
Differential Recovery Patterns of the Maxilla and Mandible after Eliminating Nasal Obstruction in Growing Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animal Model
2.2. Morphological Evaluation of Maxillofacial Bones
2.3. Tissue Preparation
2.4. Histomorphometry with Toluidine Blue Staining
2.5. Immunohistochemistry for HIF-1α, OPG, and RANKL
2.6. Statistical Analysis
3. Results
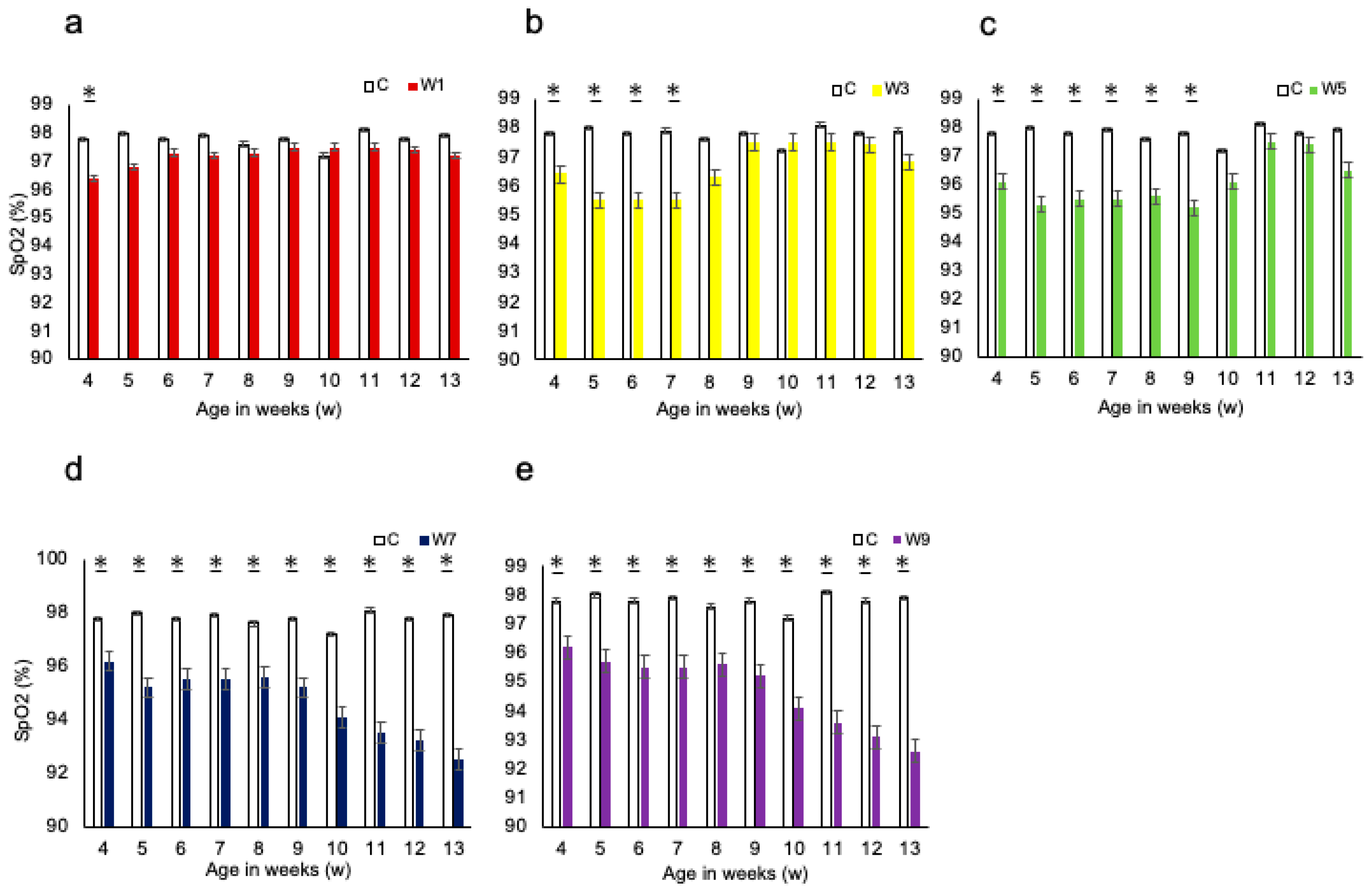
3.1. Systemic Changes
3.2. Measurement of the Maxillofacial and UA Morphology
3.3. Histomorphometry with Toluidine Blue Staining
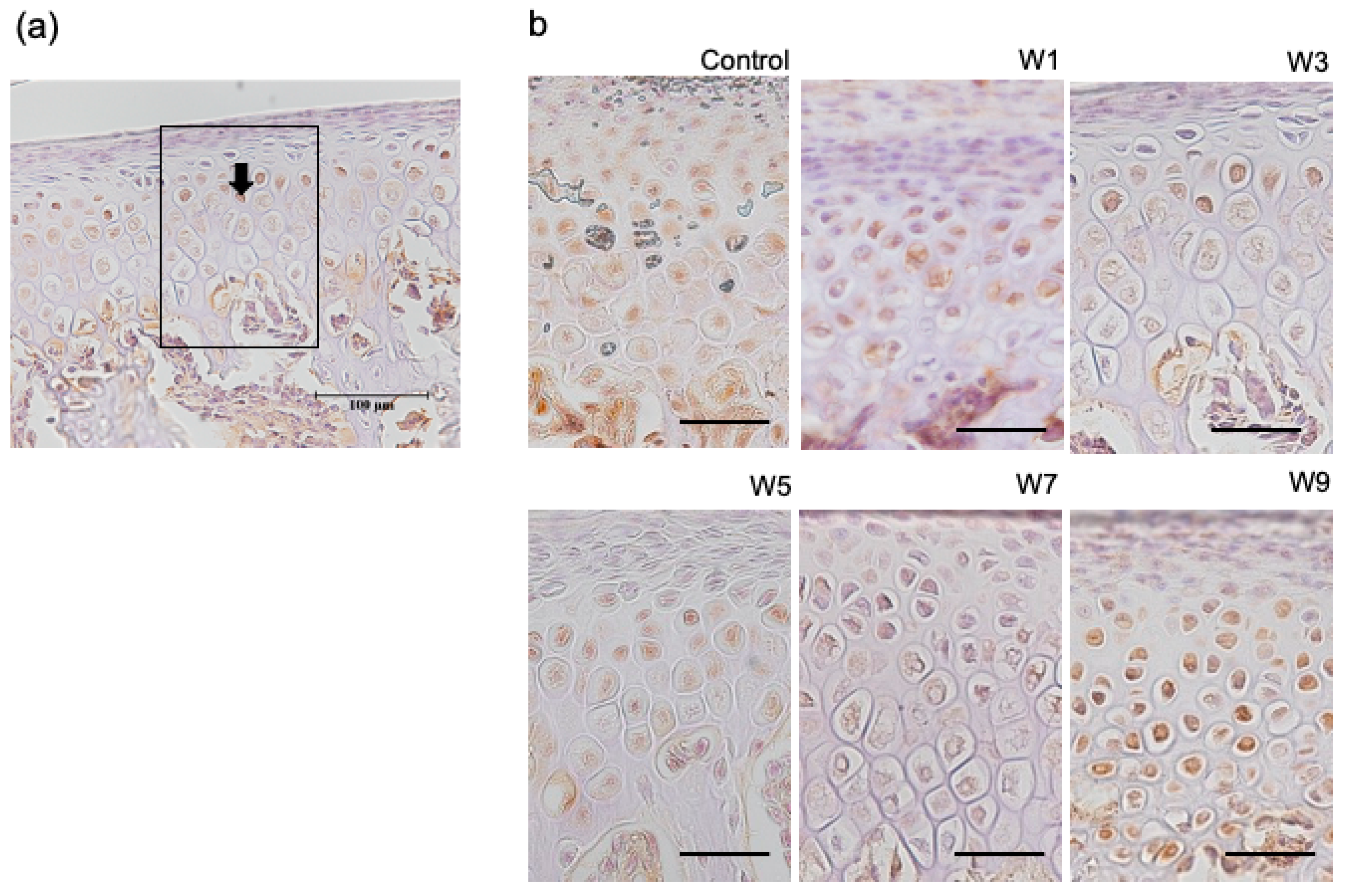
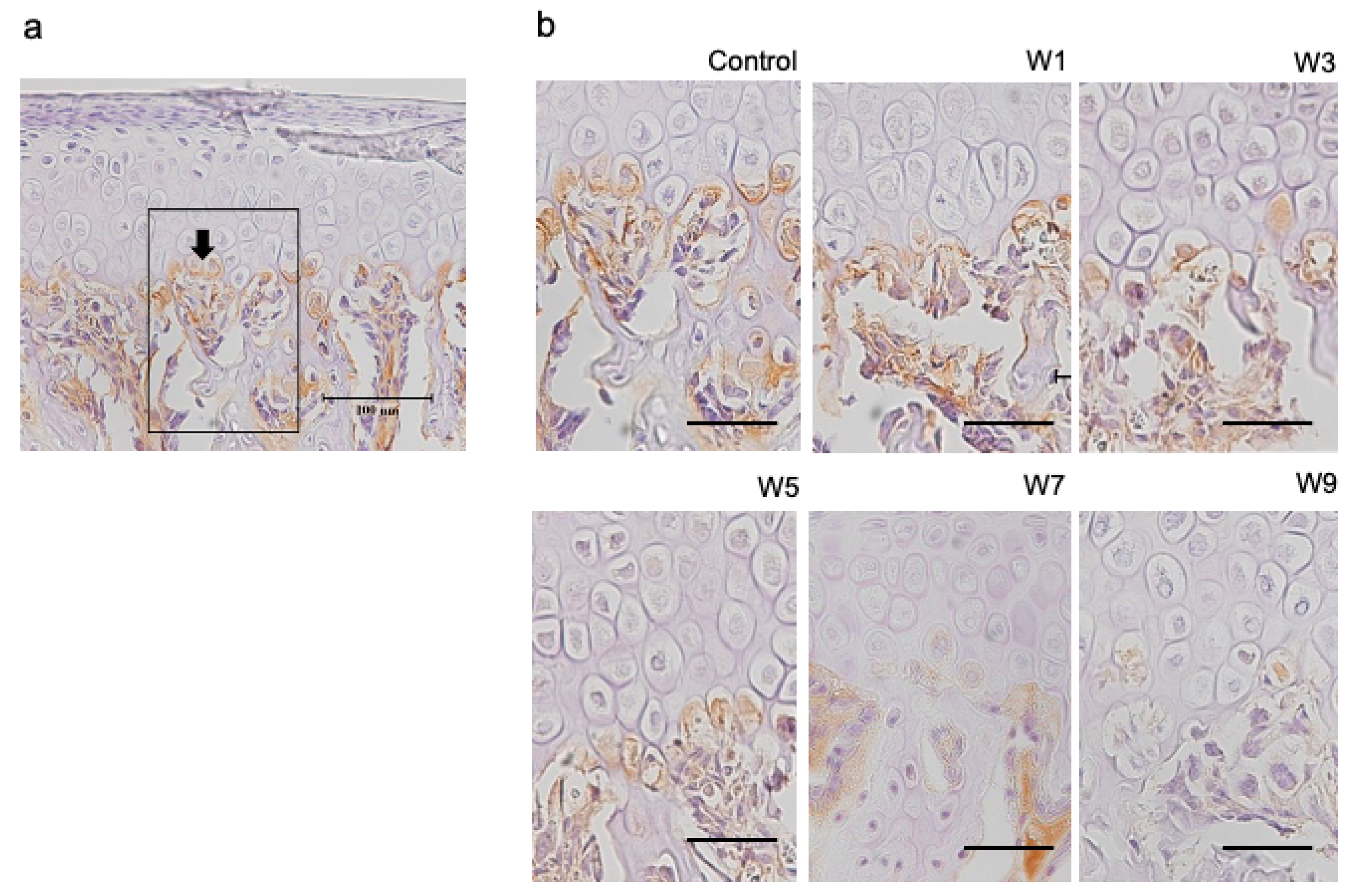
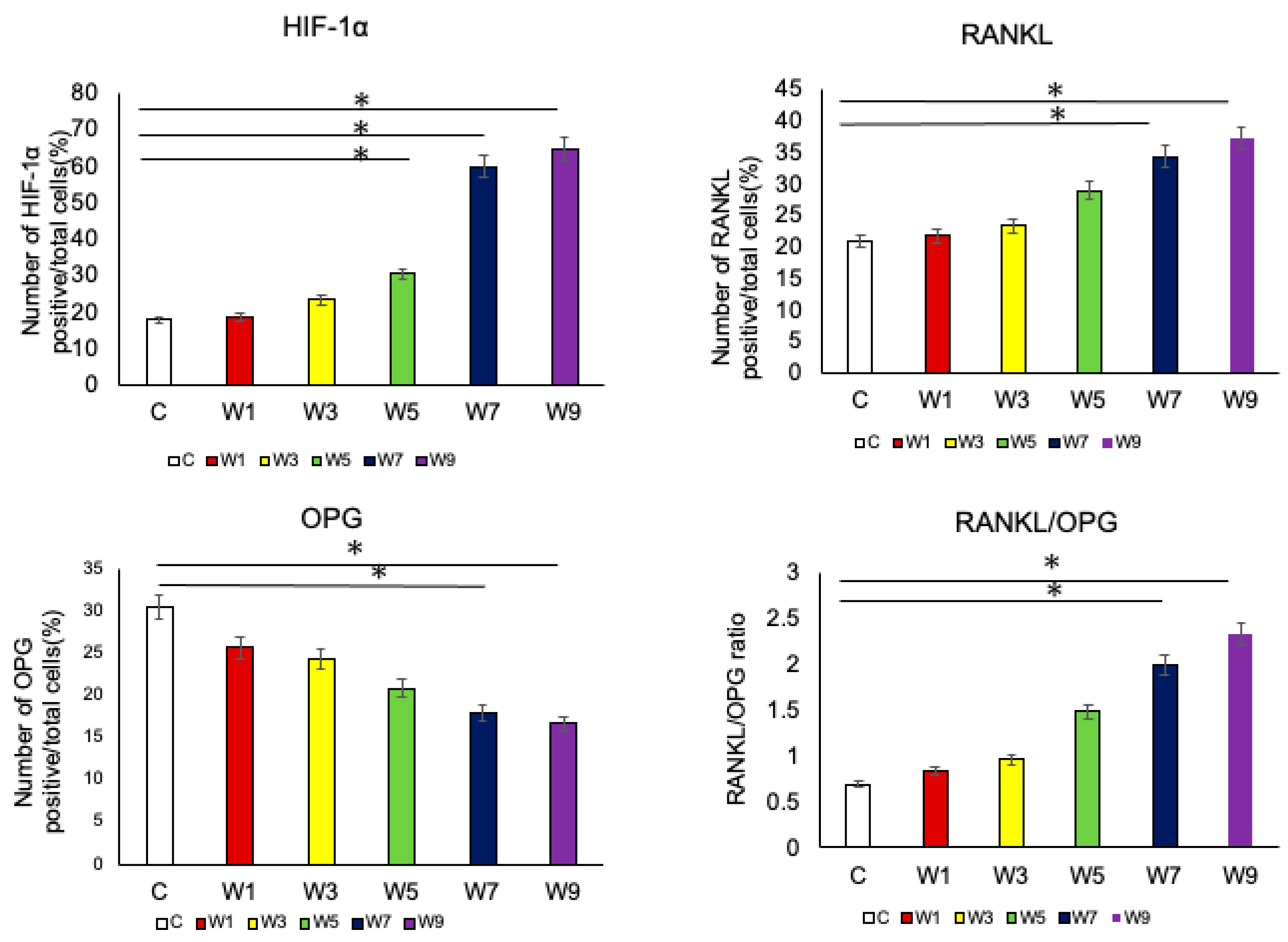
3.4. Expression of HIF-1α, OPG, and RANKL Protein in the Articular Cartilage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Udaka, T.; Suzuki, H.; Kitamura, T.; Shiomori, T.; Hiraki, N.; Fujimura, T.; Ueda, N. Relationships among nasal obstruction, daytime sleepiness, and quality of life. Laryngoscope 2006, 116, 2129–2132. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.S.; Book, D.T.; Burzynski, M.; Smith, T.L. Quality of life assessment in nasal airway obstruction. Laryngoscope 2003, 113, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Okihara, H.; Kokai, S.; Abe, Y.; Karin Harumi, U.K.; Makiguchi, M.; Kato, C.; Yabushita, T.; Michikawa, M.; Ono, T. NO during adolescence induces memory/learning impairments associated with BDNF/TrkB signaling pathway hypofunction and high corticosterone levels. J. Neurosci. Res. 2018, 96, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Ren, E.; Watari, I.; Jui-Chin, H.; Mizumachi-Kubono, M.; Podyma-Inoue, K.A.; Narukawa, M.; Misaka, T.; Watabe, T.; Ono, T. Unilateral airway obstruction alters sweet taste preference and sweet taste receptors in rat circumvallate papillae. Acta Histochem. 2019, 121, 135–142. [Google Scholar] [CrossRef]
- Posnick, J.C.; Agnihotri, N. Consequences and management of nasal airway obstruction in the dentofacial deformity patient. Curr. Opin. Otolaryngol. Head Neck Surg. 2010, 18, 323–331. [Google Scholar] [CrossRef]
- Williams, R.; Patel, V.; Chen, Y.F.; Tangbumrungtham, N.; Thamboo, A.; Most, S.P.; Nayak, J.V.; Liu, S.Y.C. The upper airway nasal complex: Structural contribution to persistent nasal obstruction. Otolaryngol. Head Neck Surg. 2019, 161, 171–177. [Google Scholar] [CrossRef]
- Zhao, Z.; Zheng, L.; Huang, X.; Li, C.; Liu, J.; Hu, Y. Effects of mouth breathing on facial skeletal development in children: A systematic review and meta-analysis. BMC Oral Health 2021, 21, 108. [Google Scholar] [CrossRef]
- Harari, D.; Redlich, M.; Miri, S.; Hamud, T.; Gross, M. The effect of mouth breathing versus nasal breathing on dentofacial and craniofacial development in orthodontic patients. Laryngoscope 2010, 120, 2089–2093. [Google Scholar] [CrossRef]
- Tang, H.; Yonemitsu, I.; Ikeda, Y.; Watakabe, K.; Shibata, S.; Hosomichi, J.; Ono, T. Effects of unilateral NO on the characteristics of jaw-closing muscles in growing rats. Angle Orthod. 2018, 89, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Watakabe, K.; Yonemitsu, I.; Ikeda, Y.; Tang, H.; Ono, T. Unilateral airway obstruction induces morphological changes of the mandibular condyle in growing. Orthod. Waves 2018, 77, 157–168. [Google Scholar] [CrossRef]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schipani, E.; Maes, C.; Carmeliet, G.; Semenza, G.L. Regulation of osteogenesis-angiogenesis coupling by HIFs and VEGF. J. Bone Miner. Res. 2009, 24, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H. Hypoxic regulation of osteoclast differentiation and bone resorption activity. Hypoxia 2015, 3, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Qiao, X.; Qi, S.; Zhang, X.; Li, S. Effect of adenoid hypertrophy on the upper airway and craniomaxillofacial region. Transl. Pediatr. 2021, 10, 2563–2572. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, J.; Tang, Y.; Wang, X.; Xue, X.; Sun, H.; Nie, P.; Qu, X.; Zhu, M. Dental arch dimensional changes after adenoidectomy or tonsillectomy in children with airway obstruction: A meta-analysis and systematic review under PRISMA guidelines. Medicine 2016, 95, e4976. [Google Scholar] [CrossRef] [PubMed]
- Schupper, A.J.; Nation, J.; Pransky, S. Adenoidectomy in children: What is the evidence and what is its role? Curr. Otorhinolaryngol. Rep. 2018, 6, 64–73. [Google Scholar] [CrossRef]
- Greenfeld, M.; Tauman, R.; DeRowe, A.; Sivan, Y. Obstructive sleep apnea syndrome due to adenotonsillar hypertrophy in infants. Int. J. Pediatr. Otorhinolaryngol. 2003, 67, 1055–1060. [Google Scholar] [CrossRef]
- Zettergren-Wijk, L.; Forsberg, C.M.; Linder-Aronson, S. Changes in dentofacial morphology after adeno-/tonsillectomy in young children with obstructive sleep apnoea—A 5-year follow-up study. Eur. J. Orthod. 2006, 28, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Becking, B.E.; Verweij, J.P.; Kalf-Scholte, S.M.; Valkenburg, C.; Bakker, E.W.P.; Merkesteyn, J.P.R. Impact of adenotonsillectomy on the dentofacial development of obstructed children: A systematic review and meta-analysis. Eur. J. Orthod. 2017, 39, 509–518. [Google Scholar] [CrossRef]
- Proffit, W.R.; Fields, H.W.; Larson, B.S.; Sarver, D.M. Contemporary Orthodontics, 3rd ed.; Mosby: St. Louis, MI, USA, 2000. [Google Scholar]
- Kusumoto, K.; Sato, K.; Mitani, H. The evaluation of the orthopedic appliances by using standard growth curves of maxilla and mandible. J. Jpn. Orthod. Soc. 1996, 55, 311–321. [Google Scholar]
- Ranly, D.M. Craniofacial growth. Dent. Clin. North Am. 2000, 44, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Yamaguchi, M.; Murakami, Y.; Horigome, Y.; Negishi, S.; Kasai, K. Changes in maxillofacial morphology due to improvement of nasal obstruction in rats. Orthod. Craniofac. Res. 2018, 21, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Yonemitsu, I.; Takei, M.; Shibata, S.; Ono, T. Mechanical loading leads to osteoarthritis-like changes in the hypofunctional temporomandibular joint in rats. Arch. Oral Biol. 2014, 59, 1368–1376. [Google Scholar] [CrossRef] [PubMed]
- Ishidori, H.; Okihara, H.; Ogawa, T.; Abe, Y.; Kato, C.; Phyo, T.A.; Fujita, A.; Kokai, S.; Ono, T. Nasal obstruction during the growth period modulates the Wnt/beta-catenin pathway and brain-derived neurotrophic factor production in association with tyrosine kinase receptor B mRNA reduction in mouse hippocampus. Eur. J. Neurosci. 2022, 55, 5–17. [Google Scholar] [CrossRef]
- Abe, Y.; Kato, C.; Uchima, K.; Okihara, H.; Ishida, T.; Fujita, K.; Yabushita, T.; Kokai, S.; Ono, T. Unilateral NO affects motor representation development within the face primary motor cortex in growing rats. J. Appl. Physol. 2017, 122, 1494–1503. [Google Scholar] [CrossRef] [Green Version]
- Padzys, G.S.; Martrette, J.M.; Tankosic, C.; Thornton, S.N.; Trabalon, M. Effects of short term forced oral breathing: Physiological changes and structural adaptation of diaphragm and orofacial muscles in rats. Arch. Oral Biol. 2011, 56, 1646–1654. [Google Scholar] [CrossRef]
- Padzys, G.S.; Tankosic, C.; Trabalon, M.; Martrette, J.M. Craniofacial development and physiological state after early oral breathing in rats. Eur. J. Oral. Sci. 2012, 120, 21–28. [Google Scholar] [CrossRef]
- Moss-Salentijn, L.; Melvin, L. Moss and the functional matrix. J. Dent. Res. 1997, 76, 1814–1817. [Google Scholar] [CrossRef]
- Kilic, N.; Oktay, H. Effects of rapid maxillary expansion on nasal breathing and some naso-respiratory and breathing problems in growing children: A literature review. Int. J. Pediatr. Otorhinolaryngol. 2008, 72, 1595–1601. [Google Scholar] [CrossRef]
- Saitoh, S.; Takahashi, I.; Mizoguchi, I.; Sasano, Y.; Kagayama, M.; Mitani, H. Compressive force promotes chondrogenic differentiation and hypertrophy in midpalatal suture cartilage in growing rats. Anat. Rec. 2000, 260, 392–401. [Google Scholar] [CrossRef]
- Abed, G.S.; Buschang, P.H.; Taylor, R.; Hinton, R.J. Maturational and functional related differences in rat craniofacial growth. Arch. Oral Biol. 2007, 52, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, H.; Zhu, Y.; Tang, Y.; Xue, X.; Nie, P.; Zhu, M.; Wang, B. Bilateral intermittent NO in adolescent rats leads to the growth defects of mandibular condyle. Arch. Oral Biol. 2019, 106, 104473. [Google Scholar] [CrossRef] [PubMed]
- Yonemitsu, I.; Muramoto, T.; Soma, K. The influence of masseter activity on rat mandibular growth. Arch. Oral Biol. 2007, 52, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Hashimoto, K.; Enomoto, S.; Shioda, S.; Nojyo, Y.; Sano, K. Maxillofacial deformity and change in the histochemical characteristics of the masseter muscle after unilateral sectioning of the facial nerve in growing rabbits. Acta Histochem. Cytochem. 2002, 35, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Kiliaridis, S. Masticatory muscle influence on craniofacial growth. Acta Odontol. Scand. 1995, 53, 196–202. [Google Scholar] [CrossRef]
- Hamrick, M.W.; McNeil, P.L.; Patterson, S.L. Role of muscle-derived growth factors in bone formation. J. Musculoskelet. Neuronal. Interact 2010, 10, 64–70. [Google Scholar]
- Yang, L.; Tsang, K.Y.; Tang, H.C.; Chan, D.; Cheah, K.S. Hypertrophic chondrocytes can become osteoblasts and osteocytes in endochondral bone formation. Proc. Natl. Acad. Sci. USA 2014, 111, 12097–12102. [Google Scholar] [CrossRef] [Green Version]
- Chung, U.I. Essential role of hypertrophic chondrocytes in endochondral bone development. Endocr. J. 2004, 51, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Leijten, J.C.; Moreira Teixeira, L.S.; Landman, E.B.; van Blitterswijk, C.A.; Karperien, M. Hypoxia inhibits hypertrophic differentiation and endochondral ossification in explanted tibiae. PLoS ONE 2012, 7, e49896. [Google Scholar] [CrossRef] [Green Version]
- Shirakura, M.; Tanimoto, K.; Eguchi, H.; Miyauchi, M.; Nakamura, H.; Hiyama, K.; Tanimoto, K.; Tanaka, E.; Takata, T.; Tanne, K. Activation of the hypoxia-inducible factor-1 in overloaded temporomandibular joint, and induction of osteoclastogenesis. Biochem. Biophys. Res. Commun. 2010, 393, 800–805. [Google Scholar] [CrossRef]
- Grimaud, E.; Soubigou, L.; Couillaud, S.; Coipeau, P.; Moreau, A.; Passuti, N.; Gouin, F.; Redini, F.; Heymann, D. Receptor activator of nuclear factor κB ligand (RANKL)/osteoprotegerin (OPG) ratio is increased in severe osteolysis. Am. J. Pathol. 2003, 163, 2021–2031. [Google Scholar] [CrossRef] [PubMed]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Wielockx, B.; Rauner, M.; Bozec, A. Hypoxia-inducible factors regulate osteoclasts in health and disease. Front. Cell Dev. Biol. 2021, 9, 658893. [Google Scholar] [CrossRef] [PubMed]
- Corso, P.F.C.L.; Meger, M.N.; Petean, I.B.F.; Souza, J.F.; Brancher, J.A.; da Silva, L.A.B.; Rebelatto, N.L.B.; Kluppel, L.E.; Sousa-Neto, M.D.; Küchler, E.C.; et al. Examination of OPG, RANK, RANKL andHIF-1α polymorphisms in temporomandibular joint ankylosis patients. J. Craniomaxillofac. Surg. 2019, 47, 766–770. [Google Scholar] [CrossRef]
- Zhu, J.; Tang, Y.; Wu, Q.; Ji, Y.C.; Feng, Z.F.; Kang, F.W. HIF-1α facilitates osteocyte-mediated osteoclastogenesis by activating JAK2/STAT3 pathway in vitro. J. Cell. Physiol. 2019, 234, 21182–21192. [Google Scholar] [CrossRef]
- Vandeberg, J.R.; Buschang, P.H.; Hinton, R.J. Craniofacial growth in growth hormone-deficient rats. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 278, 561–570. [Google Scholar] [CrossRef]
- Ochoa, B.K.; Nanda, R.S. Comparison of maxillary and mandibular growth. Am. J. Orthod. Dentofacial. Orthop. 2004, 125, 148–159. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landmarks | Definition |
|---|---|
| E | The intersection between the frontal bone and the most superior and anterior point of the ethmoid |
| U | The intersection between the maxillary sinus and the distal surface of the third superior molar tooth |
| Co | The most posterior and superior point on the mandibular condyle (Co, right; Co2, left) |
| Go | The most posterior point on the mandibular ramus |
| Mn | The most concave portion of the concavity on the inferior border of the mandibular corpus |
| Gn | The most inferior point on the ramus that lies on a perpendicular bisector of the line Go–Mn |
| Me | The most inferior and anterior point of the lower border of the mandible |
| L1 | The most anterior and superior point on the alveolar bone of the mandibular incisor |
| Mi | The junction of the alveolar bone and the mesial surface of the first mandibular molar |
| ANS | The most anterior part of the palate |
| PNS | The most posterior part of the palate |
| X1 | The most anterior and superior point in the molar process of the right maxilla |
| X2 | The most anterior and superior point in the molar process of the left maxilla |
| Measurements | Interpretation |
|---|---|
| X1–X2 | Maxillary width |
| ANS–PNS | Maxillary length |
| U–E | Maxillary height |
| Co–L1 | Total mandibular length |
| Mi–L1 | Anterior mandibular length |
| Go–Mn | Posterior mandibular length |
| Co–Gn | Ramus height |
| Co–Me | Mandibular body length |
| Co1–Co2 | Mandibular width |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keitoku, M.; Yonemitsu, I.; Ikeda, Y.; Tang, H.; Ono, T. Differential Recovery Patterns of the Maxilla and Mandible after Eliminating Nasal Obstruction in Growing Rats. J. Clin. Med. 2022, 11, 7359. https://doi.org/10.3390/jcm11247359
Keitoku M, Yonemitsu I, Ikeda Y, Tang H, Ono T. Differential Recovery Patterns of the Maxilla and Mandible after Eliminating Nasal Obstruction in Growing Rats. Journal of Clinical Medicine. 2022; 11(24):7359. https://doi.org/10.3390/jcm11247359
Chicago/Turabian StyleKeitoku, Mirei, Ikuo Yonemitsu, Yuhei Ikeda, Huan Tang, and Takashi Ono. 2022. "Differential Recovery Patterns of the Maxilla and Mandible after Eliminating Nasal Obstruction in Growing Rats" Journal of Clinical Medicine 11, no. 24: 7359. https://doi.org/10.3390/jcm11247359