
microRNAs in Subarachnoid Hemorrhage (Review of Literature)


Abstract
:1. Subarachnoid Hemorrhage
1.1. Introduction
1.2. Epidemiology
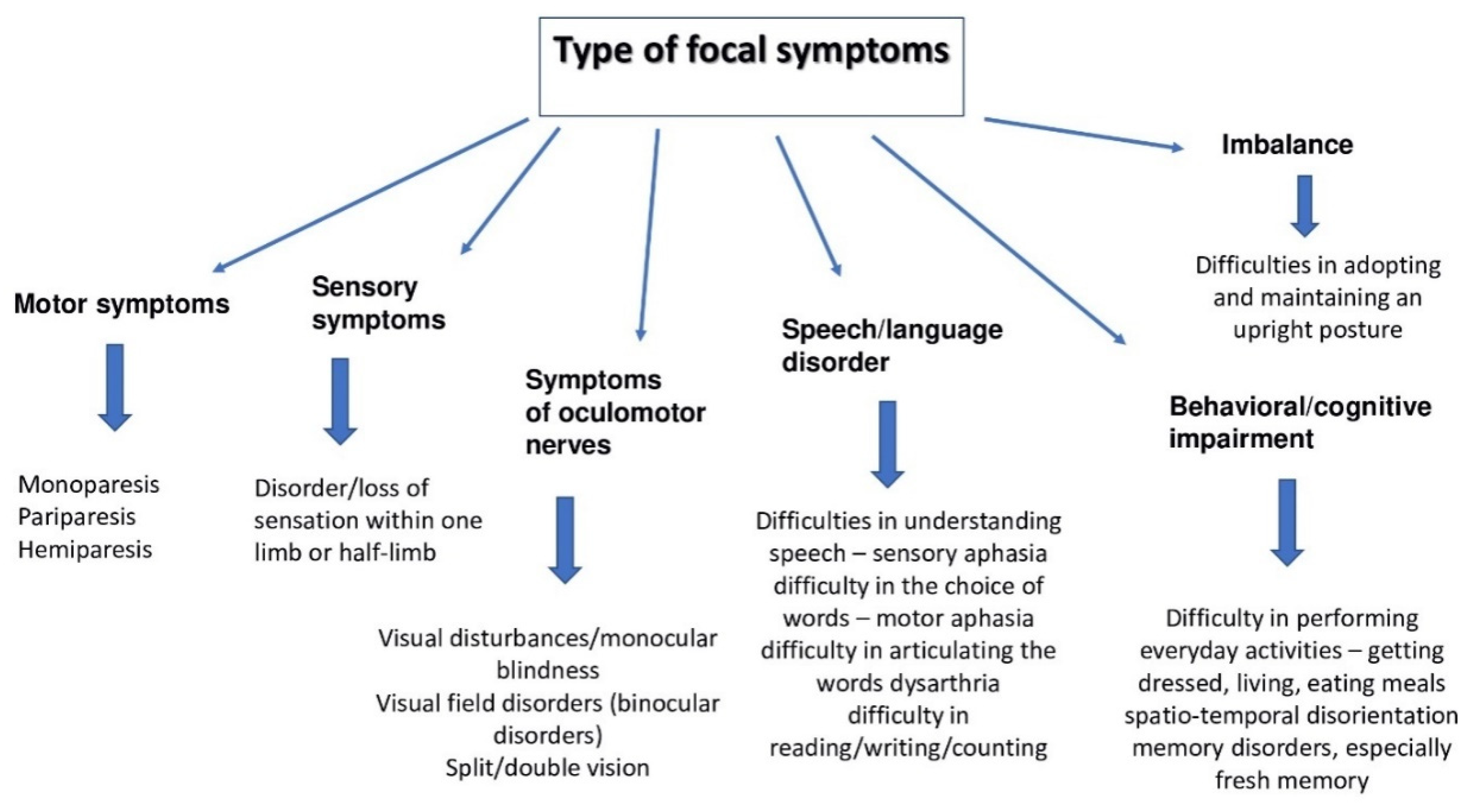
1.3. Symptoms
- -
- Sudden, severe headache;
- -
- Meningeal symptoms;
- -
- Nausea and vomiting—in a retrospective review of 109 patients, Fontanarosa et al. reported that 77% of patients with aSAH had nausea or vomiting [21];
- -
- Photophobia;
- -
- Disturbances of consciousness up to and including loss of consciousness;
- -
- Neurological deficits;
- -
- Seizures of an epileptic nature [5].
1.4. Differential Diagnosis
- -
- Venous cerebral thrombosis—a disease whose predominant symptoms are headache, impaired consciousness, and swelling of the optic nerve disc; focal symptoms with various motor and sensory deficits and defects, as well as speech and vision disorders may also occur [23];
- -
- Meningitis—the main symptoms are slowly developing headache, fever, neck stiffness, and impaired consciousness; vomiting, convulsions, photophobia, hypersensitivity to sounds may also occur [24];
- -
- Migraine—manifested by a severe half headache; it can occur with an aura or without. The peak of the disease is observed in the age group of 30–50 years [25].
1.5. Interview
1.6. Diagnosis and Treatment
- -
- To avoid this type of complication, the 3H rule is used:
- -
- Hypervolemia—patients with subarachnoid hemorrhage tend to fall into hypovolemia, associated with improper cerebral flow;
- -
- Maintaining increased blood pressure (hypertension);
- -
- Hemodilution [30].
2. microRNAs
miRNA Biogenesis
3. miRNA in Neurological Disorders
4. Circulating miRNAs in SAH
5. Exosomal miRNAs in aSAH
6. The Clinical Significance of miRNA—Future Perspective and Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muehlschlegel, S. Subarachnoid Hemorrhage. Contin. MinneapMinn 2018, 24, 1623–1657. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.H. Subarachnoid Hemorrhage. Contin. MinneapMinn 2021, 27, 1201–1245. [Google Scholar] [CrossRef] [PubMed]
- Lawton, M.T.; Vates, G.E. Subarachnoid Hemorrhage. N. Engl. J. Med. 2017, 377, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.S.; Kim, J.E.; Park, S.Q.; Ko, J.K.; Kim, D.W.; Park, J.C.; Yeon, J.Y.; Chung, S.Y.; Chung, J.; Joo, S.P.; et al. Korean Clinical Practice Guidelines for Aneurysmal Subarachnoid Hemorrhage. J. Korean Neurosurg. Soc. 2018, 61, 127–166. [Google Scholar] [CrossRef] [Green Version]
- Petridis, A.K.; Kamp, M.A.; Cornelius, J.F.; Beez, T.; Beseoglu, K.; Turowski, B.; Steiger, H.J. Aneurysmal Subarachnoid Hemorrhage. Dtsch. Arztebl. Int. 2017, 114, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Brinjikji, W.; Zhu, Y.Q.; Lanzino, G.; Cloft, H.J.; Murad, M.H.; Wang, Z.; Kallmes, D.F. Factors for Growth of Intracranial Aneurysms: A Systematic Review and Meta-Analysis. AJNR Am. J. Neuroradiol. 2016, 37, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Karic, T.; Røe, C.; Nordenmark, T.H.; Becker, F.; Sorteberg, W.; Sorteberg, A. Effect of early mobilization and rehabilitation on complications in aneurysmal subarachnoid hemorrhage. J. Neurosurg. 2017, 126, 518–526. [Google Scholar] [CrossRef] [Green Version]
- Hughe, J.D.; Bond, K.M.; Mekary, R.A.; Dewan, M.C.; Rattani, A.; Baticulon, R.; Kato, Y.; Azevedo-Filho, H.; Morcos, J.J.; Park, K.B. Estimating the Global Incidence of Aneurysmal Subarachnoid Hemorrhage: A Systematic Review for Central Nervous System Vascular Lesions and Meta-Analysis of Ruptured Aneurysms. World Neurosurg. 2018, 115, 430–447. [Google Scholar] [CrossRef]
- Johnston, S.C.; Selvin, S.; Gress, D.R. The burden, trends, and demographics of mortality from subarachnoid hemorrhage. Neurology 1998, 50, 1413–1418. [Google Scholar] [CrossRef]
- Ingall, T.; Asplund, K.; Mahonen, M.; Bonita, R. A multinational comparison of subarachnoid hemorrhage epidemiology in the WHO MONICA stroke study. Stroke 2000, 31, 1054–1061. [Google Scholar] [CrossRef]
- Dhandapani, S.; Singh, A.; Singla, N.; Praneeth, K.; Aggarwal, A.; Sodhi, H.B.; Pal, S.S.; Goudihalli, S.; Salunke, P.; Mohindra, S.; et al. Has outcome of subarachnoid hemorrhage changed with improvements in neurosurgical services? Stroke 2018, 49, 2890–2895. [Google Scholar] [CrossRef] [PubMed]
- Osman, N.; Ramesh, N. Perimesencephalic non-aneurysmal subarachnoid haemorrhage. BMJ Case Rep. 2018, 2018, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koffijberg, H.; Buskens, E.; Granath, F.; Adami, J.; Ekbom, A.; Rinkel, G.J.; Blomqvist, P. Subarachnoid haemorrhage in Sweden 1987–2002: Regional incidence and case fatality rates. J. Neurol. Neurosurg. Psychiatry 2008, 79, 294–299. [Google Scholar] [CrossRef] [PubMed]
- van Munster, C.E.; von und zu Fraunberg, M.; Rinkel, G.J.; Rinne, J.; Koivisto, T.; Ronkainen, A. Differences in aneurysm and patient characteristics between cohorts of Finnish and Dutch patients with subarachnoid hemorrhage: Time trends between 1986 and 2005. Stroke 2008, 39, 3166–3171. [Google Scholar] [CrossRef]
- Wáng, Y.X.; He, J.; Zhang, L.; Li, Y.; Zhao, L.; Liu, H.; Yang, L.; Zeng, X.J.; Yang, J.; Peng, G.M.; et al. A higher aneurysmal subarachnoid hemorrhage incidence in women prior to menopause: A retrospective analysis of 4,895 cases from eight hospitals in China. Quant. Imaging Med. Surg. 2016, 6, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Donkor, E.S. Stroke in the century: A snapshot of the burden, epidemiology, and quality of life. Stroke Res. Treat. 2018, 2018, 3238165. [Google Scholar] [PubMed] [Green Version]
- de Falco, F.A. Sentinel headache. Neurol. Sci. 2004, 25, S215–S217. [Google Scholar] [CrossRef]
- Aboulhosn, R.; Raju, B.; Jumah, F.; Majmundar, N.; Prenner, J.; Matin, T.; Roychowdhury, S.; Singla, A.; Khandelwal, P.; Nanda, A.; et al. Terson’s syndrome, the current concepts and management strategies: A review of literature. Clin. Neurol. Neurosurg. 2021, 210, 107008. [Google Scholar] [CrossRef]
- Salem, R.; Vallée, F.; Dépret, F.; Callebert, J.; Saint Maurice, J.P.; Marty, P.; Matéo, J.; Madadaki, C.; Houdart, E.; Bresson, D.; et al. Subarachnoid hemorrhage induces an early and reversible cardiac injury associated with catecholamine release: One-week follow-up study. Crit. Care 2014, 18, 558. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.Y.; Shin, J.I. Syndrome of inappropriate antidiuretic hormone secretion and cerebral/renal salt wasting syndrome: Similarities and differences. Front. Pediatr. 2015, 2, 146. [Google Scholar] [CrossRef] [Green Version]
- Fontanarosa, P.B. Recognition of subarachnoid hemorrhage. Ann. Emerg. Med. 1989, 18, 1199–1205. [Google Scholar] [CrossRef]
- Hunt, W.E.; Hess, R.M. Surgical risk as related to time of intervention in the repair of intracranial aneurysms. J. Neurosurg. 1968, 28, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.; Danesh, J.; Kuruvilla, D. Cerebral Venous Thrombosis Headache. Curr. Pain Headache Rep. 2019, 23, 47. [Google Scholar] [CrossRef] [PubMed]
- Aksamit, A.J., Jr.; Berkowitz, A.L. Meningitis. Contin. MinneapMinn 2021, 27, 836–854. [Google Scholar] [CrossRef] [PubMed]
- Sacco, S.; Russo, A.; Geppetti, P.; Grazzi, L.; Negro, A.; Tassorelli, C.; Tedeschi, G.; Martelletti, P. What is changing in chronic migraine treatment? An algorithm for onabotulinumtoxin A treatment by the Italian chronic migraine group. Expert Rev. Neurother. 2020, 20, 1275–1286. [Google Scholar] [CrossRef]
- Mazur, R.; Świerkocka-Miastkowska, M. Udar mózgu—Pierwsze objawy. Chor. Serca I Naczyń 2005, 2, 84–87. [Google Scholar]
- Tasiou, A.; Brotis, A.G.; Tzerefos, C.; Lambrianou, X.; Fountas, K.N. Methodological assessment of guidelines for the diagnosis and management of cerebral vasospasm using the AGREE-II tool. Neurosurg. Focus 2022, 52, E11. [Google Scholar] [CrossRef]
- Colmer, H.G.; Powell, E.K.; Kreitzer, N. Acute Stroke: From Prehospital Care to Inhospital Management. J. Emerg. Med. Serv. May 2018. Available online: https://www.jems.com/articles/print/volume-43/issue-5/features/acute-stroke-from-rehospital-care-to-in-hospital-management.html (accessed on 20 May 2019).
- Woodcock, R., Jr.; Short, J.; Do, H.M.; Jensen, M.E.; Kallmes, D.F. Imaging of Acute Subarachnoid Hemorrhage with a Fluid -Attenuated Inversion Recovery Sequence in an Animal Model: Comparison with Non–Contrast-Enhanced CT. AJNR Am. J. Neuroradiol. 2001, 22, 1698–1703. [Google Scholar]
- Diringer, M.N.; Zazulia, A.R. Aneurysmal Subarachnoid Hemorrhage: Strategies for preventing vasospam in the intensive care unit. Semin. Respir. Crit. Care Med. 2017, 38, 760–767. [Google Scholar]
- Jagt, M. Fluid management of neurological patient: A concise review. Crit. Care 2016, 20, 126. [Google Scholar] [CrossRef] [Green Version]
- Dash, H. Recent advances in the management of postoperative intracerebral vasospasm. J. Neuroanaesth. Critl. Care 2017, 4, S56–S59. [Google Scholar]
- Chen, S.; Luo, J.; Reis, C.; Manaenko, A.; Zhang, J. Hydrocephalus after Subarachnoid Hemorrhage: Pathophysiology, Diagnosis, and Treatment. BioMed Res. Int. 2017, 2017, 8584753. [Google Scholar] [CrossRef] [Green Version]
- Ramos, M.B.; Teixeira, M.J.; Figueiredo, E.F. Seizures and Epilepsy following Subarachnoid Hemorrhage: A Review on Incidence, Risk Factors, Outcome and Treatment. Arq. Bras. Neurocir. 2018, 37, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.L.; Soo, K.M.; Chen, C.W.; Lin, Y.K.; Lin, T.Y.; Kuo, L.C.; Lee, W.C.; Huang, S.L. Incidence, national trend, and outcome of nontraumatic subarachnoid haemorrhage in Taiwan: Initial lower mortality, poor long-term outcome. BioMed Res. Int. 2014, 2014, 274572. [Google Scholar] [CrossRef] [PubMed]
- Korja, M.; Silventoinen, K.; McCarron, P.; Zdravkovic, S.; Skytthe, A.; Haapanen, A.; de Faire, U.; Pedersen, N.L.; Christensen, K.; Koskenvuo, M.; et al. Genetic epidemiology of spontaneous subarachnoid hemorrhage: Nordic twin study. Stroke 2010, 41, 2458–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skodvin, T.; Evju, Ø.; Sorteberg, A.; Isaksen, J.G. Prerupture intracranial aneurysm morphology in predicting risk of rupture: A matched case-control study. Neurosurgery 2019, 84, 132–140. [Google Scholar] [CrossRef]
- Rivero-Arias, O.; Gray, A.; Wolstenholme, J. Burden of disease and costs of aneurysmal subarachnoid haemorrhage (aSAH) in the United Kingdom. Cost Eff. Resour. Alloc. 2010, 27, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, Y.; Kong, F.; Han, R.; Song, W.; Chen, D.; Bu, L.; Wang, S.; Yue, J.; Ma, L. Identification of a six-gene prognostic signature for oral squamous cell carcinoma. J. Cell Physiol. 2020, 235, 3056–3068. [Google Scholar] [CrossRef]
- Wang, Q.; Luo, Q.; Yang, Z.; Zhao, Y.H.; Li, J.; Wang, J.; Piao, J.; Chen, X. Weighted gene co-expression network analysis identified six hub genes associated with rupture of intracranial aneurysms. PLoS ONE 2020, 15, e0229308. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and functions of long non-coding RNAs at multiple regulatory levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Yi, J.; Zhou, T.; Gong, X.; Jiang, H.; Yao, X. Toward understanding non-coding RNA roles in intracranial aneurysms and subarachnoid hemorrhage. Transl. Neurosci. 2017, 8, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans. Plant. Cell 1990, 2, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, N.; Macino, G. Quelling: Transient inactivation of gene expression in Neurospora crassa by transformation with homologous sequences. Mol. Microbiol. 1992, 6, 3343–3353. [Google Scholar] [CrossRef]
- Pal-Bhadra, M.; Bhadra, U.; Birchler, J.A. Cosuppression in Drosophila: Gene silencing of Alcohol dehydrogenase by white-Adh transgenes is Polycomb dependent. Cell 1997, 90, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell 2000, 101, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Ambros, V. The evolution of our thinking about microRNAs. Nat. Med. 2008, 14, 1036–1040. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. A matter of life or death. World J. Biol. Chem. 2010, 1, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isik, M.; Korswagen, H.C.; Berezikov, E. Expression patterns of intronic microRNAs in Caenorhabditis elegans. Silence 2010, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shomron, N.; Levy, C. MicroRNA-biogenesis and pre-mRNA splicing crosstalk. J. Biomed. Biotechnol. 2009, 2009, 594678. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 20, 4051–4060. [Google Scholar] [CrossRef]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA polimerase III transcribes human microRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, D.L.; Perron, M.P.; Gobeli, L.A.; Plante, P.; Provost, P. MicroRNA in gene regulation: When the smallest governs it all. J. Biomed. Biotechnol. 2006, 2006, 69616. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, Y.; Yeom, K.H.; Nam, J.W.; Heo, I.; Rhee, J.K.; Sohn, S.Y.; Cho, Y.; Zhang, B.T.; Kim, V.N. Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Cullen, B.R. Structural requirements for pre-microRNA binding and nuclear export by Exportin 5. Nucl. Acids Res. 2004, 32, 4776–4785. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.S.; Hutvagner, G.; Du, T.; Xu, Z.; Aronin, N.; Zamore, P.D. Asymmetry in the assembly of the RNAi enzyme complex. Cell 2003, 115, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Liu, N.; Lai, E.C. Distinct mechanism for microRNA stranded selection by Drosophila Argonautes. Mol. Cell 2009, 36, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelfattah, A.M.; Park, C.; Choi, M.Y. Update on non-canonical microRNAs. Biomol. Concepts 2014, 5, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Hagen, J.W.; Duan, H.; Tyler, D.M.; Lai, E.C. The mintron pathway generates microRNA-class regulatory RNAs in Drosophila. Cell 2007, 130, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass Drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J.S. Small RNAs derived form snoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Ender, C.; Krek, A.; Fridlander, M.R.; Beitzinger, M.; Weinmann, L.; Chen, W.; Pfeffer, S.; Rajewsky, N.; Meister, G. A human snoRNA with microRNA-like functions. Mol. Cell 2008, 32, 519–528. [Google Scholar] [CrossRef]
- Babiarz, J.E.; Ruby, J.G.; Wang, Y.; Bartel, D.P.; Blelloch, R. Mouse ES cells express endogenous shRNAs, siRNAs, and other Microprocessor-independent, Dicer-dependent small RNAs. Genome Dev. 2008, 22, 2773–2785. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [Green Version]
- Cheluofi, S.; Dos Santos, C.O.; Chong, M.M.; Hannon, G.J. A Dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuentes, D.; Xue, H.; Taylor, D.W.; Patnode, H.; Mishima, Y.; Cheloufi, S.; Ma, E.; Mane, S.; Hannon, G.J.; Lawson, N.D.; et al. A novel miRNA procesing pathway independent of Dicer requires Argonaute2 catalytic activity. Science 2010, 328, 1694–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Integrating microRNA annotation and deep-sequencing data. Nucl. Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [Green Version]
- Bogerd, H.P.; Karnowski, H.W.; Cai, X.; Shin, J.; Pohlers, M.; Cullen, B.R. A mammalian herpesvirus uses noncanonical expression and processing mechanisms to generate viral microRNAs. Mol. Cell 2010, 37, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA virus microRNA that mimics a B-cell oncomir. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef] [Green Version]
- Kincaid, R.P.; Sullivan, C.S. Virus-encoded microRNAa: An overview and a look to the future. PLoS Pathog. 2012, 8, e1003018. [Google Scholar] [CrossRef] [Green Version]
- Kincaid, R.P.; Chen, Y.; Cox, J.E.; Rethwilm, A.; Sullivan, C.S. Noncanonical microRNA (miRNA) biogenesis gives rise to retroviral mimics of lymphoproliferative and immunosuppressive host miRNAs. MBio 2014, 5, e00074. [Google Scholar] [CrossRef] [Green Version]
- Lagos, D.; Pollara, G.; Henderson, S.; Gratrix, F.; Fabani, M.; Milne, R.S.; Gotch, F.; Boshoff, C. miR-132 regulates antiviral innate immunity through suppression of the p300 transcriptional co-activator. Nat. Cell Biol. 2010, 12, 513–519. [Google Scholar] [CrossRef]
- Braun, J.E.; Huntzinger, E.; Fauser, M.; Izaurralde, E. GW182 proteins directly recruit cytoplasmic deadenylase complexes to miRNA targets. Mol. Cell 2011, 44, 120–133. [Google Scholar] [CrossRef]
- Wienholds, E.; Plasterk, R.H.A. MicroRNA function in animal development. FEBS Lett. 2005, 579, 5911–5922. [Google Scholar] [CrossRef] [Green Version]
- Helak, A.; Kudla, G.; Dudnakova, T.; Tollervey, D. Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding. Cell 2013, 153, 654–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory function. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orom, U.A.; Nielson, F.C.; Lund, A.H. MicroRNA-10a binds the 5’UTR of ribosomal protein mRNAs and enhances their translation. Mol. Cell 2008, 30, 460–471. [Google Scholar] [CrossRef]
- Duursma, A.M.; Kedde, M.; Schrier, M.; le Sage, C.; Agami, R. miR-148 targets human DNMT3b protein coding region. RNA 2008, 14, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Fan, J.; Belasco, J.G. Micro RNAs direct rapid deadenylation of mRNA. Proc. Natl. Acad. Sci. USA 2006, 103, 4034–4039. [Google Scholar] [CrossRef] [Green Version]
- Nishihara, T.; Zekri, L.; Braun, J.E.; Izaurralde, E. miRISC recruits decapping factors to miRNA targets to enhance their degradation. Nuclic Acids Res. 2013, 41, 8692–8705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, D.T.; Westman, B.J.; Martin, D.I.K.; Preiss, T. MicroRNA control translation initiation by inhibiting eukaryotic initiation factor 4E/cap and poly(A) tail function. Proc. Natl. Acad. Sci. USA 2005, 102, 16961–16966. [Google Scholar] [CrossRef] [Green Version]
- Zdanowicz, A.; Thermann, R.; Kowalska, J.; Jemielity, J.; Duncan, K.; Preiss, T.; Darzynkiewicz, E.; Hentze, M.W. Drosophila miR2 primarily targets the m7G pppN cap structure for translational repression. Mol. Cell 2009, 35, 881–888. [Google Scholar] [CrossRef]
- Wang, B.; Yanez, A.; Novina, C.D. MicroRNA-repressed mRNAs contain 40S but not 60S components. Proc. Natl. Acad. Sci. USA 2008, 105, 5343–5348. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.P.; Bordeleau, M.E.; Pelletier, J.; Sharp, P. Short RNAs repress translation after initiation in mammalian cells. Mol. Cell 2006, 21, 533–542. [Google Scholar] [CrossRef]
- Politz, J.C.R.; Zhang, F.; Pederson, T. MicroRNA-206 colocalizes with ribosome-rich regions in both the nucleus and cytoplasm of rat myogenic cells. Proc. Natl. Acad. Sci. USA 2006, 103, 18957–18962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, J.I.; Goergen, D.; Zheng, J.; Song, Y.; Schuttler, C.G.; Fehr, C.; Junermann, C.; Niepmann, M. MicroRNA-122 stimulates translation of hepatitis C virus RNA. EMBO J. 2008, 27, 3300–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybycien-Szymanska, M.M.; Ashley, W.W., Jr. Biomarker discovery in cerebral vasospasm after aneurysmal subarachnoid hemorrhage. J. Stroke Cerebrovasc. Diseases Off. J. Natl. Stroke Assoc. 2015, 24, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- van’t Hof, F.N.; Ruigrok, Y.M.; Medic, J.; Sanjabi, B.; van der Vlies, P.; Rinkel, G.J.; Veldink, J.H. Whole blood gene expression profiles of patients with a past aneurysmal subarachnoid hemorrhage. PLoS ONE 2015, 10, e0139352. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Q.; Wu, X.; Yang, X.; Zhang, Y.; Li, Y.; Jiang, F. Circulating microRNAs serve as novel biological markers for intracranial aneurysms. J. Am. Heart Assoc. 2014, 3, e000972. [Google Scholar] [CrossRef] [Green Version]
- Gareev, I.; Beylerli, O.; Yang, G.; Izmailov, A.; Shi, H.; Sun, J.; Zhao, B.; Liu, B.; Zhao, S. Diagnostic and prognostic potential of circulating miRNAs for intracranial aneurysms. Neurosurg. Rev. 2021, 44, 2025–2039. [Google Scholar] [CrossRef]
- Wang, C.; Ji, B.; Cheng, B.; Chen, J.; Bai, B. Neuroprotection of microRNA in neurological disorders (Review). Biomed. Rep. 2014, 2, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, E.R.; Okun, E.; Mattson, M.P. The therapeutic potential of microRNAs in nervous system damage, degeneration, and repair. Neuromol. Med. 2009, 11, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Backes, C.; Meese, E.; Keller, A. Specific miRNA disease biomarkers in blood, serum and plasma: Challenges and prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef]
- Feng, X.; Peng, F.; Zhang, B.; Wang, L.; Guo, E.; Li, Y.; Jiang, C.; Wu, Z.; Liu, A. Lower miR-143/145 and higher matrix metalloproteinase-9 levels in circulation may be associated with intracranial aneurysm formation and rupture: A pilot study. Clin. Neurol. Neurosurg. 2018, 173, 124–129. [Google Scholar] [CrossRef]
- Yang, X.; Peng, J.; Pang, J.; Wan, W.; Chen, L. A functional polymorphism in the promoter region of miR-155 predicts the risk of intracranial hemorrhage caused by rupture intracranial aneurysm. J. Cell Biochem. 2019, 120, 18618–18628. [Google Scholar] [CrossRef] [PubMed]
- Welten, S.M.; Goossens, E.A.; Quax, P.H.; Nossent, A.Y. The multifactorial nature of microRNAs in vascular remodelling. Cardiovasc. Res. 2016, 110, 6–22. [Google Scholar] [CrossRef]
- Supriya, M.; Christopher, R.; Indira Devi, B.; Bhat, D.I.; Shukla, D. Circulating MicroRNAs as potential molecular biomarkers for intracranial. Mol. Diagn. Ther. 2020, 24, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Nozaki, K. Preemptive medicine for cerebral aneurysms. Neurol. Med. Chir. 2016, 56, 552–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, B.; Lai, N.S.; Yao, Y.; Dong, J.; Li, Z.B.; Zhao, X.T.; Liu, J.Q.; Li, X.Q.; Fang, X.G. Early serummiR-1297 is an indicator of poor neurological outcome in patients with aSAH. Biosci. Rep. 2018, 38, BSR20180646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, B.; Fang, X.; Liu, C.; Wu, D.; Xia, D.; Xu, S.; Lai, N. Persistent high levels of miR-502-5p are associated with poor neurologicoutcome in patients with aneurysmal subarachnoid hemorrhage. World Neurosurg. 2018, 116, e92–e99. [Google Scholar] [CrossRef]
- Lai, N.S.; Zhang, J.Q.; Qin, F.Y.; Sheng, B.; Fang, X.G.; Li, Z.B. Serum microRNAs are non-invasive biomarkers for the presenceand progression of subarachnoid haemorrhage. Biosci. Rep. 2017, 37, BSR20160480. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.H.; Wang, Y.H.; Zheng, L.L.; Li, X.W.; Hao, F.; Guo, D. MicroRNA-29a: A potential biomarker in the development of intracranial aneurysm. J. Neurol. Sci. 2016, 364, 84–89. [Google Scholar] [CrossRef]
- Meeuwsen, J.A.L.; van Hof, F.N.G.T.; van Rheenen, W.; Rinkel, G.J.E.; Veldink, J.H.; Ruigrok, Y.M. Circulating microRNAs in patients with intracranial aneurysms. PLoS ONE 2017, 12, e0176558. [Google Scholar]
- Yang, F.; Xing, W.W.; Shen, D.W.; Tong, M.F.; Xie, F.M. Effect of miR-126 on intracranial aneurysms and its predictive value for rupture of aneurysms. Eur Rev. Med. Pharmacol. Sci. 2020, 24, 3245–3253. [Google Scholar]
- Su, X.W.; Chan, A.H.; Lu, G.; Lin, M.; Sze, J.; Zhou, J.Y.; Poon, W.S.; Liu, Q.; Zheng, V.Z.; Wong, G.K. Circulating microRNA 132-3p and 324-3p profiles in patients after acute aneurysmal subarachnoid hemorrhage. PLoS ONE 2015, 10, e0144724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, C.J.; Dickerson, R.; Zhang, S.W.; Rink, C.; Roy, S.; Sen, C.K. Human cerebrospinal fluid microRNA: Temporal changes following subarachnoid hemorrhage. Physiol. Genom. 2016, 48, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, A.V.; Palade, C.; Voinescu, D.; Nica, D.A. Subarachnoid hemorrhage and cerebral vasospasm—Literature review. J. Med. Life 2013, 6, 120–125. [Google Scholar] [PubMed]
- Kikkawa, Y.; Ogura, T.; Nakajima, H.; Ikeda, T.; Takeda, R.; Neki, H.; Kohyama, S.; Yamane, F.; Kurogi, R.; Amano, T.; et al. Altered expression of MicroRNA-15a and Kruppel-like factor 4 in cerebrospinal fluid and plasma after aneurysmal subarachnoid hemorrhage. World Neurosurg. 2017, 108, 909–916. [Google Scholar] [CrossRef]
- Kim, S.H.; Weiß, C.; Hoffmann, U.; Borggrefe, M.; Akin, I.; Behnes, M. Advantages and limitations of current biomarker research: From experimental research to clinical application. Curr. Pharm. Biotechnol. 2017, 18, 445–455. [Google Scholar] [CrossRef]
- Zaporozhchenko, I.A.; Ponomaryova, A.A.; Rykova, E.Y.; Laktionov, P.P. The potential of circulating cell-free RNA as a cancer bio-marker: Challenges and opportunities. Expert Rev. Mol. Diagn. 2018, 18, 133. [Google Scholar] [CrossRef]
- Li, M.; Zhang, J. Circulating MicroRNAs: Potential andemerging biomarkers for diagnosis of cardiovascular and cerebrovascular diseases. BioMed Res. Int. 2015, 2015, 730535–730539. [Google Scholar] [CrossRef]
- Korostynski, M.; Morga, R.; Piechota, M.; Hoinkis, D.; Golda, S.; Dziedzic, T.; Slowik, A.; Moskala, M.; Pera, J. Inflammatory responses induced by the rupture of intracranial aneurysms are modulated by miRNAs. Mol. Neurobiol. 2020, 57, 988–996. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, S. Aneurysmal subarachnoid hemorrhage. J. Neurosurg. Anesthesiol. 2015, 27, 222–240. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, R.L.; Schweizer, T.A. Spontaneous subarachnoid haemorrhage. Lancet 2017, 389, 655–666. [Google Scholar] [CrossRef]
- Crobeddu, E.; Pilloni, G.; Tardivo, V.; Fontanella, M.M.; Panciani, P.P.; Spena, G.; Fornaro, R.; Altieri, R.; Agnoletti, A.; Ajello, M.; et al. Role of nitric oxide and mechanisms involved in cerebral injury after subarachnoid hemorrhage: Is nitric oxide a possible answer to cerebral vasospasm? J. Neurosurg. Sci. 2016, 60, 385–391. [Google Scholar] [PubMed]
- Li, H.T.; Wang, J.; Li, S.F.; Cheng, L.; Tang, W.Z.; Feng, Y.G. Upregulation of microRNA-24 causes vasospasm following subarachnoid hemorrhage by suppressing the expression of endothelial nitric oxide synthase. Mol. Med. Rep. 2018, 18, 1181–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulcrano-Nicolas, A.S.; Proust, C.; Clarençon, F.; Jacquens, A.; Perret, C.; Roux, M.; Shotar, E.; Thibord, F.; Puybasset, L.; Garnier, S.; et al. Whole-blood miRNA sequencing profiling for vasospasm in patients with aneurysmal subarachnoid hemorrhage. Stroke 2018, 49, 2220–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.H.; Zhou, X.M.; Zhang, J.X.; Gao, H.; Jiang, J.F.; Liu, N. Circulating microRNAs act as fingerprints in patients after acuteaneurysmal subarachnoid hemorrhage. Int. J. Clin. Exp. Pathol. 2017, 10, 7154–7160. [Google Scholar]
- Lopes, K.P.; Vinasco-Sandoval, T.; Vialle, R.A.; Paschoal, F.M., Jr.; Bastos, V.A.P.A.; Bor-Seng-Shu, E.; Teixeira, M.J.; Yamada, E.S.; Pinto, P.; Vidal, A.F.; et al. Global miRNA expression profile reveals novel molecular players in aneurysmal subarachnoid haemorrhage. Sci. Rep. 2018, 8, 8786. [Google Scholar] [CrossRef]
- Bache, S.; Rasmussen, R.; Rossing, M.; Laigaard, F.P.; Nielsen, F.C.; Møller, K. MicroRNA changes in cerebrospinal fluid after subarachnoid. hemorrhage. Stroke 2017, 48, 2391–2398. [Google Scholar] [CrossRef]
- Stylli, S.S.; Adamides, A.A.; Koldej, R.M.; Luwor, R.B.; Ritchie, D.S.; Ziogas, J.; Kaye, A.H. miRNA expression profiling of cerebrospinal fluid in patients with aneurysmal subarachnoid hemorrhage. J. Neurosurg. 2017, 126, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- An, J.Y.; Zhou, L.L.; Sun, P.; Pang, H.G.; Li, D.D.; Li, Y.; Zhang, M.; Song, J.N. Role of the AMPK signaling pathway in early brain injury after subarachnoid hemorrhage in rats. Acta Neurochir. 2015, 157, 781–792. [Google Scholar] [CrossRef]
- Weng, Y. Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage. Open Life Sci. 2021, 16, 1377–1392. [Google Scholar] [CrossRef]
- Kato, H.; Kawaguchi, M.; Inoue, S.; Hirai, K.; Furuya, H. The effects of beta-adrenoceptor antagonists on proinflammatory cytokine concentrations after subarachnoid hemorrhage in rats. Anesth. Analg. 2009, 108, 288–295. [Google Scholar] [CrossRef]
- Yan, H.; Chen, Y.; Li, L.; Jiang, J.; Wu, G.; Zuo, Y.; Zhang, J.H.; Feng, H.; Yan, X.; Liu, F. Decorin alleviated chronic hydrocephalus via inhibiting TGF-β1/Smad/CTGF pathway after subarachnoid hemorrhage in rats. Brain Res. 2016, 1630, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, L.; Xie, D.; Niu, J. Protective effects of transforming growth factor-β1 knockdown in human umbilical cord mesenchymal stem cells against subarachnoid hemorrhage in a rat model. Cerebrovasc. Dis. 2020, 49, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Niu, T.; Li, X. MicroRNA-126-3p Attenuates Intracerebral Hemorrhage-Induced Blood-Brain Barrier Disruption by Regulating VCAM-1 Expression. Front. Neurosci. 2019, 13, 866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, T.; Jin, F.; Zhu, Y.; Wang, J.; Tang, L.; Wang, Y.; Liebeskind, D.S.; Scalzo, F.; He, Z. miR-27a-3p protects against blood-brain barrier disruption and brain injury after intracerebral hemorrhage by targeting endothelial aquaporin-11. J. Biol. Chem. 2018, 293, 20041–20050. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Li, L.; Hu, D.; Zhang, X. LINC00612/miR-31-5p/Notch1 Axis Regulates Apoptosis, Inflammation, and Oxidative Stress in Human Pulmonary Microvascular Endothelial Cells Induced by Cigarette Smoke Extract. Int. J. Chronic Obstr. Pulm. Dis. 2020, 15, 2049–2060. [Google Scholar] [CrossRef]
- Cai, L.; Ge, B.; Xu, S.; Chen, X.; Yang, H. Up-regulation of circARF3 reduces blood-brain barrier damage in rat subarachnoid hemorrhage model via miR-31-5p/MyD88/NF-κB axis. Aging 2021, 13, 21345–21363. [Google Scholar] [CrossRef]
- Rahmati, S.; Shojaei, F.; Shojaeian, A.; Rezakhani, L.; Dehkordi, M.B. An overview of current knowledge in biological functionsand potential theragnostic applications of exosomes. Chem. Phys. Lipids 2020, 226, 104836. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhang, X.; Li, L.; Wang, C.; Feng, M.; Zhao, K.; Zhao, R.; Liu, J.; Fang, Y. Tumor-associated macrophage-derived exosomal microRNA-155-5p stimulates intracranial aneurysm formation and macrophage infiltration. Clin. Sci. 2019, 133, 2265–2282. [Google Scholar] [CrossRef]
- Falcone, G.; Felsani, A.; D’Agnano, I. Signaling by exosomal microRNAs in cancer. J. Exp. Clin. Cancer Res. 2015, 34, 32. [Google Scholar] [CrossRef] [Green Version]
- Liao, B.; Zhou, M.X.; Zhou, F.K.; Luo, X.M.; Zhong, S.X.; Zhou, Y.F.; Qin, Y.S.; Li, P.P.; Qin, C. Exosome-derivedMiRNAs as biomarkersof the development and progression of intracranial aneurysms. J. Atheroscler. Thromb. 2019, 27, 545–610. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zhang, W.; Li, Z.; Li, M.; Guo, J.; Wang, H.; Wang, X. The expression of cerebrospinal fluid exosomal miR-630 plays an import ant role in the dysfunction of endothelial cells after subarachnoid hemorrhage. Sci. Rep. 2019, 9, 11510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Yin, W.N.; Suzuki, T.; Zhang, X.H.; Zhang, Y.; Song, L.L.; Jin, L.S.; Zhan, H.; Zhang, H.; Li, J.S.; et al. Exosome-mediatedmiR-155 transfer from smooth muscle cells to endothelial cells induces endothelial injury and promotes atherosclerosis. Mol. Ther. 2017, 25, 1279–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalani, M.Y.S.; Alsop, E.; Meechoovet, B.; Beecroft, T.; Agrawal, K.; Whitsett, T.G.; Huentelman, M.J.; Spetzler, R.F.; Nakaji, P.; Kim, S.; et al. Extracellular microRNAs in blood differentia te between ischaemic and haemorrhagic stroke subtypes. J. Extracell. Vesicles 2020, 9, 1713540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, N.; Wu, D.; Liang, T.; Pan, P.; Yuan, G.; Li, X.; Li, H.; Shen, H.; Wang, Z.; Chen, G. Systemic exosomal miR-193b-3p delivery attenuates neuroinflammation in early brain injury after subarachnoid hemorrhage in mice. J. Neuroinflamm. 2020, 17, 74. [Google Scholar] [CrossRef] [PubMed]
- Strimbu, K.; Tavel, J.A. What are biomarkers? Curr. Opin. HIV AIDS 2010, 5, 463–466. [Google Scholar] [CrossRef]
- Willeit, P.; Zampetaki, A.; Dudek, K.; Kaudewitz, D.; King, A.; Kirkby, N.S.; Crosby-Nwaobi, R.; Prokopi, M.; Drozdov, I.; Langley, S.R.; et al. Circulating microRNAs as novel biomarkers forplatelet activation. Circ. Res. 2013, 112, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Mattox, A.K.; Yan, H.; Bettegowda, C. The potential of cerebrospinal fluid-based liquid biopsy approaches in CNS tumors. Neuro Oncol. 2019, 21, 1509–1518. [Google Scholar] [CrossRef] [Green Version]
- Donati, S.; Ciuffi, S.; Brandi, M.L. Human circulating miRNAs real-time qRT-PCR-based analysis: An overview of endogenousreference genes used for data normalization. Int. J. Mol. Sci. 2019, 20, 4353. [Google Scholar] [CrossRef] [Green Version]
- Schlosser, K.; McIntyre, L.A.; White, R.J.; Stewart, D.J. Customized internal reference controls for improved assessmentof circulating MicroRNAs in disease. PLoS ONE 2015, 10, e0127443. [Google Scholar] [CrossRef]
- Xiang, M.; Zeng, Y.; Yang, R.; Xu, H.; Chen, Z.; Zhong, J.; Xie, H.; Xu, Y.; Zeng, X. U6 is not a suitable endogenous control for thequantification of circulating microRNAs. Biochem. Biophys. Res. Commun. 2014, 454, 210–214. [Google Scholar] [CrossRef]
- Bottani, M.; Banfi, G.; Lombardi, G. Circulating miRNAs as diagnostic and prognostic biomarkers in common solid tumors: Focus on lung, breast, prostate cancers, and osteosarcoma. J. Clin. Med. 2019, 8, 1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Grade | Symptoms |
|---|---|
| I | No symptoms, possible presence of neck stiffness |
| II | Severe headaches, neck stiffness, cranial nerve paralysis |
| III | Confusion, drowsiness, slight focal symptoms |
| IV | Stupor, hemiparesis, decerebrate rigidity, vegetative disorders |
| V | Deep coma, decerebrate rigidity, moribund |
| Grade | Glasgow Scale | Movement Disorders |
|---|---|---|
| I | 15 | Absent |
| II | 14–13 | Absent |
| III | 14–13 | Present |
| IV | 12–7 | Present or not |
| V | 6–3 | Present or not |
| Grade | Amount of Blood in Computed Tomography |
|---|---|
| I | Lack of blood in the computed tomography image |
| II | Leaked or thin layer of blood < 1 mm thick |
| III | Localized clot and/or blood layer > 1 mm thick |
| IV | Intraventricular and/or interstitial blood clot |
| miRNA | Regulation | |
|---|---|---|
| Diagnostic of IAs or aSAH | miR-1297 | Up [106] |
| miR-502-5p | Down [107] | |
| miR-4320 | Up [108] | |
| miR-143 | Down [101] | |
| miR-145 | Down [101] | |
| miR-155 | Up [102] | |
| miR-29a | Down [109] | |
| miR-200a-3p | Up [110] | |
| miR-let7-b | Down [110] | |
| miR-16 | Up [96] | |
| miR-25 | Up [96] | |
| miR-15a-5p | Up [104] | |
| miR-146-5p | Down [104] | |
| miR-126 | Up [111] | |
| miR-132-3p | Up [112] | |
| Prognostic of aSAH | miR-1297 | Up [106] |
| miR-502-5p | Down [107] | |
| miR-29a | Down [109] | |
| miR-200a-3p | Up [110] | |
| miR-146-5p | Down [104] | |
| miR-92a | Down [113] | |
| let-7b | Down [113] | |
| miR-3177-3p | Up [114] | |
| miR-132-3p | Up [112] | |
| miR-15a | Up [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makowska, M.; Smolarz, B.; Romanowicz, H. microRNAs in Subarachnoid Hemorrhage (Review of Literature). J. Clin. Med. 2022, 11, 4630. https://doi.org/10.3390/jcm11154630
Makowska M, Smolarz B, Romanowicz H. microRNAs in Subarachnoid Hemorrhage (Review of Literature). Journal of Clinical Medicine. 2022; 11(15):4630. https://doi.org/10.3390/jcm11154630
Chicago/Turabian StyleMakowska, Marianna, Beata Smolarz, and Hanna Romanowicz. 2022. "microRNAs in Subarachnoid Hemorrhage (Review of Literature)" Journal of Clinical Medicine 11, no. 15: 4630. https://doi.org/10.3390/jcm11154630