R.Z.; Jaskuła, E.; Basak, G.W.; Jacewicz, R.; Pięta, A.; Makowska, Ż.; Jedrzejczyk, M.; Wierzbowska, A.; Pluta, A.; et al. Donor age and C1orf132/MIR29B2C determine age-related methylation signature of blood after allogeneic hematopoietic stem cell transplantation. Clin. Epigenetics 2016, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Fabre, C.; Koscielny, S.; Mohty, M.; Fegueux, N.; Blaise, D.; Maillard, N.; Tabrizi, R.; Michallet, M.; Socié, G.; Yakoub-Agha, I.; et al. Younger donor’s age and upfront tandem are two independent prognostic factors for survival in multiple myeloma patients treated by tandem autologous-allogeneic stem cell transplantation: A retrospective study from the Société Française de Greffe de Moelle et de Thérapie Cellulaire (SFGM-TC). Haematologica 2012, 97, 482–490. [Google Scholar]
- Lazarevic, V.L. Acute myeloid leukaemia in patients we judge as being older and/or unfit. J. Intern. Med. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Magliano, G.; Bacigalupo, A. Allogeneic Hematopoietic Stem Cell Transplantation for Acute Myeloid Leukemia of the Elderly: Review of Literature and New Perspectives. Mediterr. J. Hematol. Infect. Dis. 2020, 1, e2020081. [Google Scholar] [CrossRef]
- Leufven, E.; Bruserud, Ø. Immunosuppression and Immunotargeted Therapy in Acute Myeloid Leukemia—The Potential Use of Checkpoint Inhibitors in Combination with Other Treatments. Curr. Med. Chem. 2019, 26, 5244–5261. [Google Scholar] [CrossRef]
- Chittezhath, M.; Dhillon, M.K.; Lim, J.Y.; Laoui, D.; Shalova, I.N.; Teo, Y.L.; Chen, J.; Kamaraj, R.; Raman, L.; Lum, J.; et al. Molecular profiling reveals a tumor-promoting phenotype of monocytes and macrophages in human cancer progression. Immunity 2014, 41, 815–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarstad, H.H.; Moe, S.E.E.; Bruserud, Ø.; Lybak, S.; Aarstad, H.J.; Tvedt, T.H.A. The Acute Phase Reaction and Its Prognostic Impact in Patients with Head and Neck Squamous Cell Carcinoma: Single Biomarkers Including C-Reactive Protein Versus Biomarker Profiles. Biomedicines 2020, 8, 418. [Google Scholar] [CrossRef] [PubMed]
- Aarstad, H.H.; Guðbrandsdottir, G.; Hjelle, K.M.; Bostad, L.; Bruserud, Ø.; Tvedt, T.H.A.; Beisland, C. The Biological Context of C-Reactive Protein as a Prognostic Marker in Renal Cell Carcinoma: Studies on the Acute Phase Cytokine Profile. Cancers 2020, 12, 1961. [Google Scholar] [CrossRef] [PubMed]
- Kip, M.M.; Schop, A.; Stouten, K.; Dekker, S.; Dinant, G.J.; Koffijberg, H.; Bindels, P.J.; IJzerman, M.J.; Levin, M.D.; Kusters, R. Assessing the cost-effectiveness of a routine versus an extensive laboratory work-up in the diagnosis of anaemia in Dutch general practice. Ann. Clin. Biochem. 2018, 55, 630–638. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genetic: Genetic instability with accumulating DNA damage and clonal hematopoiesis; this is due to the altered function of several mechanisms involved in genomic maintenance/DNA repair. |
| Epigenetic: Epigenetic modulation with altered chromatin organization, posttranscriptional histone modulation and DNA methylation; transcriptional regulation is thereby altered. |
| Polarity: Reduced cytoplasmic and nuclear polarity, reduced ability of asymmetric cell division. |
| Metabolism: A shift to higher oxidative metabolism, altered proteostasis due to reduced autophagy and reduced activity of the proteasome system, reduced endoplasmic reticulum stress response with the accumulation of misfolded or damaged proteins. |
| Senescence, signaling and communication: Accumulation of cell-cycle-arrested senescent cells, altered intercellular communication and intracellular signaling possibly involving auto- and paracrine circuits, reduced regenerative capacity of hematopoietic stem cells. |
| Stromal Component | Important Effect of Aging |
|---|---|
| MSCs [19,52,53,54,55,56,57,58,59,60,61] | Maintained or increased central MSCs with decreased hematopoietic growth factor production; loss of periarteriolar MSCs. Decreased bone formation and increased adipogenesis; MSC aging is also characterized by increased senescence (including altered mediator secretion) and epigenetic modifications. |
| Osteoblastic cells [19,55,62,63] | Decreased number of osteoblasts, increased differentiation in the direction of adipocytes. Decreased numbers of osteoblasts represent decreased support of lymphopoiesis. Decreased osteopontin release; this cytokine can attenuate the aging-associated phenotype of hematopoietic stem cells. |
| Osteoblasts [27,64] | Decreased number and release of osteopontin, decreased osteoblast number and thereby reduced support of lymphopoiesis. |
| Adipocytes [19,65,66,68,69] | Increased number of adipocytes during aging. The effects of adipocytes on normal hematopoiesis depend on the biological context. Hematopoiesis is often suppressive, and the release of adiponectin probably contributes to this inhibition. However, adipocytes or a subset of them also seem to facilitate regeneration after chemotherapy or irradiation through their release of SCF. |
| Endothelial cells [19,53,54,55,56,57,72,73] | Loss of certain capillaries and arterioles, decreased release of SCF and CXCL12 in addition to decreased expression of the Notch ligand Jagged1. Increased or unaltered endothelial cell pool, altered microvascular function with increased vascular leak. |
| Perivascular cells [17,70] | Aging of these cells is associated with reduced number of these cells and thereby reduced release of soluble stem cell-supporting mediators, e.g., SCF. |
| Sympathetic innervation [19,54,57,74] | Loss of sympathetic innervation in aging; this leads to expansion of medullary MSCs with decreased supportive effect, reduces the number of arterioles and increases the hematopoietic stem cell number. |
| Megakaryocytes [18,19,54,57,79,80,81,82,83] | Increased number and TGFβ release in aging. |
| Definitions of Anemia [95,96,99] | ||
|---|---|---|
| Definition Men | Definition Women | |
| WHO definition of anemia (Hb) | <13 g/dL | <12 g/dL |
| Alternative definitions | <12 g/dL | <12 g/dL |
| Decrease in Hb > 2 g/dL | Decrease in Hb > 2 g/dL | |
| Prevalence of Hemoglobin Levels in Elderly Individuals [97] | ||
| Percent of Individuals | Criteria Men | Criteria Women |
| 22.0% | <14 g/dL | <13 g/dL |
| 5.6% | <13.2 g/dL | <12.2 g/dL |
| 3.8% | <13.0 g/dL | <12.0 g/dL |
| 0.6% | <11.0 g/dL | <11 g/dL |
| Prevalence of Anemia in Various Subsets of Elderly Patients [99,100,101] | ||
| Percent of Individuals | Subset of Elderly Individuals | |
| 12% | Elderly living in private homes | |
| 47% | Elderly living in nursery homes | |
| 40% | Elderly admitted to hospital | |
| Cause of Anemia | Percent of Patients |
|---|---|
| Total fraction: malnutrition, specific deficiencies | 20% |
| Folic acid/cobalamin deficiency | 12–15% |
| Iron deficiency | 20% |
| Renal failure | 8–10% |
| Other chronic diseases, including inflammatory diseases | 20% |
| Renal failure combined with another chronic disease | <5% |
| Multiple etiologies | 20% |
| Unknown cause of anemia | 30–35% |
| Type of Marker | Recommended Single Analyses (Peripheral Blood) |
|---|---|
| Peripheral blood cells | Hemoglobin, MCV, MCH, differential blood cell count, reticulocyte count, reticulocyte hemoglobin and erythropoietin |
| Nutritional status | Vitamin B12, serum folate, transferrin saturation and ferritin |
| Hemolysis | Lactate dehydrogenase, haptoglobin and bilirubin |
| Organ markers | Creatinine and glomerular filtration rate Alanine aminotransferase and aspartate aminotransferase |
| Markers of inflammation | C-reactive protein |
| Others | Serum electrophoresis and thyrotropin-releasing hormone |
| Study | Population and Methodology | Observation |
|---|---|---|
| Schop et al. [98] | The study included 4152 individuals from the general population above 50 years of age (median age: 75). Newly diagnosed anemia. | After an extensive evaluation in general practice the cause was unclear for 20%, one cause was seen for 59% and multiple etiologies for 22%. The most common single etiologies were anemia of chronic disease and iron deficiency. The frequency of patients with renal anemia increased with age. |
| Patel et al. [116] | The study included 4089 Americans above 65 years of age. | For non-Hispanic white Americans the mortality increased with the degree of anemia, and the anemia threshold for increased mortality corresponded to 0.4 and 0.2 g/100 mL above the WHO definition of anemia (see Table 2). For black Americans the threshold for increased mortality was 0.7 g/100 mL below the WHO definition. |
| Guralnik et al. [99] | A population-based study including 39,695 individuals, 5252 of them being older than 65. | Anemia prevalence rates increased after 50 years of age. For individuals ≥65 years of age 11.0% of men and 10.2% of women were anemic, and 20% of individuals ≥85 years of age were anemic. Nutrient deficiency was present in one-third, one-third had renal and/or chronic inflammatory anemia and the anemia was unexplained for one-third. Hb levels <11.0 were observed for 1.6% of men and 2.8% of women. Anemia was most frequent in elderly black people (27.8%) and less frequent in Mexican Americans (10.4%) and white non-Hispanics (9.0%). |
| Penninx et al. [117] | The study included 3607 individuals aged 71 or older, with a mean age of 78.2. | Anemia according to the WHO criteria was observed for 12.5%. The mortality was significantly higher for anemic participants (37.0% vs. 22.1%, p < 0.001) and they were hospitalized more frequently and spent more days in hospital. These differences remained significant after excluding persons with prevalent disease. |
| Shavelle et al. [118] | The study included 7171 community-dwelling individuals (aged ≥ 50), 862 of whom were anemic according to the WHO definition. | Significant negative impact of anemia on overall survival with relative risk 1.8 (p < 0.001). Relative risk depended on cause: (i) nutritional (2.34, p < 0.0001); (ii) chronic renal disease (1.70, p < 0.0001); (iii) chronic inflammation (1.48, p < 0.0001); and (iv) unexplained (1.26, p < 0.01). |
| Zakai et al. [119] | The development of anemia was evaluated for 3758 community-dwelling individuals aged 65 or older without anemia at inclusion. | Of the individuals, 498 (8.5%) developed anemia according to the WHO criteria. Baseline increasing age, being African American and kidney disease predicted anemia development over 3 years. Both anemia development and hemoglobin decline predicted subsequent mortality in men and women. |
| den Enzen [120] | A population-based study of 562 individuals aged 85. | The prevalence of anemia at baseline was 26.7%, and anemic individuals had more comorbidity with more disabilities, worse cognitive function and more depressive symptoms. Both prevalent and incident anemia was significantly associated with survival in adjusted analyses, including adjustment for C-reactive protein. Mortality increased with severity of anemia. |
| Study | Population and Methodology | Observation |
|---|---|---|
| Chan et al. [121] | Retrospective, cross-sectional study at nine Chinese nursing homes (812 residents, mean age of 86 years). | A total of 67% were anemic, and the anemic residents were older and had a higher incidence of renal impairment; no significant associations with other comorbidities were observed. |
| Resnick et al. [122] | Including 451 residents, mean age of 83.7 years. | Anemia was more common among black than white residents; physical capacity was worse in anemic patients. |
| Westerlind et al. [100] | Including 390 patients (mean age 85.1 years), follow-up 7 years from baseline including Hb for 220 patients. | Prevalence of anemia at baseline was 52% for men and 32% for women. Two-year mortality was 61% for men with and 29% for men without anemia (p = 0.001), but for women no significant difference was observed (49% vs. 43%). Increased mortality in anemic men was independent of age, BNP and eGFR. Among men, anemia correlated with BNP/eGFR/CRP; for women, anemia correlated with several inflammatory markers including CRP. Anemic men were less physically active. Reduction in Hb with more than 0.9 g/100 mL during the first 2 years of follow-up was associated with increased mortality. |
| Pandya et al. [124] | Including 564 residents, mean age of 81 years. | In this study, 64% of males and 53% of females were anemic. Anemia was significantly associated with being African American, low eGFR, cancer, gastrointestinal bleeding and inflammatory disease, |
| Landi et al. [125] | Including 372 residents admitted to nursing home, aged 65 years or older. | At enrolment 63.1% of patients were anemic according to the WHO criteria. The death rate of anemic patients (38%) was higher than for nonanemic patients (28%, p = 0.03). This difference was independent of frailty, cognitive impairment, eGFR, cancer, stroke, body mass index and pressure ulcer. |
| Robinson et al. [126] | Evaluated 6200 residents, mean age of 83.2 years. | Of the residents, 59.6% were anemic. Older age was associated with lower hemoglobin in patients without kidney disease. However, for the whole study population chronic kidney disease seemed to contribute more strongly to the development of anemia than high age. |
| Study and Study Population | Observation |
|---|---|
| Tracy et al. [198]. Prospective, nested case–control study. There were 5201 persons in the original sample, age ≥ 65. Of these, 146 cases with incident cardiovascular events were identified and 146 control subjects were matched (mean age of 72.8 and 72.9 years, respectively). | The mean CRP level was only significantly higher for case subjects than for control subjects in women. CRP levels were generally higher in persons with subclinical disease. Among the elderly with subclinical disease, CRP was associated with myocardial infarction with an overall odds ratio (OR) of 2.67 (CI 1.04–6.81), with the association being stronger for women, with an OR of 4.50 (CI 0.97 to 20.8), than for men, with an OR of 1.75 (CI 0.51 to 5.98). |
| Cesari et al. [199]. Cross-sectional study including 3045 well-functioning persons with a mean age of 74.2 years (range of 70–79 years). Subclinical cardiovascular disease was defined as signs of angina or claudication according to the Rose Questionnaire, positive ankle/brachial index or electrocardiographic abnormalities. | CRP was significantly associated with congestive heart failure, with an OR of 1.64 (95% CI 1.11 to 2.41), but not with any other manifestations of cardiovascular disease. When comparing patients in the highest versus those in the lowest IL6 tertile, the OR for subclinical cardiovascular disease was 1.58 (95% CI 1.26 to 1.97) and for clinical cardiovascular disease was 2.35 (95% 1.79 to 3.09). This was similar for TNFα, with an OR of 1.48 (95% CI 1.16 to 1.88) and 2.05 (95% CI 1.55 to 2.72), respectively. Only soluble TNFR1 but not soluble IL6R or soluble IL2R showed a significant association with cardiovascular disease. |
| Cesari et al. [200]. Prospective study including 2225 persons with a mean age of 74.0 years (range of 70–79 years). Outcomes were hospitalizations for coronary heart disease (CHD), stroke or congestive heart failure (CHF). | CRP was significantly associated only with congestive heart failure. In contrast, IL6 was significantly associated with all three outcomes, and TNFα was significantly associated both with coronary heart disease and congestive heart failure. The risk of both coronary heart disease and congestive heart failure was highest for patients with levels in the highest tertile for all three markers; these patients had a two- to three-fold increase in coronary disease and heart failure compared with patients with no levels in the highest tertile. These differences also remained significant in adjusted analyses. |
| Makita et al. [201]. Cross-sectional study including 2056 individuals with a mean age of 58.3 years (range of 25–86 years). All examined with CRP and carotid ultrasound. | An association between plaque score and increasing CRP levels was seen only for men (p < 0.01); this association remained significant after being adjusted for age and other risk factors. Intima–media complex thickness and arterial dilation showed significant associations with CRP only in univariate, but not in adjusted, analyses. |
| Hosford-Donovan et al. [202]. Cross-sectional study of 108 elderly women with a mean age of 67.5 years (range of 65–70 years). | Body mass index (BMI), waist circumference, systolic blood pressure (SBP) and diastolic blood pressure (DBP) were significantly higher in patients with high CRP above the median level. SBP and DBP remained significantly higher in the high-CRP group after adjusting for BMI and use of antihypertensive medication. The influence of CRP on SBP was attenuated when adjusted for waist circumference (p = 0.062). Serum derived from high-CRP patients decreased the proliferation and the capillary tube length of in vitro cultured endothelial cells. |
| Labonté et al. [203]. Cross-sectional study of 801 Inuits (mean age of 36.3 years, range of 18–74 years). | Increased plasma CRP levels >2.0 mg/L were more prevalent among women. SBP was significantly and independently associated with increased CRP levels. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruserud, Ø.; Vo, A.K.; Rekvam, H. Hematopoiesis, Inflammation and Aging—The Biological Background and Clinical Impact of Anemia and Increased C-Reactive Protein Levels on Elderly Individuals. J. Clin. Med. 2022, 11, 706. https://doi.org/10.3390/jcm11030706
Bruserud Ø, Vo AK, Rekvam H. Hematopoiesis, Inflammation and Aging—The Biological Background and Clinical Impact of Anemia and Increased C-Reactive Protein Levels on Elderly Individuals. Journal of Clinical Medicine. 2022; 11(3):706. https://doi.org/10.3390/jcm11030706
Chicago/Turabian StyleBruserud, Øystein, Anh Khoi Vo, and Håkon Rekvam. 2022. "Hematopoiesis, Inflammation and Aging—The Biological Background and Clinical Impact of Anemia and Increased C-Reactive Protein Levels on Elderly Individuals" Journal of Clinical Medicine 11, no. 3: 706. https://doi.org/10.3390/jcm11030706