

Association of Transferrin Gene Polymorphism with Cognitive Deficits and Psychiatric Symptoms in Patients with Chronic Schizophrenia
 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Clinical Examination and Cognitive Assessments
2.3. DNA Extraction and SNP Genotyping
2.4. Statistical Analysis
3. Results
3.1. Clinical Data, and Allele and Genotype Frequencies
3.2. Genotypic Effects on Cognitive Functions between Patients and Controls
3.3. Relationship between the rs3811655 Genotypes and Clinical Characteristics in Patients with Schizophrenia
3.4. Stepwise Multiple Regression Predicting Psychiatric Symptoms from Transferrin Gene Polymorphism, Cognitive Measures, and Demographic Measures among Patients with Schizophrenia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lange, K.W.; Kornhuber, J.; Kruzik, P.; Rausch, W.D.; Gabriel, E.; Jellinger, K.; Riederer, P. Brain iron and schizophrenia. In Iron in Central Nervous System Disorders; Springer: Vienne, Autriche, 1993; pp. 37–43. [Google Scholar]
- Peralta, V.; Cuesta, M.J.; Mata, I.; Serrano, J.F.; Perez-Nievas, F.; Natividad, M.C. Serum iron in catatonic and noncatatonic psychotic patients. Biol. Psychiatry 1999, 45, 788–790. [Google Scholar] [CrossRef]
- Xu, M.; Guo, Y.; Cheng, J.; Xue, K.; Yang, M.; Song, X.; Feng, Y.; Cheng, J. Brain Iron Assessment in Patients with First-episode Schizophrenia using Quantitative Susceptibility Mapping. NeuroImage Clin. 2021, 31, 102736. [Google Scholar] [CrossRef] [PubMed]
- Yanik, M.; Kocyigit, A.; Tutkun, H.; Vural, H.; Herken, H. Plasma manganese, selenium, zinc, copper, and iron concentrations in patients with schizophrenia. Biol. Trace Elem. Res. 2004, 98, 109–118. [Google Scholar] [CrossRef]
- Stankiewicz, J.; Panter, S.S.; Neema, M.; Arora, A.; Batt, C.E.; Bakshi, R. Iron in chronic brain disorders: Imaging and neurotherapeutic implications. Neurotherapeutics 2007, 4, 371–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Insel, B.J.; Schaefer, C.A.; McKeague, I.W.; Susser, E.S.; Brown, A.S. Maternal iron deficiency and the risk of schizophrenia in offspring. Arch. Gen. Psychiatry 2008, 65, 1136–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, A.M.; Rao, R.B. Perinatal iron deficiency as an early risk factor for schizophrenia. Nutr. Neurosci. 2021, 25, 2218–2227. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, H.J.; Nielsen, P.R.; Pedersen, C.B.; Mortensen, P.B. Association between prepartum maternal iron deficiency and offspring risk of schizophrenia: Population-based cohort study with linkage of Danish national registers. Schizophr. Bull. 2011, 37, 982–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambling, L.; Charania, Z.; Hannah, L.; Antipatis, C.; Lea, R.G.; McArdle, H.J. Effect of iron deficiency on placental cytokine expression and fetal growth in the pregnant rat. Biol. Reprod. 2002, 66, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, J.L.; Felt, B.; Schallert, T.; Burhans, M.; Connor, J.R.; Georgieff, M.K. Moderate iron deficiency in infancy: Biology and behavior in young rats. Behav. Brain Res. 2006, 170, 224–232. [Google Scholar] [CrossRef]
- Eyles, D.W.; Dean, A.J. Maternal nutritional deficiencies and schizophrenia: Lessons from animal models with a focus on developmental vitamin D deficiency. In Handbook of Behavioral Neuroscience; Elsevier: Amsterdam, The Netherlands, 2016; pp. 243–264. [Google Scholar]
- McGrath, J.; Brown, A.; St Clair, D. Prevention and schizophrenia—The role of dietary factors. Schizophr. Bull. 2011, 37, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahanshad, N.; Rajagopalan, P.; Thompson, P.M. Neuroimaging, nutrition, and iron-related genes. Cell. Mol. Life Sci. 2013, 70, 4449–4461. [Google Scholar] [CrossRef] [Green Version]
- Bourque, S.L.; Iqbal, U.; Reynolds, J.N.; Adams, M.A.; Nakatsu, K. Perinatal iron deficiency affects locomotor behavior and water maze performance in adult male and female rats. J. Nutr. 2008, 138, 931–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisansky, M.T.; Wickham, R.J.; Su, J.; Fretham, S.; Yuan, L.-L.; Sun, M.; Gewirtz, J.C.; Georgieff, M.K. Iron deficiency with or without anemia impairs prepulse inhibition of the startle reflex. Hippocampus 2013, 23, 952–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhukovskaya, E.; Karelin, A.; Rumyantsev, A. Neurocognitive Dysfunctions in Iron Deficiency Patients. Iron Defic. Anemia 2019. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Transferrin and transferrin receptor function in brain barrier systems. Cell. Mol. Neurobiol. 2000, 20, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Wiser, M.; Levkowitch, Y.; Neuman, M.; Yehuda, S.; Wiser, M. Decrease of serum iron in acutely psychotic schizophrenic patients. Int. J. Neurosci. 1994, 78, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Devi, P.U.; Chinnaswamy, P.; Murugan, S.; Selvi, S. Plasma levels of trace elements in patients with different symptoms of Schizophrenia. Biosci. Biotechnol. Res. Asia 2016, 5, 261–268. [Google Scholar]
- Sienaert, P.A.; Dhossche, D.M.; Vancampfort, D.; De Hert, M.; Gazdag, G. A clinical review of the treatment of catatonia. Front. Psychiatry 2014, 5, 181. [Google Scholar] [CrossRef]
- Seidman, L.J.; Giuliano, A.J.; Meyer, E.C.; Addington, J.; Cadenhead, K.S.; Cannon, T.D.; North American Prodrome Longitudinal Study (NAPLS) Group. Neuropsychology of the prodrome to psychosis in the NAPLS consortium: Relationship to family history and conversion to psychosis. Arch. Gen. Psychiatry 2010, 67, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, M.; Abdallah, M.; Swendsen, J.; N’Kaoua, B.; Chanraud, S.; Schweitzer, P.; Fatseas, M.; Serre, F.; Barse, E.; Auriacombe, M.; et al. Real-time cognitive performance and positive symptom expression in schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Galderisi, S.; Mucci, A.; Buchanan, R.W.; Arango, C. Negative symptoms of schizophrenia: New developments and unanswered research questions. Lancet Psychiatry 2018, 5, 664–677. [Google Scholar] [CrossRef]
- Pelletier-Baldelli, A.; Holt, D.J. Are negative symptoms merely the “real world” consequences of deficits in social cognition? Schizophr. Bull. 2020, 46, 236–241. [Google Scholar] [CrossRef]
- Leitner, D.F.; Connor, J.R. Functional roles of transferrin in the brain. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 393–402. [Google Scholar] [CrossRef]
- Owiredu, W.; Brenya, P.K.; Osei, Y.; Laing, E.F.; Okrah, C.O.; Obirikorang, C.; Donkor, S. Evaluation of serum iron overload, AST: ALT ratio and log 10 ferritin: AST ratio among schizophrenia patients in the Kumasi Metropolis, Ghana: A case-control study. BMC Res. Notes 2019, 12, 802. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.; Saha, N.; Tsoi, W. Serum iron, total iron-binding capacity, transferrin and haptoglobin concentration in schizophrenic patients. Age 1985, 24, 214. [Google Scholar]
- Kuloglu, M.; Atmaca, M.; Üstündag, B.; Canatan, H.; Gecici, O.; Tezcan, E. Serum iron levels in schizophrenic patients with or without akathisia. Eur. Neuropsychopharmacol. 2003, 13, 67–71. [Google Scholar] [CrossRef]
- Gemmati, D.; Zeri, G.; Orioli, E.E.; De Gaetano, F.; Salvi, F.; Bartolomei, I.; D’Alfonso, S.; Dall’Osso, C.; A Leone, M.; Singh, A.V.; et al. Polymorphisms in the genes coding for iron binding and transporting proteins are associated with disability, severity, and early progression in multiple sclerosis. BMC Med. Genet. 2012, 13, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Yui, K.; Imataka, G.; Nakamura, H.; Ohara, N.; Naito, Y. Eicosanoids derived from arachidonic acid and their family prostaglandins and cyclooxygenase in psychiatric disorders. Curr. Neuropharmacol. 2015, 13, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Hakak, Y.; Walker, J.R.; Li, C.; Wong, W.H.; Davis, K.L.; Buxbaum, J.D.; Fienberg, A.A. Genome-wide expression analysis reveals dysregulation of myelination-related genes in chronic schizophrenia. Proc. Natl. Acad. Sci. USA 2001, 98, 4746–4751. [Google Scholar] [CrossRef] [Green Version]
- Davis, K.L.; Davis, K.L.; Stewart, D.G.; Friedman, J.I.; Buchsbaum, M.; Harvey, P.D.; Hof, P.R.; Haroutunian, V. White matter changes in schizophrenia: Evidence for myelin-related dysfunction. Arch. Gen. Psychiatry 2003, 60, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Yue, W.; Tang, F.; Wang, L.; Han, Y.; Zhang, D. Polymorphisms of Transferrin gene are associated with schizophrenia in Chinese Han population. J. Psychiatr. Res. 2008, 42, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Buretić-Tomljanović, A.; Vraneković, J.; Rubeša, G.; Jonovska, S.; Tomljanović, D.; Šendula-Jengić, V.; Ristić, S. HFE mutations and transferrin C1/C2 polymorphism among Croatian patients with schizophrenia and schizoaffective disorder. Mol. Biol. Rep. 2012, 39, 2253–2258. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wang, D.; Xiu, M.; Chen, D.; Lackey, B.; Wu, H.E.; Zhou, J.; Zhang, X. Polymorphism of Transferrin Gene Impacts the Mediating Effects of Psychotic Symptoms on the Relationship between Oxidative Stress and Cognition in Patients with Chronic Schizophrenia. Antioxidants 2022, 11, 125. [Google Scholar] [CrossRef]
- Sponheim, S.; Jung, R.; Seidman, L.; Mesholam-Gately, R.; Manoach, D.; O’Leary, D.; Ho, B.-C.; Andreasen, N.; Lauriello, J.; Schulz, S. Cognitive deficits in recent-onset and chronic schizophrenia. J. Psychiatr. Res. 2010, 44, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Woods, S.W. Chlorpromazine equivalent doses for the newer atypical antipsychotics. J. Clin. Psychiatry 2003, 64, 663–667. [Google Scholar] [CrossRef]
- Randolph, C.; Tierney, M.C.; Mohr, E.; Chase, T.N. The Repeatable Battery for the Assessment of Neuropsychological Status (RBANS): Preliminary clinical validity. J. Clin. Exp. Neuropsychol. 1998, 20, 310–319. [Google Scholar] [CrossRef]
- MWer, S.; Dykes, D.; Polesky, H. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res 1988, 16, 1215. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Maeno, N.; Takahashi, N.; Saito, S.; Ji, X.; Branko, A.; Ishihara, R.; Yoshida, K.; Inada, T.; Iidaka, T.; Ozaki, N. Association study between the transferrin gene and schizophrenia in the Japanese population. Neuroreport 2007, 18, 517–520. [Google Scholar] [CrossRef]
- Fananas, L.; Moral, P.; Tusquets, J.M.; Bertranpetit, J. Genetic markers in schizophrenia: ACP1, ESD, TF and GC polymorphisms. Hum. Hered. 1990, 40, 136–140. [Google Scholar] [PubMed]
- Jahanshad, N.; Kohannim, O.; Hibar, D.P.; Stein, J.L.; McMahon, K.L.; de Zubicaray, G.I.; Medland, S.E.; Montgomery, G.W.; Whitfield, J.B.; Martin, N.G.; et al. Brain structure in healthy adults is related to serum transferrin and the H63D polymorphism in the HFE gene. Proc. Natl. Acad. Sci. USA 2012, 109, E851–E859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.; Neves, P.; Gozzelino, R. Multilevel impacts of Iron in the brain: The cross talk between neurophysiological mechanisms, cognition, and social behavior. Pharmaceuticals 2019, 12, 126. [Google Scholar] [CrossRef] [Green Version]
- Damulina, A.; Pirpamer, L.; Soellradl, M.; Sackl, M.; Tinauer, C.; Hofer, E.; Enzinger, C.; Gesierich, B.; Duering, M.; Ropele, S.; et al. Cross-sectional and longitudinal assessment of brain iron level in Alzheimer disease using 3-T MRI. Radiology 2020, 296, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Bender, S.; Weisbrod, M.; Resch, F. Which perspectives can endophenotypes and biological markers offer in the early recognition of schizophrenia? J. Neural Transm. 2007, 114, 1199–1215. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J.; Fone, K.; Steckler, T.; Horan, W.P. Negative symptoms of schizophrenia: Clinical characteristics, pathophysiological substrates, experimental models and prospects for improved treatment. Eur. Neuropsychopharmacol. 2014, 24, 645–692. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-W.; Stewart, R.; Park, W.Y.; Jhon, M.; Lee, J.Y.; Kim, S.Y.; Yoon, J.S. Latent iron deficiency as a marker of negative symptoms in patients with first-episode schizophrenia spectrum disorder. Nutrients 2018, 10, 1707. [Google Scholar] [CrossRef] [Green Version]
- Matak, P.; Matak, A.; Moustafa, S.; Aryal, D.K.; Benner, E.J.; Wetsel, W.; Andrews, N.C. Disrupted iron homeostasis causes dopaminergic neurodegeneration in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 3428–3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daubner, S.C.; Le, L.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Unger, E.L.; Wiesinger, J.A.; Hao, L.; Beard, J.L. Dopamine D2 Receptor Expression Is Altered by Changes in Cellular Iron Levels in PC12 Cells and Rat Brain Tissue. J. Nutr. 2008, 138, 2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benyamin, B.; McRae, A.F.; Zhu, G.; Gordon, S.; Henders, A.K.; Palotie, A.; Peltonen, L.; Martin, N.G.; Montgomery, G.W.; Whitfield, J.B.; et al. Variants in TF and HFE explain∼40% of genetic variation in serum-transferrin levels. Am. J. Hum. Genet. 2009, 84, 60–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali-Rahmani, F.; Grigson, P.S.; Lee, S.; Neely, E.; Connor, J.R.; Schengrund, C.-L. H63D mutation in hemochromatosis alters cholesterol metabolism and induces memory impairment. Neurobiol. Aging 2014, 35, 1511.e1–1511.e12. [Google Scholar] [CrossRef] [PubMed]


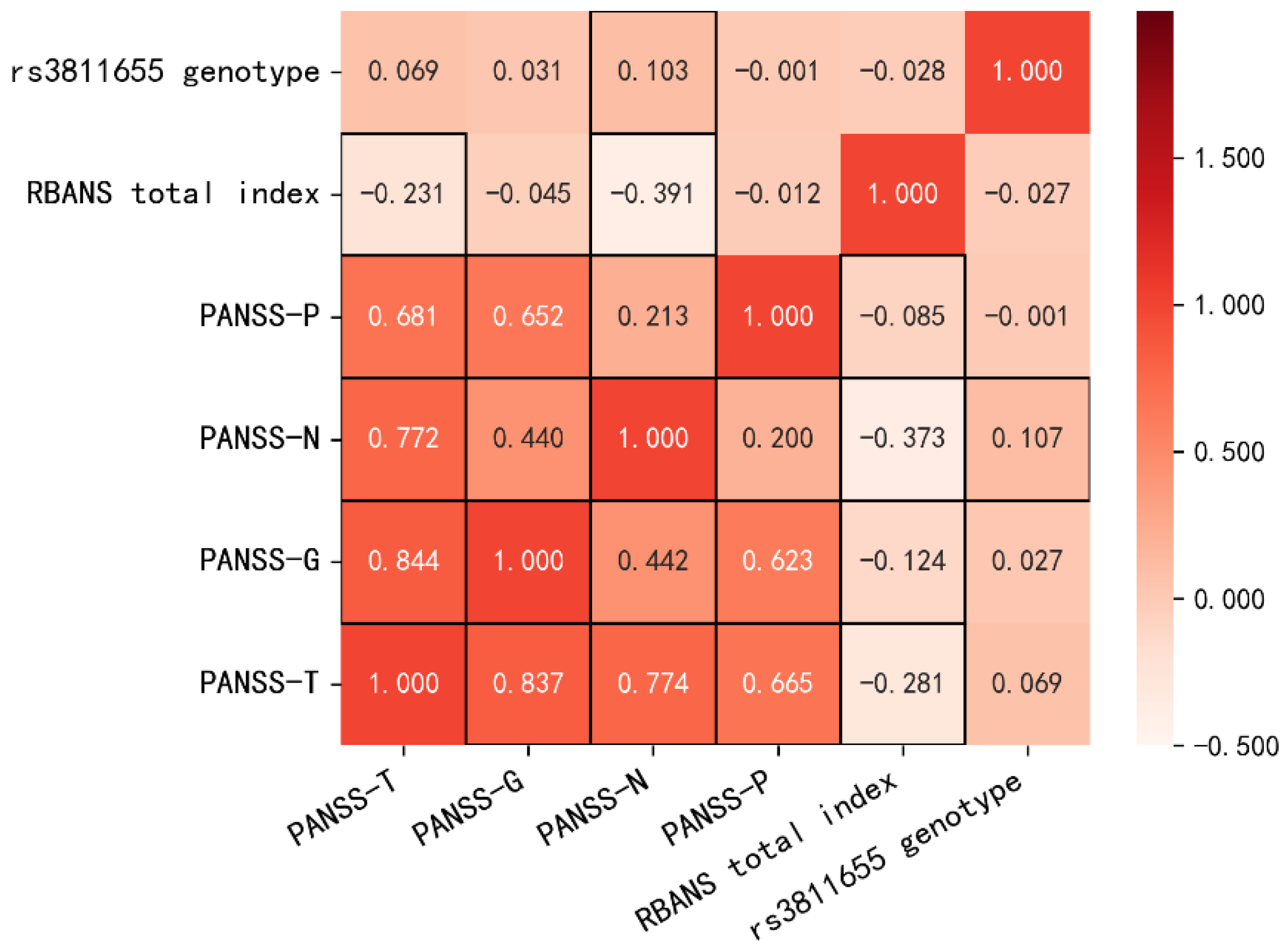{kind=link}
| Schizophrenia Patients | Healthy Controls | FGenotypic Effects (p) | FCase vs. Controls (p) | FIntreaction Effects (p) | |||||
|---|---|---|---|---|---|---|---|---|---|
| RBANS Scores | C/C (n = 350) | G/C (n = 186) | G/G (n = 28) | C/C (n = 256) | G/C (n = 148) | G/G (n = 18) | |||
| Immediate memory | 58.6 ± 16.2 | 59.2 ± 16.5 | 54.8 ± 13.9 | 74.0 ± 16.4 | 75.7 ± 17.8 | 70.1 ± 13.2 | 1.86 (0.16) | 46.78 (<0.001) | 0.46 (0.63) |
| Visuospatial | 77.6 ± 18.8 | 77.7 ± 18.2 | 73.1 ± 20.8 | 78.4 ± 14.1 | 79.4 ± 15.2 | 76.4 ± 15.0 | 0.90 (0.41) | 0.11 (0.74) | 0.37 (0.69) |
| Language | 81.7 ± 15.8 | 82.1 ± 14.4 | 76.8 ± 15.0 | 93.3 ± 12.4 | 94.5 ± 13.7 | 89.7 ± 10.6 | 2.41 (0.09) | 50.03 (<0.001) | 0.38 (0.69) |
| Attention | 71.4 ± 17.5 | 70.3 ± 17.4 | 61.4 ± 14.7 | 85.8 ± 18.8 | 87.3 ± 19.3 | 82.9 ± 21.5 | 2.57 (0.08) | 59.07 (<0.001) | 1.85 (0.16) |
| Delayed memory | 67.2 ± 19.0 | 65.6 ± 19.8 | 63.5 ± 19.1 | 85.0 ± 14.6 | 87.9 ± 14.4 | 81.4 ± 15.8 | 1.09 (0.34) | 74.01 (<0.001) | 2.37 (0.09) |
| Total index | 64.7 ± 15.1 | 64.5 ± 14.9 | 60.4 ± 12.7 | 78.4 ± 13.7 | 80.7 ± 15.0 | 73.3 ± 14.4 | 3.12 (<0.05) | 57.34 (<0.001) | 1.97 (0.14) |
| Genotype | F/χ2 (p) | |||
|---|---|---|---|---|
| C/C (n = 350) | G/C (n = 186) | G/G (n = 28) | ||
| Age of onset (years) | 23.1 ± 4.8 | 23.3 ± 4.8 | 24.4 ± 4.5 | 0.31 (0.73) |
| Duration of illness (years) | 24.6 ± 8.8 | 24.2 ± 8.8 | 26.5 ± 7.6 | 0.23 (0.80) |
| Daily antipsychotic dose (mg/day) (chlorpromazine equivalent) | 437.5 ± 389.7 | 426.8 ± 287.7 | 463.3 ± 443.0 | 0.17 (0.84) |
| HGB (g/dl) | 141.4 ± 21.6 | 139.1 ± 12.0 | 140.6 ± 13.8 | 0.06 (0.94) |
| PANSS scores | ||||
| P subscore | 11.1 ± 4.6 | 11.4 ± 5.0 | 11.0 ± 4.8 | 0.42 (0.66) |
| N subscore | 21.7 ± 7.2 | 23.0 ± 7.3 | 25.1 ± 7.4 | 4.27 (<0.05) |
| G subscore | 24.6 ± 5.2 | 25.2 ± 6.0 | 25.0 ± 4.9 | 0.81 (0.45) |
| Total score | 57.3 ± 13.0 | 59.7 ± 14.8 | 61.1 ± 11.5 | 2.27 (0.10) |
| Predictors | Stepwise Regression | Correlations | ||||
|---|---|---|---|---|---|---|
| B (95%CI) | SE B | β | p | Partial * | Part † | |
| Dependent Variable: PANSS-N | ||||||
| Constant | 34.18 (28.08 to 40.28) | 3.11 | 0.00 | |||
| Language | −0.11 (−0.15 to −0.06) | 0.02 | −0.23 | 0.00 | −0.21 | −0.18 |
| Delayed memory | −0.06 (−0.10 to −0.02) | 0.02 | −0.15 | 0.00 | −0.12 | −0.11 |
| rs3811655 genotype | 0.73 (0.17 to 1.29) | 0.29 | 0.09 | 0.01 | 0.11 | 0.09 |
| Dependent Variable: PANSS-T | ||||||
| Constant | 54.07 (42.09 to 66.04) | 6.10 | 0.00 | |||
| Daily antipsychotic dose | 0.00 (0.00 to 0.01) | 0.00 | 0.10 | 0.01 | 0.10 | 0.09 |
| Language | −0.13 (−0.21 to −0.04) | 0.04 | −0.14 | 0.00 | −0.13 | −0.11 |
| Delayed memory | −0.17 (−0.24 to −0.09) | 0.04 | −0.24 | 0.00 | −0.18 | −0.17 |
| rs3811655 genotype | 1.08 (−0.02 to 2.19) | 0.56 | 0.07 | 0.06 | 0.08 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.; Wang, D.; Xiu, M.; Chen, D.; Lackey, B.; Wu, H.E.; Wang, L.; Zhang, X. Association of Transferrin Gene Polymorphism with Cognitive Deficits and Psychiatric Symptoms in Patients with Chronic Schizophrenia. J. Clin. Med. 2022, 11, 6414. https://doi.org/10.3390/jcm11216414
Chen P, Wang D, Xiu M, Chen D, Lackey B, Wu HE, Wang L, Zhang X. Association of Transferrin Gene Polymorphism with Cognitive Deficits and Psychiatric Symptoms in Patients with Chronic Schizophrenia. Journal of Clinical Medicine. 2022; 11(21):6414. https://doi.org/10.3390/jcm11216414
Chicago/Turabian StyleChen, Pinhong, Dongmei Wang, Meihong Xiu, Dachun Chen, Blake Lackey, Hanjing E. Wu, Lubin Wang, and Xiangyang Zhang. 2022. "Association of Transferrin Gene Polymorphism with Cognitive Deficits and Psychiatric Symptoms in Patients with Chronic Schizophrenia" Journal of Clinical Medicine 11, no. 21: 6414. https://doi.org/10.3390/jcm11216414