
Pathogenesis of Human Adenomyosis: Current Understanding and Its Association with Infertility


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidemiology
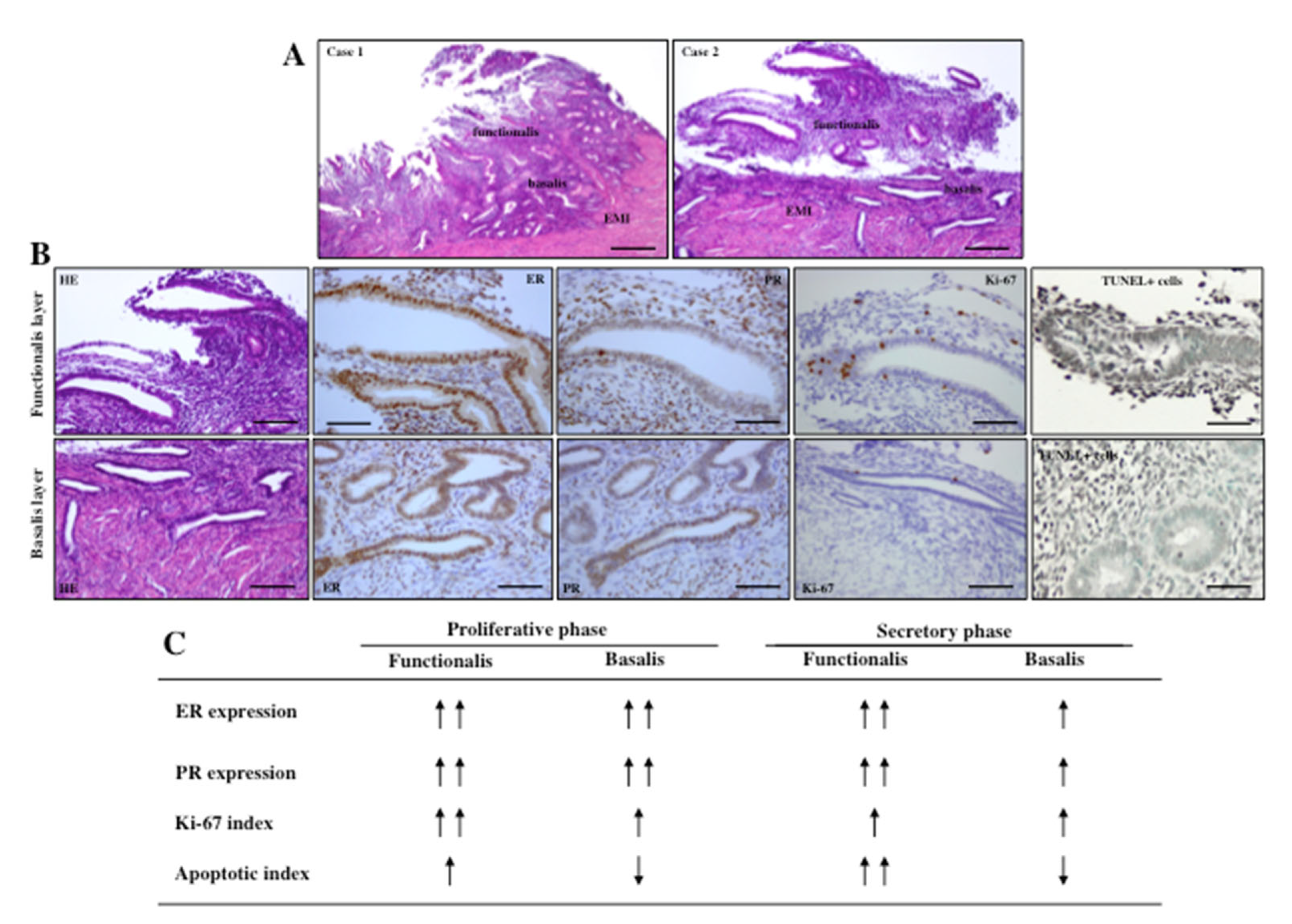
3. Biology of Endometrium and Myometrium
4. Etiology and Pathogenesis of Adenomyosis
4.1. Invagination: Role of Estrogen, TIAR and EMID
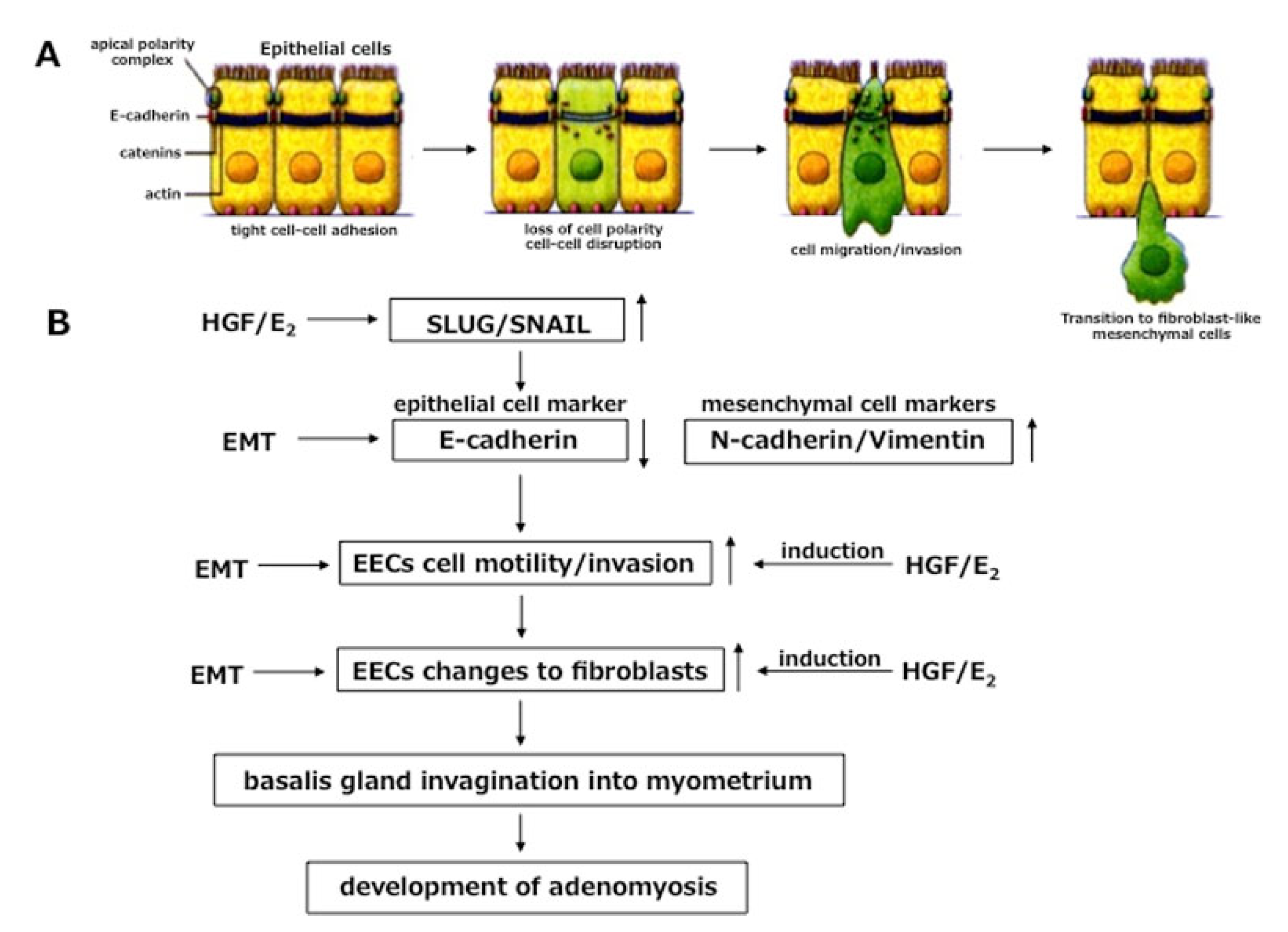
4.2. Role of EMT in Invagination
4.3. Metaplasia of Müllerian Remnants
4.4. Role of Endometrial Stem/Progenitor Cells
4.5. Genetic and Epigenetic Alteration in Adenomyosis
4.6. Somatic Mutation in Adenomyosis
4.7. Role of microRNAs in Adenomyosis
5. Clinical Classification of Adenomyosis
5.1. Biological Differences between Focal and Diffuse Adenomyosis
5.2. Biological Differences between Intrinsic and Extrinsic Adenomyosis
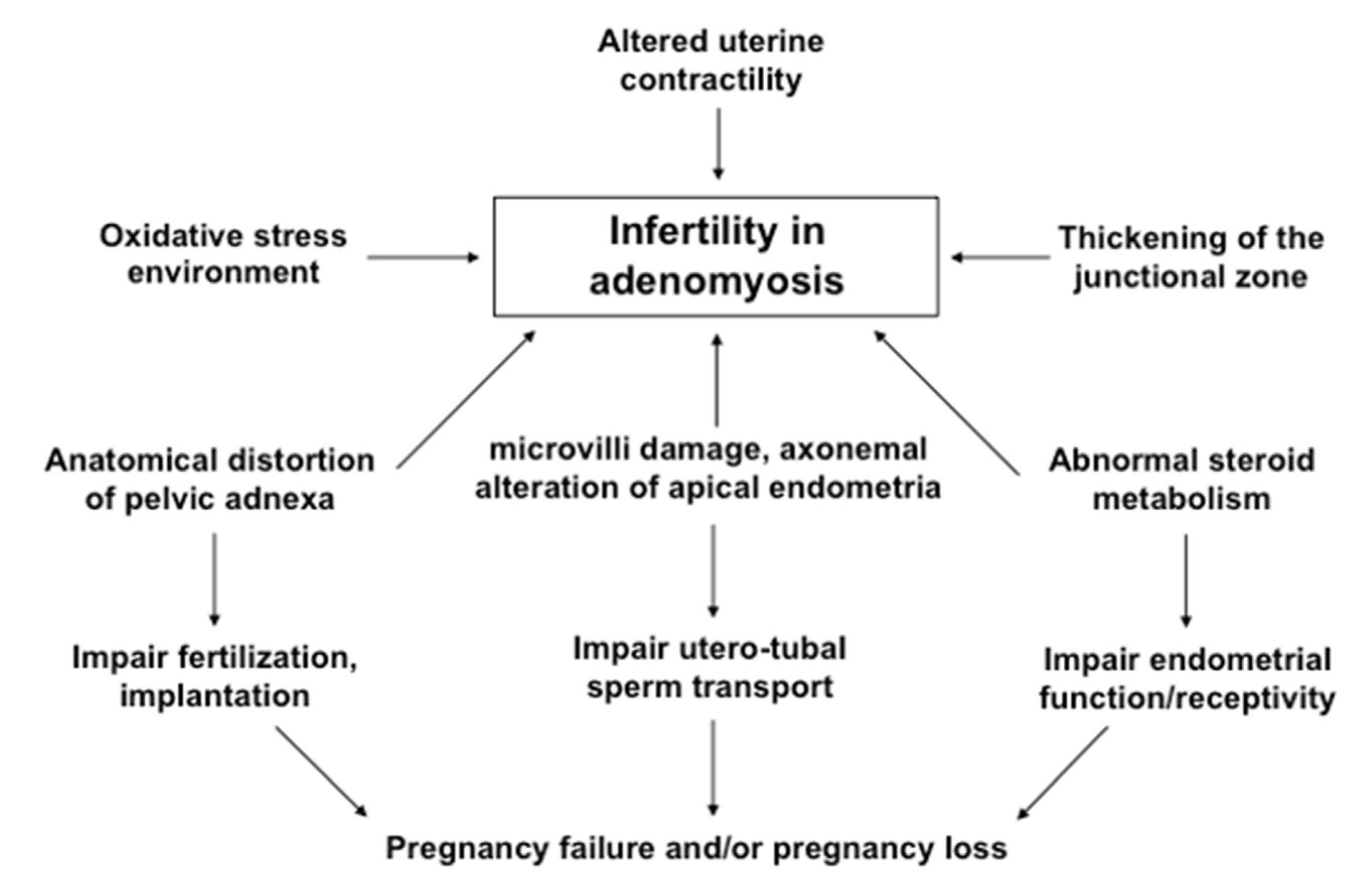
6. Association with Infertility
6.1. Proposed Mechanisms
6.2. Role of Microvilli and Axonemal Alteration
7. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergeron, C.; Amant, F.; Ferenczy, A. The pathology and physiopathology of adenomyosis. Best Pract. Res. Clin. Obstet. Gynecol. 2006, 20, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Ferenczy, A. Pathophysiology of adenomyosis. Hum. Reprod. Update 1998, 4, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, M.G. Classification and reporting systems for adenomyosis. J. Minim. Invasive Gynecol. 2020, 27, 296–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emage, L.A. The elusive adenomyosis of the uterus. Its historical past and its present state of recognition. Am. J. Obstet. Gynecol. 1962, 83, 1541–1563. [Google Scholar]
- Bulun, S.E.; Yildiz, S.; Adli, M.; Wei, J.-J. Adenomyosis pathogenesis: Insights from next generation sequencing. Hum. Reprod. Update 2021, 27, 1086–1097. [Google Scholar] [CrossRef]
- Chapron, C.; Tosti, C.; Marcellin, L.; Bourdon, M.; Lafay-Pillet, M.C.; Millischer, A.E.; Streuli, I.; Borghese, B.; Petraglia, F.; Santulli, P. Relationship between the magnetic resonance imaging appearance of adenomyosis and endometriosis phenotypes. Hum. Reprod. 2017, 32, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Leyendecker, G.; Bilgicyildirim, A.; Inacker, M.; Stalf, T.; Huppert, P.; Mall, G.; Bottcher, B.; Wildt, L. Adenomyosis and endometriosis. Re-visiting their association and further insights into the mechanisms of auto-traumatisation. An MRI study. Arch. Gynecol. Obstet. 2015, 291, 917–932. [Google Scholar] [CrossRef] [Green Version]
- Kunz, G.; Beil, D.; Huppert, P.; Noe, M.; Kissler, S.; Leyendecker, G. Adenomyosis in endometriosis-prevalence and impact on fertility. Evidence from magnetic resonance imaging. Hum. Reprod. 2005, 20, 2309–2316. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Chen, M.; Liu, X.; Guo, S.W. Constitutive and tumor necrosis factor-alpha induced activation of nuclear factor kappa-B in adenomyosis and its inhibition by andrographolide. Fertil. Steril. 2013, 100, 568–577. [Google Scholar] [CrossRef]
- Donnez, O.; Van Langendonckt, A.; Defrere, S.; Colette, S.; Van Kerk, O.; Dehoux, J.P.; Suifflet, J.; Donnez, J. Induction of endometriosis nodules in an experimental baboon model mimicking human deep nodular lesions. Fertil. Steril. 2013, 99, 783–789.e3. [Google Scholar] [CrossRef]
- Donnez, O.; Orellana, R.; Van Kerk, O.; Dehoux, J.P.; Donnez, J.; Dolmans, M.M. Invasion process of induced deep nodular endometriosis in an experimental baboon model: Similarities with collective cell migration? Fertil. Steril. 2015, 104, 491–497.e2. [Google Scholar] [CrossRef] [PubMed]
- Benagiano, G.; Brosens, I.; Habiba, M. Structural and molecular features of the endomyometrium in endometriosis and adenomyosis. Hum. Reprod. Update 2014, 20, 386–402. [Google Scholar] [CrossRef]
- Upson, K.; Missmer, S.A. Epidemiology of adenomyosis. Semin. Reprod. Med. 2020, 38, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W. The pathogenesis of adenomyosis vis-à-vis endometriosis. J. Clin. Med. 2020, 9, 845–863. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Solares, J.; Donnez, J.; Donnez, O.; Dolmans, M.M. Pathogenesis of uterine adenomyosis: Invagination or metaplasia? Fertil. Steril. 2018, 109, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Nakashima, M.; Masuzaki, H. Involvement of hepatocyte growth factor-induced epithelial-mesenchymal transition in human adenomyosis. Biol. Reprod. 2015, 92, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Fujishita, A.; Kitajima, M.; Masuzaki, H.; Nakashima, M.; Kitawaki, J. Biological differences between functionalis and basalis endometria in women with and without adenomyosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 203, 49–55. [Google Scholar] [CrossRef]
- Zhai, J.; Vannuccini, S.; Petralgia, F.; Giuduce, L.C. Adenomyosis: Mechanisms and pathogenesisi. Semin. Reprod. Med. 2020, 38, 129–143. [Google Scholar] [CrossRef]
- Exacoustos, C.; Manganaro, L.; Zupi, E. Imaging for the evaluation of endometriosis and adenomyosis. Best Pract. Res. Clin. Obstet. Gynecol. 2014, 28, 655–681. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.H.; Fuh, J.L.; Chao, H.T.; Liu, W.M.; Cheng, M.H.; Chao, K.C. Is the surgical approach beneficial to subfertile women with symptomatic extensive adenomyosis? J. Obstet. Gynecol. Res. 2009, 35, 495–502. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Guo, S.W. Clinical profiles of 710 pre-menopausal women with adenomyosis who underwent hysterectomy. J. Obstet. Gynecol. Res. 2014, 40, 485–494. [Google Scholar] [CrossRef]
- Azziz, R. Adenomyosis: Current perspectives. Obstet. Gynecol. Clin. N. Am. 1989, 16, 221–235. [Google Scholar] [CrossRef]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Daguati, R.; Abbiati, A.; Fedele, L. Ademonyosis: Epidemiological factors. Best Pract. Res. Clin. Obstet. Gynecol. 2006, 20, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Bergholt, T.; Eriksen, L.; Berendt, N.; Jacobsen, M.; Hertz, J.B. Prevalence and risk factors of adenomyosis at hysterectomy. Hum. Reprod. 2001, 16, 2418–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, C.C.; McElin, T.W.; Manalo-Estrella, P. The elusive adenomyosis of the uterus-revisited. Am. J. Obstet. Gynecol. 1972, 112, 583–593. [Google Scholar] [CrossRef]
- Templeman, C.; Marchall, S.F.; Ursun, G.; Horn-Ross, P.L.; Clarke, C.A.; Allen, M.; Deapen, D.; Ziogas, A.; Reynolds, P.; Cress, R.; et al. Adenomyosis and endometriosis in the California Teachers Study: Reproductive and lifestyle correlates. Fertil. Steril. 2008, 90, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean-Baptiste, H.; Tetrokalashvili, M.; Williams, T.; Fogel, J.; Hsu, C.D. Characteristics associated with postoperative diagnosis of adenomyosis or combined adenomyosis with fibroids. Int. J. Gynecol. Obstet. 2013, 122, 112–114. [Google Scholar] [CrossRef]
- Morassutto, C.; Monasta, L.; Ricci, G.; Barbone, F.; Ronfani, L. Incidence and estimated prevalence of endometriosis and adenomyosis in Northeast Italy, a data linkage study. PLoS ONE 2016, 11, e0154227. [Google Scholar] [CrossRef] [Green Version]
- Shafrir, A.L.; Farland, L.V.; Shah, D.K.; Harris, H.R.; Kvaskoff, M.; Zondervan, K.; Missmer, S.A. Risk for and consequences of endometriosis, a critical epidemiologic review. Best Pract. Res. Clin. Obstet. Gynecol. 2018, 51, 1–15. [Google Scholar] [CrossRef]
- Taran, F.A.; Stewart, E.A.; Brucker, S. Adenomyosis, epidemiology, risk factors, clinical phenotypes and surgical and interventional alternatives to hysterectomy. Geburtshilfe Frauenheilkd. 2013, 73, 924–931. [Google Scholar] [CrossRef] [Green Version]
- Parazzini, F.; Vercellini, P.; Panazza, S.; Chatenoud, L.; Oldani, S.; Crosignani, P.G. Risk factors for adenomyosis. Hum. Reprod. 1997, 12, 1275–1279. [Google Scholar] [CrossRef] [Green Version]
- Taran, F.A.; Wallwiener, M.; Kabashi, D.; Rothmund, R.; Rall, K.; Kraemer, R.; Brucker, S.Y. Clinical characteristics indicating adenomyosis at the time of hyterectomy: A retrospective study in 291 patients. Arch. Gynecol. Obstet. 2012, 285, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Dada, O.A.; Laditan, A.A. Circulating hormonal levels during proloned lactational amenorrhea. Clin. Chim. Acta 1982, 123, 287–292. [Google Scholar] [CrossRef]
- Parazzini, F.; Mais, V.; Cipriani, S.; Busacca, M.; Venturini, P.; on behalf of GISE. Determinants of adenomyosis in women who underwent hysterectomy for benign gynecological conditions: Results from a prospective multicenter study in Italy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 143, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.; Obrant, K.J.; Karlsson, M. Pregnancy and lactation confer reversible bone loss in humans. Osteoporos. Int. 2001, 12, 828–834. [Google Scholar] [CrossRef]
- Levgur, M.; Abadi, M.A.; Tucker, A. Adenomyosis, symptoms, histology and pregnancy terminations. Obstet. Gynecol. 2000, 95, 688–691. [Google Scholar] [CrossRef]
- Curtis, K.M.; Hillis, S.D.; Marchbanks, P.A.; Peterson, H.B. Disruption of endometrial-myometrial border during pregnancy as a risk factor for adenomyosis. Am. J. Obstet. Gynecol. 2002, 187, 543–544. [Google Scholar] [CrossRef]
- Panganamamula, U.R.; Harmanli, O.H.; Isik-Akbay, E.F.; Grotegut, C.A.; Dandolu, V.; Gaughan, J.P. Is prior uterine surgery a risk factor for adenomyosis? Obstet. Gynecol. 2004, 104, 1034–1038. [Google Scholar] [CrossRef]
- McCluggage, W.G.; Desai, V.; Manek, S. Tamoxifen-associated postmenopausal adenomyosis exhibits stromal fibrosis, glandular dilatation, and epithelial metaplasias. Histopathology 2000, 37, 340–346. [Google Scholar] [CrossRef]
- Cohen, I.; Beyth, Y.; Shapira, J.; Tepper, R.; Fishman, A.; Cordoba, M.; Bernheim, J.; Yigael, D.; Altaras, M.M. High frequency of adenomyosis in postmenopausal breast cancer patients treated with tamoxifen. Gynecol. Obstet. Investig. 1997, 44, 200–205. [Google Scholar] [CrossRef]
- Varras, M.; Polyzos, D.; Akrivis, C. Effects of tamoxifen on the human female genital tract: Review of the literature. Eur. J. Gynecol. Oncol. 2003, 24, 258–268. [Google Scholar]
- Huang, P.C.; Tsai, E.M.; Li, W.F.; Liao, P.C.; Chung, M.C.; Wang, Y.H.; Wang, S.L. Association between phthalate exposure and glutathione S-transferase M1 polymorphism in adenomyosis, leiomyoma and endometriosis. Hum. Reprod. 2010, 25, 986–994. [Google Scholar] [CrossRef] [Green Version]
- Yen, C.F.; Huang, S.J.; Lee, C.L.; Wang, H.S.; Liao, S.K. Molecular characteristics of the endometrium in uterine adenomyosis and its biochemical microenvironment. Reprod. Sci. 2017, 24, 1346–1361. [Google Scholar] [CrossRef] [PubMed]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, S.W. Aberrant immunoreactivity of deoxyribonucleic acid methyltransferases in adenomyosis. Gynecol. Obstet. Investig. 2012, 74, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Setoguchi, T. Histology of endometrium. In Practical Histology, 1st ed.; Kanehara Suppan: Tokyo, Japan, 1979; pp. 228–229. (In Japanese) [Google Scholar]
- Brosens, J.J.; de Souza, N.M.; Barker, F.G. Uterine junctional zone: Function and disease. Lancet 1995, 346, 558–560. [Google Scholar] [CrossRef]
- Naftalin, J.; Jurkovic, D. The endometrial-myometrial junction: A fresh look at a busy crossing. Ultrasound Obstet. Gynecol. 2009, 34, 1–11. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, L.; Li, T.C.; Duan, H.; Yu, P.; Wang, H.Y. Ultrastructural features of endometrial-myometrial interface and its alteration in adenomyosis. Int. J. Clin. Exp. Pathol. 2014, 7, 1469–1477. [Google Scholar]
- Hricak, H.; Alpers, C.; Crooks, L.E.; Sheldon, P.E. Magnetic resonance imaging of the female pelvis: Initial experience. Am. J. Roentgenol. 1983, 141, 1119–1128. [Google Scholar] [CrossRef]
- Mehasseb, M.K.; Bell, S.C.; Brown, L.; Pringle, J.H.; Habiba, M. Phenotypic characterization of the inner and outer myometrium in normal and adenomyotic uteri. Gynecol. Obstet. Investig. 2011, 71, 217–224. [Google Scholar] [CrossRef]
- Tetlow, R.L.; Richmond, I.; Manton, D.J.; Greeman, J.; Turnbull, L.W.; Killick, S.R. Histological analysis of the uterine junctional zone as seen by transvaginal ultrasound. Ultrasound Obstet. Gynecol. 1999, 14, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Hoad, C.L.; Raine-Fenning, N.J.; Fulford, J.; Campbell, B.K.; Johnson, I.R.; Gowland, P.A. Uterine tissue development in healthy women during the normal menstrual cycle and investigations with magnetic resonance imaging. Am. J. Obstet. Gynecol. 2005, 192, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Noe, M.; Kunz, G.; Herbertz, M.; Mall, G.; Leyendecker, G. The cyclic pattern of the immunocytochemical expression of estrogen and progesterone receptors in human myometrial and endometrial layers: Characterization of endometrial-subendometrial unit. Hum. Reprod. 1999, 14, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Anguilar, H.N.; Mitchell, B.F. Physiological pathways and molecular mechanisms regulating uterine contractility. Hum. Reprod. Update 2010, 16, 725–744. [Google Scholar] [CrossRef] [Green Version]
- Kurowicks, B.; Franczak, A.; Oponowicz, A.; Kotwica, G. In vitro contractile activity of porcine myometrium during luteolysis and early pregnancy: Effect of oxytocin and progesterone. Reprod. Biol. 2005, 5, 151–169. [Google Scholar]
- Kunz, G.; Beil, D.; Deininger, H.; Wildt, L.; Leyendecker, G. The dynamics of rapid sperm transport through the female genital tract: Evidence from vaginal sonography of uterine peristalsis and hysterosalpingoscintigraphy. Hum. Reprod. 1996, 11, 627–632. [Google Scholar] [CrossRef]
- De Vries, K.; Lyons, E.A.; Ballard, G.; Levi, C.S.; Lindsay, D.J. Contractions of the inner third of the myometrium. Am. J. Obstet. Gynecol. 1990, 162, 679–682. [Google Scholar] [CrossRef]
- Lyons, E.A.; Taylor, P.J.; Zheng, X.H.; Ballard, G.; Levi, C.S.; Kredentser, J.V. Characterization of subendmetrial myometrial contractions throughout the menstrual cycle in normal fertile women. Fertil. Steril. 1991, 55, 771–774. [Google Scholar] [CrossRef]
- Leyendecker, G.; Wildt, L.; Mall, G. The pathophysiology of endometriosis and adenomyosis: Tissue injury and repair. Arch. Gynecol. Obstet. 2009, 280, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Meenakshi, M.; McCluggage, W.G. Vascular involvement in adenomyosis: Report of a large series of of a common phenomenon with observations on the pathogenesis of adenomyosis. Int. J. Gynecol. Pathol. 2010, 29, 117–121. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Ussia, A.; Adamyan, L.; Wattiez, A.; Gomel, V.; Martin, D.C. Pathogenesis of endometriosis: The genetic and epigenetic theory. Fertil. Steril. 2019, 111, 327–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitawaki, J.; Noguchi, T.; Amatsu, T.; Maeda, K.; Tsukamoto, K.; Yamamoto, T.; Fushiki, S.; Osawa, Y.; Honjo, H. Expression of aromatase cytochrome P450 protein and messenger ribonucleic acid in human endometriotic and adenomyotic tissues but not in normal endometrium. Biol. Reprod. 1997, 57, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitawaki, J.; Koshiba, H.; Ishihara, H.; Kusuki, I.; Tsukamoto, K.; Honju, H. Progesterone induction of 17beta-hydroxysteroid dehydrogenase type 2 during the secretory phase occurs in the endometrium of estrogen-dependent benign diseases but not in normal endometrium. J. Clin. Endocrinol. Metab. 2000, 85, 3292–3296. [Google Scholar] [PubMed] [Green Version]
- Mehasseb, M.K.; Panchal, R.; Taylor, A.H.; Brown, L.; Bell, S.C.; Habiba, M. Estrogen and progesterone receptor isoform distribution through the menstrual cycle in uteri with and without adenomyosis. Fertil. Steril. 2011, 95, 2228–2235.e1. [Google Scholar] [CrossRef] [PubMed]
- Jichan, N.; Xishi, L.; Guo, S.W. Promoter hypermethylation of progesterone receptor isoform B (PR-B) in adenomyosis and its rectification by a histone deacetylase inhibitor and a demethylation agent. Reprod. Sci. 2010, 17, 995–1005. [Google Scholar]
- Leyendecker, G.; Wildt, L. A new concept in endometriosis and adneomyosis: Tissue injury and repair (TIAR). Horm. Mol. Biol. Clin. Investig. 2011, 5, 125–142. [Google Scholar] [PubMed] [Green Version]
- Shaked, S.; Jaffa, A.J.; Grisaru, D.; Elad, D. Uterine peristalsis-induced stresses with the uterine wall may sprout adenomyosis. Biomed. Model. Mechano-Biol. 2015, 14, 437–444. [Google Scholar] [CrossRef]
- Samenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Andrikopoulou, E.; Zhang, X.; Sebastian, R.; Marti, G.; Liu, L.; Milner, S.M.; Harmon, J.W. Current insights into the role of HIF-1 in cutaneous wound healing. Curr. Mol. Med. 2011, 11, 218–235. [Google Scholar] [CrossRef]
- Hong, W.X.; Hu, M.S.; Esquivel, M.; Liang, G.Y.; Rennert, R.C.; McArdle, A.; Paik, K.J.; Duscher, D.; Gurtner, G.C.; Lorenz, H.P.; et al. The role of hypoxia-inducible factor in wound healing. Adv. Wound Care 2014, 3, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Guo, S.W.; Liu, X. Activated platelets induce hypoxia-inducible factor-1α expression likely through transforming growth factor-β1 in human endometrial stromal cells. Reprod. Dev. Med. 2019, 3, 69–76. [Google Scholar]
- Gear, A.R.; Camerini, D. Platelet chemokines and chemokine receptors, linking hemostasis, inflammation, and host defense. Microcirculation 2003, 10, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.D.; Burger, P.C. Platelets in inflammation and thrombosis. Arter. Thromb. Vasc. Biol. 2003, 23, 2131–2137. [Google Scholar] [CrossRef] [PubMed]
- Horsythe, J.A.; Jiang, B.H.; Iyer, N.V.; Agani, F.; Leung, S.W.; Koos, R.D.; Samenza, G.L. Activation of vascular endothelial growth factor gene expression by hypoxia-inducible factor 1. Mol. Cell. Biol. 1996, 16, 4604–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, E.D. Organization and fine structure of epithelium and mesenchyme in the development of chick embryo. In Epithelial-Mesenchymal Interactions; Fleischmajer, R., Billingham, R.E., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1968; pp. 31–55. [Google Scholar]
- Shapiro, L.; Fannon, A.M.; Kwong, P.D.; Thompson, A.; Lehmann, M.S.; Grubel, G.; Legrand, J.F.; Als-Nielsen, J.; Colman, D.R.; Hendrickson, W.A. Structural basis of cell-cell adhesion by cadherins. Nature 1995, 374, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Gaetje, R.; Kotzian, S.; Herrmann, G.; Baumann, R.; Starzinski-Powitz, A. Nonmalignant epithelial cells, potentially invasive in human endometriosis, lacks the tumor suppressor molecule E-cadherin. Am. J. Pathol. 1997, 150, 461–467. [Google Scholar] [PubMed]
- Hiscox, S.; Jiang, W.G. HGF/SF regulates the phosphorylation of β-catenin and cell-cell adhesion in cancer cells. Proc. Am. Assoc. Cancer Res. 1998, 39, 500–501. [Google Scholar]
- Chen, Y.J.; Li, H.Y.; Huang, C.H.; Twu, N.F.; Yen, M.S.; Wang, P.H.; Chou, T.Y.; Liu, Y.N.; Chao, K.C.; Yang, M.H. Estrogen-induced epithelial-mesenchymal transition of endometrial epithelial cells contributes to the development of adenomyosis. J. Pathol. 2010, 222, 261–270. [Google Scholar]
- Park, S.H.; Cheung, L.W.T.; Wong, A.S.T.; Leung, P.C.K. Estrogen regulates Snail and Slug in the down-regulation of E-cadherin and induces mestastatic potential of ovarian cancer cells through estrogen receptor α. Mol. Endocrinol. 2008, 22, 2085–2098. [Google Scholar] [CrossRef]
- Asea, A.; Kraftt, S.K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Fingerg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependent pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Asea, A.; Rehli, M.; Kabingu, E.; Boch, J.A.; Bare, O.; Auron, P.E.; Stevenson, M.A.; Calderwood, S.K. Novel signal transduction pathway utilized by extracellular HSP70: Role of toll-like receptor (TLR) 2 and TLR4. J. Biol. Chem. 2002, 277, 15028–15034. [Google Scholar] [CrossRef] [Green Version]
- Matzinger, P. An innate sense of danger. Semin. Immunol. 1998, 10, 399–415. [Google Scholar] [CrossRef]
- Zugel, U.; Kaufmann, S.H. Innate response against heat-shock proteins in infectious diseases. Immunobiology 1999, 201, 22–35. [Google Scholar] [CrossRef]
- Khan, K.N.; Kitajima, M.; Immamura, T.; Hiraki, K.; Fujishita, A.; Sekine, I.; Ishimaru, T.; Masuzaki, H. Toll-like receptor 4 (TLR4)-mediated growth of endometriosis by human heat shock protein 70 (HSP70). Hum. Reprod. 2008, 23, 2210–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Ysuneo Inoue Seiko Tateishi Fujishita, A.; Nakashima, M.; Masuzaki, H. Additive effects of inflammation and stress reaction on Toll-like receptor 4-mediated growth of endometriotic stromal cells. Hum. Reprod. 2013, 28, 2794–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Sekine, I.; Ishimaru, T.; Masuzaki, H. Immunopathogenesis of pelvic endometriosis: Role of hepatocyte growth factor, macrophages and ovarian steroids. Am. J. Reprod. Immunol. 2008, 60, 383–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Nakashima, M.; Masuzaki, H. Decreased expression of human heat shock protein 70 in the endometria and pathological lesions of women with adenomyosis and uterine myoma after GnRHa agonist. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 187, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Zeitvogel, A.; Baumann, R.; Starzinski-Powitz, A. Identification of an invasive, N-cadherin-expressing epithelial cell type in endometriosis using a new cell culture model. Am. J. Pathol. 2001, 159, 1839–1852. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Shen, M.; Qi, Q.; Zhang, H.; Guo, S.W. Corroborating evidence of for platelet-induced epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the development of adenomyosis. Hum. Reprod. 2016, 31, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Acloque, H.; Adams, M.S.; Fishwick, K.; Bronner-Fraser, M.; Nieto, M.A. Epithelial-mesenchymal transitions: The importance of changing cell state in development and disease. J. Clin. Investig. 2009, 119, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Masuzaki, H.; Fujishita, A.; Kitajima, M.; Sekine, I.; Matsuyama, T.; Ishimaru, T. Estrogen and progesterone receptor expression in macrophages and regulation of hepatocyte growth factor by ovarian steroids in women with endometriosis. Hum. Reprod. 2005, 20, 2004–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Fujishita, A.; Ishimaru, T.; Sekine, I.; Masuzaki, H. Multifunctional role of hepatocyte growth factor in the development of pelvic endometriosis. Am. J. Reprod. Immunol. 2008, 60, 383–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobel, V.; Zhu, Y.S.; Imperato-McGinley, J. Fetal hormones and sexual differentiation. Obstet. Gynecol. Clin. N. Am. 2004, 31, 837–856. [Google Scholar] [CrossRef]
- Spencer, T.E.; Hayashi, K.; Hu, J.; Carpenter, K.D. Comparative developmental biology of the mammalian uterus. Curr. Top. Dev. Biol. 2005, 68, 85–122. [Google Scholar] [PubMed]
- Donnez, J.; Nisole, M.; Casanas-Roux, F.; Brion, P.; Da Costa Ferreira, N. Stereometric evaluation of peritoneal endometriosis and endometriotic nodules of the rectovaginal septum. Hum. Reprod. 1996, 11, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Donnez, J.; Nisolle, M.; Gillerot, S.; Smets, M.; Bassil, S.; Casanas-Roux, F. Rectovaginal septum adenomyotic nodules: A series of 500 cases. Br. J. Obstet. Gynecol. 1997, 104, 1014–1018. [Google Scholar] [CrossRef]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometrioma, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Enatsu, A.; Harada, T.; Yoshida, S.; Iwabe, T.; Terakawa, N. Adenomyosis in a patient with the Rokitansky-Kuster-Hauser syndrome. Fertil. Steril. 2000, 73, 862–863. [Google Scholar] [CrossRef]
- Hoo, P.S.; Norhaslinda, A.R.; Reza, J.N. Rare case of leiomyoma and adenomyosis in Rokitansky-Kuster-Hauser syndrome. Case Rep. Obstet. Gynecol. 2016, 2016, 3725043. [Google Scholar] [CrossRef] [Green Version]
- Schwab, K.E.; Gargett, C.E. Co-expression of two perivascular cell markers isolates mesenchymal stem-like cells from human endometrium. Hum. Reprod. 2007, 22, 2903–2911. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Anwar, S.S.; Buhring, H.J.; Rao, J.R.; Gargett, C.E. A novel marker of human endometrial mesenchymal stem-like cells. Cell Transplant. 2012, 21, 2201–2214. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Schwab, K.E.; Zillwod, R.M.; Nguyen, H.P.; Wu, D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Hum. Reprod. 2009, 80, 1136–1145. [Google Scholar]
- Chan, R.W.; Schwab, K.E.; Gargett, C.E. Clonogenicity of human endometrial epithelial and stromal cells. Biol. Reprod. 2004, 70, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E. Uterine stem cells: What is the evidence? Hum. Reprod. Update 1998, 4, 312–322. [Google Scholar] [CrossRef]
- Okulicz, W.C.; Ace, C.I.; Scarrell, R. Zonal changes in proliferation in the rhesus endometrium during the late secretory phase and menses. Proc. Soc. Exp. Biol. Med. 1997, 214, 132–138. [Google Scholar] [CrossRef]
- Schwab, K.E.; Chan, R.W.; Gargett, C.E. Putative stem cell activity of human endometrial epithelial and stromal cells during the menstrual cycle. Fertil. Steril. 2005, 84 (Suppl. S2), 1124–1130. [Google Scholar] [CrossRef]
- Hufnagel, D.; Li, F.; Cosar, E.; Krikun, G.; Taylor, H.S. The role of stem cells in the etiology and pathophysiology of endometriosis. Semin. Reprod. Med. 2015, 33, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannuccini, S.; Tosti, C.; Carmona, F.; Huang, S.I.; Chapron, C.; Guo, S.W.; Petraglia, F. Pathogenesis of adenomyosis: An update on molecular mechanisms. Reprod. Biomed. Online 2017, 35, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Li, Y.G.; Pu, D.M. Matrix metalloproteinase-2 and -9 expression correlated with angiogenesis in human adenomyosis. Gynecol. Obstet. Investig. 2006, 62, 229–235. [Google Scholar] [CrossRef]
- Yi, K.W.; Kim, S.H.; Ihm, H.J.; Sang, Y.; Chae, H.D.; Kim, C.-H.; Kang, B.M. Increased expression of p21-activated kinase 4 in adenomyosis and its regulation of matrix metalloproteinase-2 and -9 in endometrial cells. Fertil. Steril. 2015, 103, 1089–1097. [Google Scholar] [CrossRef]
- Taylor, H.S.; Osteen, K.G.; Bruner-Tran, K.L.; Lockwood, C.J.; Krikun, G.; Sokalska, A.; Duleba, A.J. Novel therapies targeting endometriosis. Reprod. Sci. 2011, 18, 814–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Taylor, H.S. Stem cells and female reproduction. Reprod. Sci. 2009, 16, 126–139. [Google Scholar] [CrossRef] [Green Version]
- Leyendecker, G.; Herbeertz, M.; Kunz, G.; Mali, G. Endometriosis results from the dislocation of basalis endometrium. Hum. Reprod. 2002, 17, 2725–2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.G.; Chiantera, V.; Frangini, S.; Younes, S.; Kohler, C.; Taube, E.T.; Plendl, J.; Mechsner, S. Ultramicro-trauma in the endometrial-myometrial junctional zone and pale cell migration in adenomyosis. Fertil. Steril. 2015, 104, e1–e3. [Google Scholar] [CrossRef] [Green Version]
- Zeitoun, K.; Takayama, K.; Sasano, H.; Suzuki, T.; Moghtrabi, N.; Andersson, S.; Johns, A.; Meng, L.; Putman, M.; Carr, B.; et al. Deficient 17β-hydroxysteroid dehydrogenase type 2 expression in endometriosis: Failure to metabolize 17β-estradiol. J. Clin. Endocrinol. Metab. 1998, 83, 4474–4480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Tsukino, H.; Iwasaki, M.; Sasaki, H.; Tanaka, T.; Katoh, T.; Patterson, D.G., Jr.; Turner, W.; Needham, L.; Tsugane, S. Interaction between cytochrome P450 gene polymorphisms and serum serum organochlorine TEQ levels in the risk of endometriosis. Mol. Hum. Reprod. 2007, 13, 399–404. [Google Scholar] [CrossRef]
- Artymuk, N.; Zotova, O.; Gulyaeva, L. Adenomyosis: Genetics of estrogen metabolism. Horm. Mol. Biol. Clin. Investig. 2019, 37, 37. [Google Scholar] [CrossRef]
- Tong, X.; Li, Z.; Wu, Y.; Fu, X.; Zhang, Y.; Fan, H. COMT 158G/A and CYP1B1 432C/G polymorphisms increase the risk of endometriosis and adenomyosis: A meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 179, 17–21. [Google Scholar] [CrossRef]
- Guo, S.W. Epigenetics of endometriosis. Mol. Hum. Reprod. 2009, 15, 587–607. [Google Scholar] [CrossRef]
- Gordts, S.; Koninckx, P.; Brosens, I. Pathogenesis of deep endometriosis. Fertil. Steril. 2017, 108, 872–885.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Sun, Y.; Yang, B.; Yang, Y.; Zhang, Y.; Yu, T.; Huang, H.; Zhang, J.; Xu, H. Transcriptome sequencing of adenomyosis eutopic endometrium: A new insight into its pathophysiology. J. Cell. Mol. Med. 2019, 23, 8381–8391. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Nie, J.; Guo, S.W. Elevated immunoreactivity against class I histone deacetylases in adenomyosis. Gynecol. Obstet. Investig. 2012, 74, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, S.W. Valproic acid alleviates generalized hyperalgesia in mice with induced adenomyosis. J. Obstet. Gynecol. Res. 2011, 37, 696–708. [Google Scholar] [CrossRef] [PubMed]
- Anglesio, M.S.; Papadopoulos, N.; Ayhan, A.; Nazeran, T.M.; Noe, M.; Horlings, H.M.; Lum, A.; Jones, S.; Senz, J.; Seckin, T.; et al. Cancer-associated mutations in endometriosis without cancer. N. Engl. J. Med. 2017, 376, 1835–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, K.; Nakaoka, H.; Yoshihara, K.; Ishiguru, T.; Tamura, R.; Mori, Y.; Yamawaki, K.; Adachi, S.; Takahashi, T.; Kase, H.; et al. Clonal expansion and diversification of cancer-associated mutations in endometriosis and normal endometrium. Cell Rep. 2018, 24, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Hirota, Y.; Ueno, T.; Fukui, Y.; Yoshida, E.; Hayashi, T.; Kojima, S.; Takayama, R.; Hashimoto, T.; Kiyono, T.; et al. Uterine adenomyosis is an oligoclonal disorder associated with KRAS mutations. Nat. Commun. 2019, 10, 5785. [Google Scholar] [CrossRef]
- Makinen, N.; Mehine, M.; Tolvanen, J.; Kaasinen, E.; Li, Y.; Lehtonen, H.J.; Gentile, M.; Yan, J.; Enge, M.; Taipale, M.; et al. MED12, the mediator complex subunit 12 gene, is mutated at high frequency on uterine leiomyomas. Science 2011, 334, 252–255. [Google Scholar] [CrossRef]
- Mehine, M.; Kaasinen, E.; Makinen, N.; Katainen, R.; Kampjarvi, K.; Pitkanen, E.; Heinonen, H.R.; Butzow, R.; Kilpivaara, O.; Kuosmanen, A.; et al. Characterization of uterine leiomyomas by whole-genome sequencing. N. Engl. J. Med. 2013, 369, 43–53. [Google Scholar] [CrossRef]
- Moore, L.; Leongamornlert, D.; Coorens, T.H.H.; Sanders, M.A.; Ellis, P.; Dentro, S.C.; Dawson, K.J.; Butler, T.; Rahbari, R.; Mitchell, T.J.; et al. The mutational landscape of normal human endometrial epithelium. Nature 2020, 580, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.Y.; Wu, M.-H.; Tsai, S.-J. Epigenetic regulation of the pathological process in endometriosis. Reprod. Med. Biol. 2017, 16, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braza-Boils, A.; Mari-Alexandre, J.; Gilabert, J.; Sanchez-Izquierdo, D.; Espana, F.; Estelles, A.; Gilabert-Estelles, J. MicroRNA expression profile in endometriosis: Its relation to angiogenesis and fibrinolytic factors. Hum. Reprod. 2014, 29, 978–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, T.; Nasu, K.; Abe, W.; Aoyagi, Y.; Okamoto, M.; Kai, K.; Takebayashi, K.; Narahara, H. miR-503, a microRNA epigenetically expressed in endometriosis, induces apoptosis and cell-cycle arrest and inhibits cell proliferation, angiogenesis, and contractility of human ovarian endometriotic stromal cells. Hum. Reprod. 2016, 31, 2587–2597. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.A.; Bracken, C.P.; Bert, A.G.; Goodall, G.J. MicroRNAs as regulators of epithelial-mesenchymal transition. Cell Cycle 2008, 7, 3112–3118. [Google Scholar] [CrossRef]
- Bracken, C.P.; Gregory, P.A.; Khew-Goodall, Y.; Goodall, G.J. The role of microRNAs in metastasis and epithelial-mesenchymal transition. Cell. Mol. Life Sci. 2009, 66, 1682–1699. [Google Scholar] [CrossRef]
- Ohlsson Teague, E.M.C.; Print, C.G.; Hull, M.L. The role of microRNAs in endometriosis and associated reproductive conditions. Hum. Reprod. Update 2010, 16, 142–165. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Li, H.; He, Y. MicroRNA-17 downregulates expression of the PTEN gene to promote the occurrence and development of adenomyosis. Exp. Ther. Med. 2017, 14, 3805–3811. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Feldman, G.; Huang, J.; Wu, S.; Zhang, N.; Comeford, S.A.; Gayyed, M.F.; Andres, R.A.; Maitra, A.; Pan, D. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila homolog of YAP. Cell 2005, 122, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Hunag, J.-H.; Duan, H.; Wang, S.; Wang, Y.-Y.; Cheng-Xiao, L.V. Upregulated microRNA let-7a accelerates apoptosis and inhibits proliferation in uterine junctional zone smooth muscle cells in adenomyosis under conditions of a normal activated hippo-YAP1 axis. Reprod. Biol. Endocrinol. 2021, 19, 81. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Mutlu, L.; Grechukhina, O.; Taylor, H.S. Circulating microRNAs as potential biomarkers for endometriosis. Fertil. Steril. 2015, 103, 1252–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordts, S.; Brosens, J.J.; Fusi, L.; Benagiano, G.; Brosens, I. Uterine adenomyosis: A need for uniform terminology and consensus classification. Reprod. Biomed. Online 2008, 17, 244–248. [Google Scholar] [CrossRef]
- Pistofidis, G.; Makrakis, E.; Koukoura, O.; Bardis, N.; Balinakos, P.; Anaf, V. Distinct types of uterine adenomyosis based on laparoscopic and histologic criteria. Clin. Exp. Obstet. Gynecol. 2014, 41, 113–118. [Google Scholar] [CrossRef]
- Van den Bosch, T.; Dueholm, M.; Leone, F.P.; Valentin, L.; Rasmussen, C.K.; Votino, A.; Van Schoubroeck, D.; Landolfo, C.; Installe, A.J.; Guerriero, S.; et al. Terms, definitions and measurements to describe sonographic features of myometrium and uterine masses: A consensus opinion from the Morphological Uterus Sonographic Assessment (MUSA) group. Ultrasound Obstet. Gynecol. 2015, 46, 284–298. [Google Scholar] [CrossRef]
- Bazot, M.; Cortez, A.; Darai, E.; Roujer, J.; Chopier, J.; Antoine, J.M.; Uzan, S. Ultrasonography compared with magnetic resonance imaging for the diagnosis of adenomyosis: Correlation with histopathology. Hum. Reprod. 2001, 16, 2427–2433. [Google Scholar] [CrossRef] [Green Version]
- Kishi, Y.; Suginami, H.; Kumamori, R.; Yabuta, M.; Suginami, R.; Taniguchi, F. Four syptypes of adenomyosis assessed by magnetic resonance imaging and their specification. Am. J. Obstet. Gynecol. 2012, 207, 114.e1–114.e7. [Google Scholar] [CrossRef]
- Khan, K.N.; Fujishita, A.; Koshiba, A.; Kuroboshi, H.; Mori, T.; Ogi, H.; Itoh, K.; Nakashima, M.; Kitawaki, J. Biological differences between intrinsic and extrinsic adenomyosis with coexisting deep infiltrating endometriosis. Reprod. Biomed. Online 2019, 39, 343–353. [Google Scholar] [CrossRef]
- Bazot, M.; Darai, E. Role of transvaginal sonography and magnetic resonance imaging in the diagnosis of uterine adenomyosis. Fertil. Steril. 2018, 109, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Fedele, L.; Bianchi, S.; Frontino, G. Hormonal treatments for adenomyosis. Best Pract. Res. Clin. Obstet. Gynecol. 2008, 22, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Pontis, A.; D’Alterio, M.N.; Pirarba, S.; de Angelis, C.; Tinelli, R.; Angioni, S. Adenomyosis: A systematic review of medical treatment. Gynecol. Endocrinol. 2016, 32, 696–700. [Google Scholar] [CrossRef]
- Osuga, Y.; Fujimoto-Okabe, H.; Hagino, A. Evaluation of the efficacy and safety of dienogest in the treatment of painful symptoms in patients with adenomyosis: A randomized, double-blind, multicenter, placebo-controlled study. Fertil. Steril. 2017, 108, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Fujishita, A.; Koshiba, A.; Mori, T.; Kuroboshi, H.; Ogi, H.; Itoh, K.; Nakashima, M.; Kitawaki, J. Biological differences between focal and diffuse adenomyosis and response to hormonal treatment. Reprod. Biomed. Online 2019, 38, 634–646. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, X.; Critchley, H.; Fu, Z.; Guo, S.W. How does the extent of fibrosis in adenomyosis lesions contribute to heavy menstrual bleeding? Reprod. Med. Biol. 2022, 21, e12442. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Sekine, I.; Ishimaru, T.; Masuzaki, H. Changes in tissue inflammation, angiogenesis and apoptosis in endometriosis, adenomyosis and uterine myoma after GnRH agonist therapy. Hum. Reprod. 2010, 25, 642–653. [Google Scholar] [CrossRef]
- Kissler, S.; Zangos, S.; Wiegratz, I.; Kohl, J.; Rody, A.; Gaetje, R.; Doebert, N.; Wildt, L.; Kunz, G.; Leyendecker, G. Utero-tubal sperm transport and its impairment in endometriosis and adenomyosis. Ann. N. Y. Acad. Sci. 2007, 1101, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Fujishita, A.; Suematsu, T.; Ogawa, K.; Koshiba, A.; Mori, T.; Itoh, K.; Teramukai, S.; Matsuda, K.; Nakashima, M.; et al. An axonemal alteration in apical endometria of human adenomyosis. Hum. Reprod. 2021, 36, 1574–1589. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Martin, D.C. Deep endometriosis: A consequence of infiltration or retraction or possibly adenomyosis externa? Fertil. Steril. 1992, 58, 924–928. [Google Scholar] [CrossRef]
- Hasanpoor-Azghdy, S.B.; Simber, M.; Vedadhir, A. The emotional-psychological consequences of infertility among infertile women seeking treatment: Results of a qualitative study. Iran. J. Reprod. Med. 2014, 12, 131–138. [Google Scholar]
- ESHRE Capri Workshop Group. Economic aspects of infertility care: A challenge for researchers and clinicians. Hum. Reprod. 2015, 30, 2243–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njagi, P.; Groot, W.; Arsenijevic, J.; Dyer, S.; Mburu, G.; Kiarie, J. Economic costs of infertility care for patients in low-income and middle-income countries: A systematic review protocol. BMJ Open 2020, 10, e042951. [Google Scholar] [CrossRef] [PubMed]
- Bulletti, C.; Coccia, M.E.; Battistoni, S.; Borini, A. Endometriosis and infertility. J. Assist. Reprod. Genet. 2010, 27, 441–447. [Google Scholar] [CrossRef]
- Urman, B.; Yakin, K. Ovulatory disorders and infertility. J. Reprod. Med. 2006, 51, 267–282. [Google Scholar]
- Briceag, I.; Costache, A.; Purcarea, V.L.; Cergan, R.; Dumitru, M.; Briceag, I.; Sajin, M.; Ispas, A.T. Current management of tubal infertility: From hysterosalpingography to ultrasonography and surgery. J. Med. Life 2015, 8, 157–159. [Google Scholar]
- Kairo, B.N. Impaired fertility caused by endocrine dysfunction in women. Endocrinol. Metab. Clin. N. Am. 2003, 32, 573–592. [Google Scholar]
- Lessey, B.A.; Palomino, W.A.; Apparao, K.B.C.; Young, S.L.; Lininger, R.A. Estrogen-receptor-alpha (ER-alpha) and defects in uterine receptivity in women. Reprod. Biol. Endocrinol. 2006, 4, S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremellen, K.P.; Russell, P. The distribution of immune cells and macrophages in the endometrium of women with recurrent reproductive failure II: Adenomyosis and macrophages. J. Reprod. Immunol. 2012, 93, 58–63. [Google Scholar] [CrossRef]
- Zegars-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The international glossary on infertility and fertility care, 2017. Hum. Reprod. 2017, 32, 1786–1801. [Google Scholar]
- Prescott, J.; Farland, L.V.; Tobias, D.K.; Gaskins, A.J.; Spiegeiman, D.; Chavarro, J.E.; Rich-Edwards, J.W.; Barbieri, R.L.; Missmer, S.A. A prospective cohort study of endometriosis and subsequent risk of infertility. Hum. Reprod. 2016, 7, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Dueholm, M.; Lundorf, E.; Hansen, E.S.; Sorensen, J.S.; Ledertoug, S.; Olesen, F. Magnetic resonance imaging and transvaginal ultrasonography for the diagnosis of adenomyosis. Fertil. Steril. 2001, 76, 588–594. [Google Scholar] [CrossRef]
- Devlieger, R.; D’Hooghe, T.; Timmerman, D. Uterine adenomyosis in the infertility clinic. Hum. Reprod. Update 2003, 9, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pervez, S.N.; Javed, K. Adenomyosis among samples from hysterectomy due to abnormal uterine bleeding. J. Ayub Med. Coll. Abbottabad 2013, 25, 68–70. [Google Scholar] [PubMed]
- De Souza, N.M.; Brosens, J.J.; Schwieso, J.E.; Paraschos, T.; Winston, R.M. The potential value of magnetic resonance imaging in infertility. Clin. Radiol. 1995, 50, 75–79. [Google Scholar] [CrossRef]
- Tomassetti, C.; Meuleman, C.; Timmerman, D.; D’Hooghe, T. Adenomyosis and subfertility: Evidence of association and causation. Semin. Reprod. Med. 2013, 31, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Barrier, B.F.; Malinowski, M.J.; Dick, E.J., Jr.; Hubbard, G.B.; Bates, G.W. Adenomyosis in the baboon is associated with primary infertility. Fertil. Steril. 2004, 82 (Suppl. S3), 1091–1094. [Google Scholar] [CrossRef]
- Martínez-Conejero, J.A.; Morgan, M.; Montesinos, M.; Fortunio, S.; Meseguer, M.; Simon, C.; Horcajadas, J.; Pellicer, A. Adenomyosis does not affect implantation, but is associated with miscarriage in patients undergoing oocyte donation. Fertil. Steril. 2011, 96, 943–950. [Google Scholar] [CrossRef]
- Younes, G.; Tulandi, T. Effects of adenomyosis on in vitro fertilization treatment outcomes: A meta-analysis. Fertil. Steril. 2017, 108, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Khine, Y.M.; Kaponis, A.; Nikellis, T.; Decavalas, G.; Taniguchi, F. The impact of adenomyosis on women’s fertility. Obstet. Gynecol. Surv. 2016, 71, 557–568. [Google Scholar] [CrossRef]
- Kunz, G.; Beil, D.; Hupert, P.; Leyendecker, G. Structural abnormalities of the uterine wall in women with endometriosis and infertility visualized by vaginal sonography and magnetic resonance imaging. Hum. Reprod. 2000, 15, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Brosens, J.; Verhoeven, H.; Campo, R.; Gianaroli, L.; Gordt, S.; Hazekamp, J.; Hagglund, L.; Mardesic, T.; Varila, E.; Zech, J.; et al. High endometrial aromatase P450 mRNA expression is associated with poor IVF outcome. Hum. Reprod 2004, 19, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Noda, Y.; Matsumoto, H.; Umaoka, Y.; Tatsumi, K.; Kishi, J.; Mori, T. Involvement of superoxide radicals in the mouse two-cell block. Mol. Reprod. Dev. 1991, 28, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.P.; Kayisili, U.; Taylor, H.S. HOXA10 expression is decreased in endometrium of women with adenomyosis. Fertil. Steril. 2011, 95, 1133–1136. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Fujishita, A.; Ogawa, K.; Koshiba, A.; Mori, T.; Itoh, K.; Nakashima, M.; Kitawaki, J. Occurrence of chronic endometritis in different types of human adenomyosis. Reprod. Med. Biol. 2022, 1, e12421. [Google Scholar] [CrossRef]
- McQueen, D.B.; Perfetto, C.O.; Hazard, F.K.; Lathi, R.B. Pregnancy outcomes in women with chronic endometritis and recurrent pregnancy loss. Fertil. Steril. 2015, 104, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Cicinelli, E.; Matteo, M.; Trojano, G.; Mitola, P.C.; Tinelli, R.; Vitagliano, A.; Crupano, F.M.; Lepera, A.; Miragliotta, G.; Resta, L. Chronic endometritis in patients with unexplained infertility: Prevalence and effects of antibiotic treatment on spontaneous conception. Am. J. Immunol. 2018, 79, e12782. [Google Scholar] [CrossRef]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Lepera, A.; Alfonso, R.; Indraccolo, U.; Marrocchella, S.; Greco, P.; Resta, L. Prevalence of chronic endometritis in repeated unexplained implantation failure and IVF success rate after antibiotic therapy. Hum. Reprod. 2015, 30, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, K.; Mastubayashi, H.; Yamaguchi, K.; Nishiyama, R.; Takaya, Y.; Ishikawa, T.; Yasuo, T.; Yamada, H. Chronic endometritis: Potential cause of infertility and obstetric and neonatal complications. Am. J. Reprod. Immunol. 2016, 75, 13–22. [Google Scholar] [CrossRef]
- Tamura, H.; Kishi, H.; Kitade, M.; Asai-Sato, M.; Tanaka, A.; Murakami, T.; Minegishi, T.; Sugino, N. Complications and outcomes of pregnant women with adenomyosis in Japan. Reprod. Med. Biol. 2017, 16, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Vercellini, P.; Bonfanti, I.; Berlanda, N. Adenomyosis and infertility: Is there a causal link? Exp. Rev. Endocrinol. Metab. 2019, 14, 365–367. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Vanni, V.S.; Bartiromo, L.; Papaleo, E.; Ziberberg, E.; Candiani, M.; Orvieto, R.; Vigano, P. Is the oocyte quality affected by endometriosis? A review of the literature. J. Ovarian Res. 2017, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, Y.C.; Shelton, J.B.; Laird, S.M.; Richmond, M.; Kudesia, G.; Li, T.C.; Ledger, W.L. IL-1, IL-6 and TNF-alpha concentrations in the peritoneal fluid of women with pelvic adhesions. Hum. Reprod. 2002, 17, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Masuzaki, H.; Fujishita, A.; Kitajima, M.; Sekine, I.; Ishimaru, T. Differential macrophage infiltration in early and advanced endometriosis and adjacent peritoneum. Fertil. Steril. 2004, 81, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Djahanbakhch, O.; Arian, S.; Carr, B.R. Tubal transport of gametes and embryos: In review of physiology and pathophysiology. J. Assist. Reprod. Genet 2014, 31, 1337–1347. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, R. Microtubules. In Standard Cell Biology, 1st ed.; Ishikawa, H., Kondo, H., Shibata, Y., Eds.; Igaku Shoin publishing Co.: Tokyo, Japan, 1999; pp. 137–151. (In Japanese) [Google Scholar]
- Satir, P. Mechanisms of ciliary movement: Contributions from electron microscopy. Scanning Microsc. 1992, 6, 573–579. [Google Scholar]
- Satir, P. The role of axonemal components in ciliary motility. Comp. Biochem. Physiol. A Comp. Physiol. 1989, 94, 351–357. [Google Scholar] [CrossRef]
- Satir, P. Structural basis of ciliary movement. Environ. Health Perspect. 1980, 35, 77–82. [Google Scholar] [CrossRef]
- Benaglia, L.; Cardellicchio, L.; Leonardi, M.; Faulisi, S.; Vercellini, P.; Paffoni, A.; Somigliana, E.; Fedele, L. Asymptomatic adenomyosis and embryo implantation in IVF cycles. Reprod. Biomed. Online 2014, 29, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Vercellini, P.; Consonni, D.; Dridi, D.; Bracco, B.; Frattaruolo, M.P.; Somigliana, E. Uterine adenomyosis and in vitro fertilization outcome: A systematic review and meta-analysis. Hum. Reprod. 2014, 29, 964–977. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Benagiano, G.; Liu, X.; Guo, S.W. Unveiling the pathogenesis of adenomyosis through animal models. J. Clin. Med. 2022, 11, 1744. [Google Scholar] [CrossRef]
- Khan, K.N.; Fujishita, A.; Kitajima, M.; Hiraki, K.; Nakashima, M.; Masuzaki, H. Occult microscopic endometriosis: Undetectable by laparoscopy in normal peritoneum. Hum. Reprod. 2014, 29, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, K.N.; Fujishita, A.; Mori, T. Pathogenesis of Human Adenomyosis: Current Understanding and Its Association with Infertility. J. Clin. Med. 2022, 11, 4057. https://doi.org/10.3390/jcm11144057
Khan KN, Fujishita A, Mori T. Pathogenesis of Human Adenomyosis: Current Understanding and Its Association with Infertility. Journal of Clinical Medicine. 2022; 11(14):4057. https://doi.org/10.3390/jcm11144057
Chicago/Turabian StyleKhan, Khaleque N., Akira Fujishita, and Taisuke Mori. 2022. "Pathogenesis of Human Adenomyosis: Current Understanding and Its Association with Infertility" Journal of Clinical Medicine 11, no. 14: 4057. https://doi.org/10.3390/jcm11144057