
Variation of Complement Protein Levels in Maternal Plasma and Umbilical Cord Blood during Normal Pregnancy: An Observational Study

 ,
,  ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Routine Laboratory Analyses
2.3. Complement Analyses
2.4. Statistics
2.5. Ethical Approval
3. Results
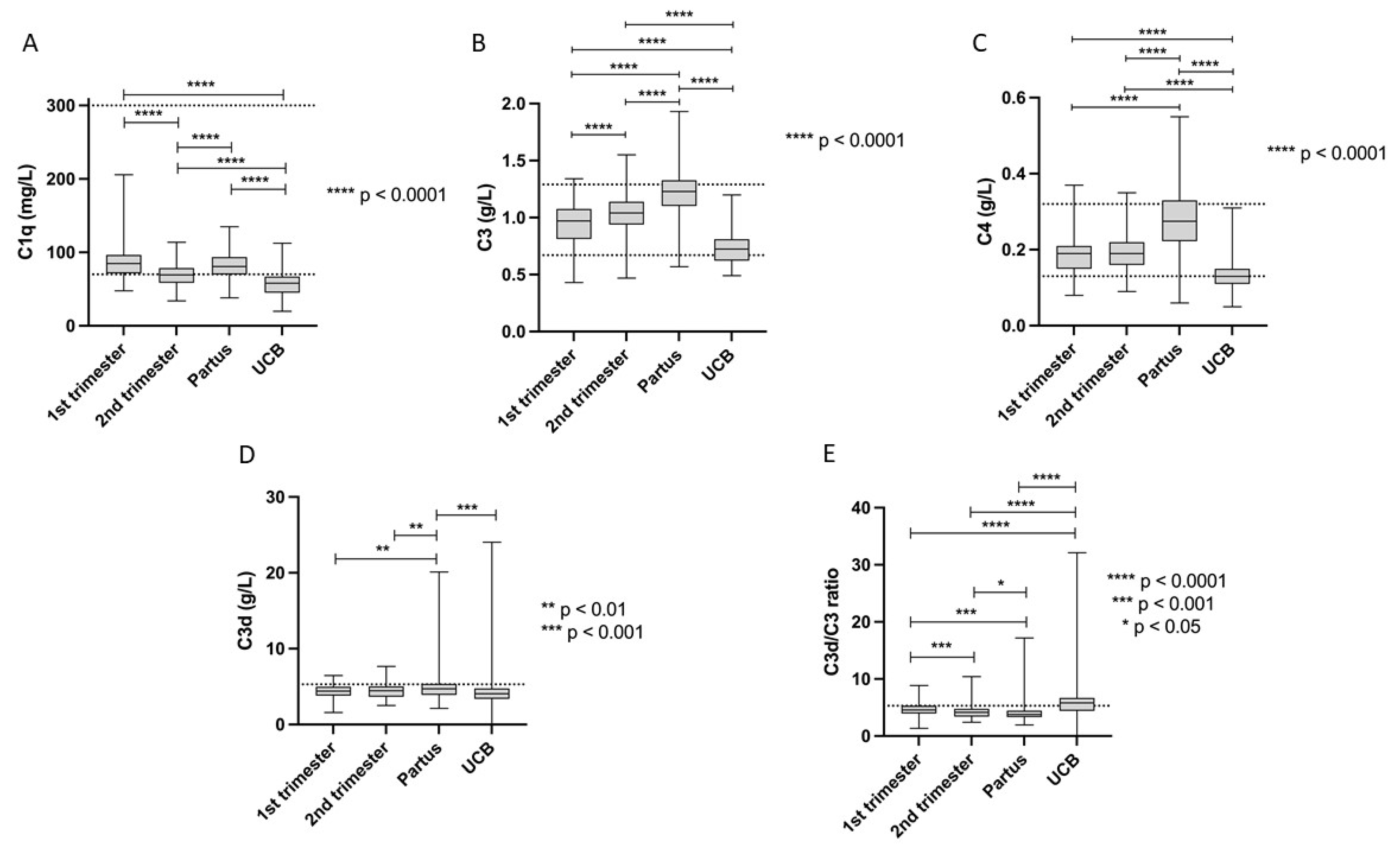
3.1. Complement Protein Levels during Pregnancy
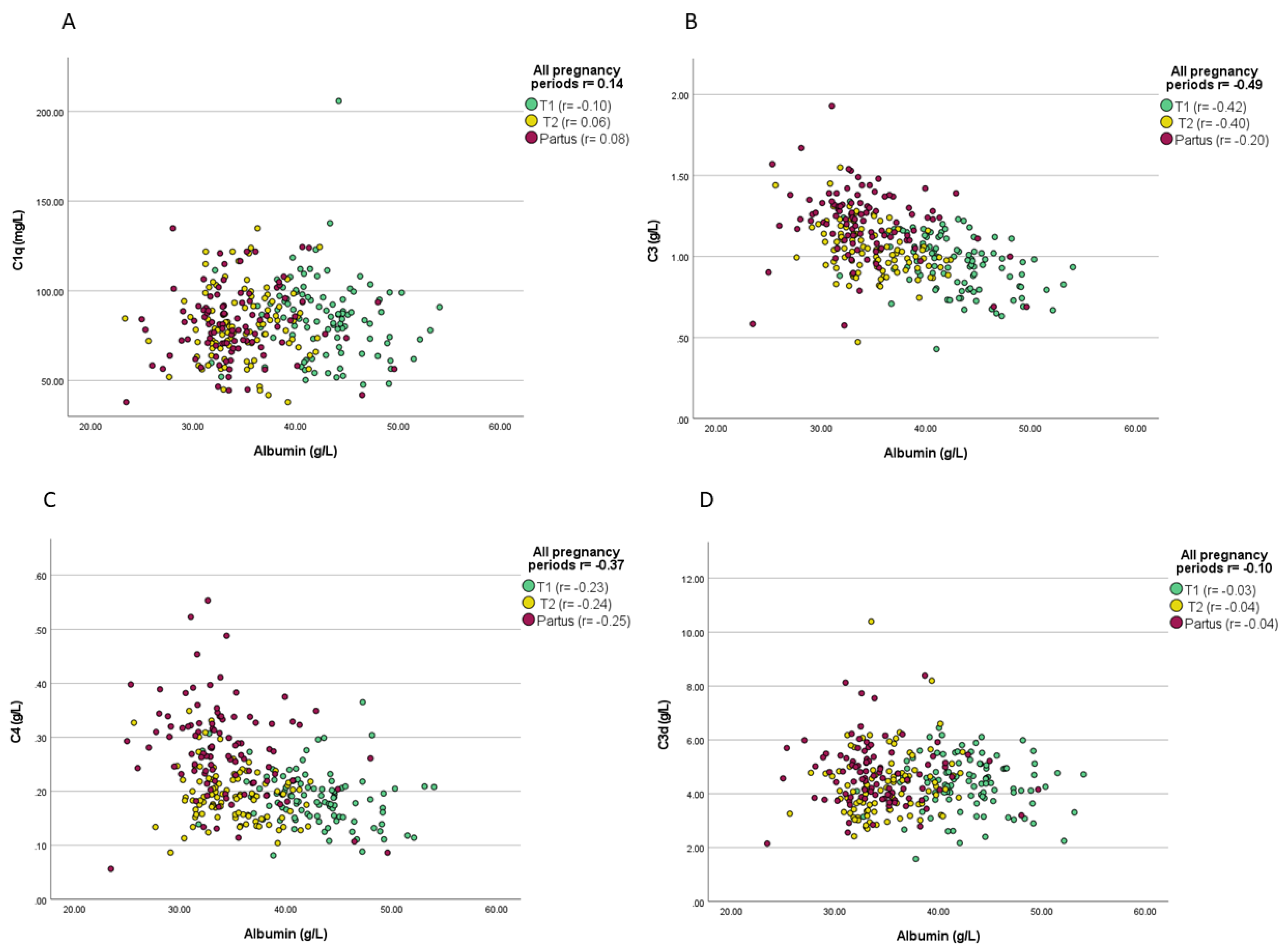
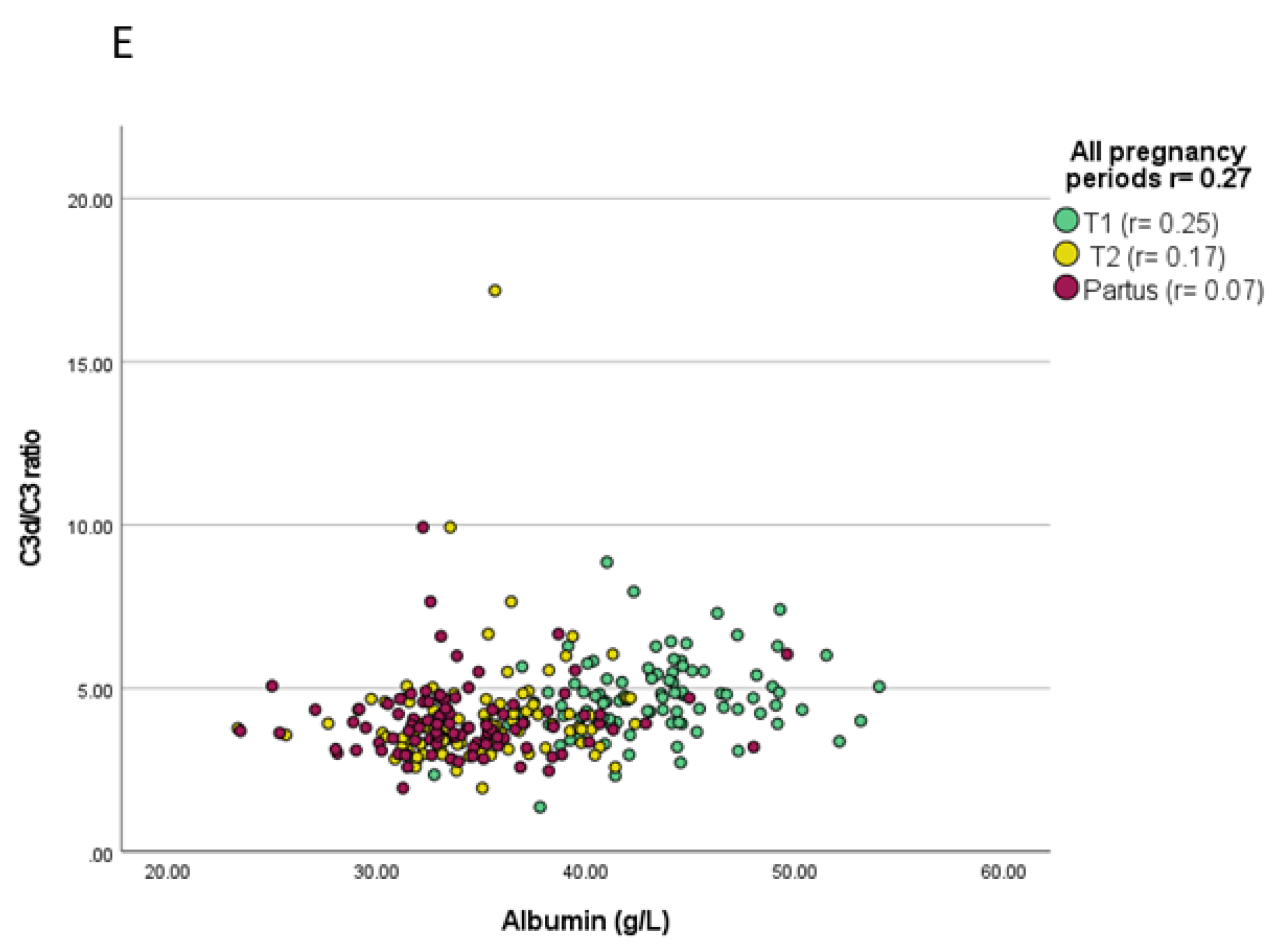
3.2. Associations between Complement Proteins and Evaluated Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rus, H.; Cudrici, C.; Niculescu, F. The role of the complement system in innate immunity. Immunol. Res. 2005, 33, 103–112. [Google Scholar] [CrossRef]
- Triggianese, P.; Perricone, C.; Chimenti, M.S.; De Carolis, C.; Perricone, R. Innate Immune System at the Maternal-Fetal Interface: Mechanisms of Disease and Targets of Therapy in Pregnancy Syndromes. Am. J. Reprod. Immunol. 2016, 76, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Girardi, G. Complement activation, a threat to pregnancy. Semin. Immunopathol. 2018, 40, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Fuzzi, B.; Rizzo, R.; Criscuoli, L.; Noci, I.; Melchiorri, L.; Scarselli, B.; Bencini, E.; Menicucci, A.; Baricordi, O.R. HLA-G expression in early embryos is a fundamental prerequisite for the obtainment of pregnancy. Eur. J. Immunol. 2002, 32, 311–315. [Google Scholar] [CrossRef]
- Rouas-Freiss, N.; Marchal, R.E.; Kirszenbaum, M.; Dausset, J.; Carosella, E.D. The alpha1 domain of HLA-G1 and HLA-G2 inhibits cytotoxicity induced by natural killer cells: Is HLA-G the public ligand for natural killer cell inhibitory receptors? Proc. Natl. Acad. Sci. USA 1997, 94, 5249–5254. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of allogeneic fetal rejection by tryptophan catabolism. Science 1998, 281, 1191–1193. [Google Scholar] [CrossRef]
- Strunk, T.; Currie, A.; Richmond, P.; Simmer, K.; Burgner, D. Innate immunity in human newborn infants: Prematurity means more than immaturity. J. Matern. Fetal Neonatal Med. 2011, 24, 25–31. [Google Scholar] [CrossRef]
- Trowsdale, J.; Betz, A.G. Mother’s little helpers: Mechanisms of maternal-fetal tolerance. Nat. Immunol. 2006, 7, 241–246. [Google Scholar] [CrossRef]
- Bulla, R.; Agostinis, C.; Bossi, F.; Rizzi, L.; Debeus, A.; Tripodo, C.; Radillo, O.; De Seta, F.; Ghebrehiwet, B.; Tedesco, F. Decidual endothelial cells express surface-bound C1q as a molecular bridge between endovascular trophoblast and decidual endothelium. Mol. Immunol. 2008, 45, 2629–2640. [Google Scholar] [CrossRef] [Green Version]
- Nayak, A.; Pednekar, L.; Reid, K.B.; Kishore, U. Complement and non-complement activating functions of C1q: A prototypical innate immune molecule. Innate Immun. 2012, 18, 350–363. [Google Scholar] [CrossRef]
- Agostinis, C.; Tedesco, F.; Bulla, R. Alternative functions of the complement protein C1q at embryo implantation site. J. Reprod. Immunol. 2017, 119, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Buurma, A.; Cohen, D.; Veraar, K.; Schonkeren, D.; Claas, F.H.; Bruijn, J.A.; Bloemenkamp, K.W.; Baelde, H.J. Preeclampsia is characterized by placental complement dysregulation. Hypertension 2012, 60, 1332–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokki, A.I.; Heikkinen-Eloranta, J.; Jarva, H.; Saisto, T.; Lokki, M.-L.; Laivuori, H.; Meri, S. Complement activation and regulation in preeclamptic placenta. Front. Immunol. 2014, 5, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banadakoppa, M.; Chauhan, M.S.; Havemann, D.; Balakrishnan, M.; Dominic, J.S.; Yallampalli, C. Spontaneous abortion is associated with elevated systemic C5a and reduced mRNA of complement inhibitory proteins in placenta. Clin. Exp. Immunol. 2014, 177, 743–749. [Google Scholar] [CrossRef]
- Lynch, A.M.; Gibbs, R.S.; Murphy, J.R.; Byers, T.; Neville, M.C.; Giclas, P.C.; Salmon, J.E.; Van Hecke, T.M.; Holers, V.M. Complement activation fragment Bb in early pregnancy and spontaneous preterm birth. Am. J. Obstet. Gynecol. 2008, 199, 345.e1–345.e8. [Google Scholar] [CrossRef]
- Regal, J.F.; Burwick, R.; Fleming, S.D. The Complement System and Preeclampsia. Curr. Hypertens. Rep. 2017, 19, 87. [Google Scholar] [CrossRef]
- Velickovic, I.; Dalloul, M.; Wong, K.A.; Bakare, O.; Schweis, F.; Garala, M.; Alam, A.; Medranda, G.; Lekovic, J.; Shuaib, W.; et al. Complement factor B activation in patients with preeclampsia. J. Reprod. Immunol. 2015, 109, 94–100. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Xu, B.; Song, D.; Yu, F.; Chen, Q.; Zhao, M. Expression of the complement system’s activation factors in plasma of patients with early/late-onset severe pre-eclampsia. Am. J. Reprod. Immunol. 2016, 76, 205–211. [Google Scholar] [CrossRef]
- Sonntag, J.; Brandenburg, U.; Polzehl, D.; Strauss, E.; Vogel, M.; Dudenhausen, J.; Obladen, M. Complement system in healthy term newborns: Reference values in umbilical cord blood. Pediatr. Dev. Pathol. 1998, 1, 131–135. [Google Scholar] [CrossRef]
- Wolach, B.; Dolfin, T.; Regev, R.; Gilboa, S.; Schlesinger, M. The development of the complement system after 28 weeks’ gestation. Acta Paediatr. 1997, 86, 523–527. [Google Scholar] [CrossRef]
- Zilow, G.; Zilow, E.P.; Burger, R.; Linderkamp, O. Complement activation in newborn infants with early onset infection. Pediatr. Res. 1993, 34, 199–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grumach, A.S.; Ceccon, M.E.; Rutz, R.; Fertig, A.; Kirschfink, M. Complement profile in neonates of different gestational ages. Scand. J. Immunol. 2014, 79, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, M.; Dobson, N.C.; Bradwell, A.R. Synthesis of two components of human complement, beta 1H and C3bINA, during fetal life. Acta Paediatr. Scand. 1981, 70, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Tandon, R.; Bhatia, B.D.; Narang, P.; Tyagi, N.K. Maternal and cord serum C3 level. Indian Pediatr. 1984, 21, 407–413. [Google Scholar] [PubMed]
- Davis, C.A.; Vallota, E.H.; Forristal, J. Serum complement levels in infancy: Age related changes. Pediatr. Res. 1979, 13, 1043–1046. [Google Scholar] [CrossRef] [Green Version]
- Saleh, M.; Sjöwall, C.; Strevens, H.; Jönsen, A.; Bengtsson, A.A.; Compagno, M. Adverse Pregnancy Outcomes after Multi-Professional Follow-Up of Women with Systemic Lupus Erythematosus: An Observational Study from a Single Centre in Sweden. J. Clin. Med. 2020, 9, 2598. [Google Scholar] [CrossRef]
- Johnson, U.; Truedsson, L.; Gustavii, B. Complement components in 100 newborns and their mothers determined by electroimmunoassay. Acta Pathol. Microbiol. Immunol. Scand. C 1983, 91, 147–150. [Google Scholar]
- Wirestam, L.; Pihl, S.; Saleh, M.; Wetterö, J.; Sjöwall, C. Plasma C-Reactive Protein and Pentraxin-3 Reference Intervals During Normal Pregnancy. Front. Immunol. 2021, 12, 722118. [Google Scholar] [CrossRef]
- Grubb, A.; Horio, M.; Hansson, L.-O.; Björk, J.; Nyman, U.; Flodin, M.; Larsson, A.; Bökenkamp, A.; Yasuda, Y.; Blufpand, H.; et al. Generation of a new cystatin C-based estimating equation for glomerular filtration rate by use of 7 assays standardized to the international calibrator. Clin. Chem. 2014, 60, 974–986. [Google Scholar] [CrossRef]
- Sandholm, K.; Persson, B.; Skattum, L.; Eggertsen, G.; Nyman, D.; Gunnarsson, I.; Svenungsson, E.; Nilsson, B.; Ekdahl, K.N. Evaluation of a Novel Immunoassay for Quantification of C1q for Clinical Diagnostic Use. Front. Immunol. 2019, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Ekdahl, K.N.; Norberg, D.; Bengtsson, A.A.; Sturfelt, G.; Nilsson, U.R.; Nilsson, B. Use of serum or buffer-changed EDTA-plasma in a rapid, inexpensive, and easy-to-perform hemolytic complement assay for differential diagnosis of systemic lupus erythematosus and monitoring of patients with the disease. Clin. Vaccine Immunol. 2007, 14, 549–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekdahl, K.N.; Persson, B.; Mohlin, C.; Sandholm, K.; Skattum, L.; Nilsson, B. Interpretation of Serological Complement Biomarkers in Disease. Front. Immunol. 2018, 9, 2237. [Google Scholar] [CrossRef] [PubMed]
- Björk, M.; Dahlström, Ö.; Wetterö, J.; Sjöwall, C. Quality of life and acquired organ damage are intimately related to activity limitations in patients with systemic lupus erythematosus. BMC Musculoskelet. Disord. 2015, 16, 188. [Google Scholar] [CrossRef] [Green Version]
- Frodlund, M.; Vikerfors, A.; Grosso, G.; Skogh, T.; Wetterö, J.; Elvin, K.; Gunnarsson, I.; Kastbom, A.; Dahlström, Ö.; Rönnelid, J.; et al. Immunoglobulin A anti-phospholipid antibodies in Swedish cases of systemic lupus erythematosus: Associations with disease phenotypes, vascular events and damage accrual. Clin. Exp. Immunol. 2018, 194, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Xu, B.; Song, D.; Wang, Y.; Yu, F.; Chen, Q.; Zhao, M. Normal range of complement components during pregnancy: A prospective study. Am. J. Reprod. Immunol. 2020, 83, e13202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, O. Innate immunity of the newborn: Basic mechanisms and clinical correlates. Nat. Rev. Immunol. 2007, 7, 379–390. [Google Scholar] [CrossRef] [PubMed]
- West, L.J. Defining critical windows in the development of the human immune system. Hum. Exp. Toxicol. 2002, 21, 499–505. [Google Scholar] [CrossRef]
- Don, B.R.; Kaysen, G. Serum albumin: Relationship to inflammation and nutrition. Semin. Dial. 2004, 17, 432–437. [Google Scholar] [CrossRef]
- Fink, N.R.; Chawes, B.; Bønnelykke, K.; Thorsen, J.; Stokholm, J.; Rasmussen, M.A.; Brix, S.; Bisgaard, H. Levels of Systemic Low-grade Inflammation in Pregnant Mothers and Their Offspring are Correlated. Sci. Rep. 2019, 9, 3043. [Google Scholar] [CrossRef]
- Watts, D.H.; Krohn, M.A.; Wener, M.H.; Eschenbach, D.A. C-reactive protein in normal pregnancy. Obstet. Gynecol. 1991, 77, 176–180. [Google Scholar] [CrossRef]
- Belo, L.; Santos-Silva, A.; Rocha, S.; Caslake, M.; Cooney, J.; Pereira-Leite, L.; Quintanilha, A.; Rebelo, I. Fluctuations in C-reactive protein concentration and neutrophil activation during normal human pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 123, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Sjöwall, C.; Wetterö, J.; Bengtsson, T.; Askendal, A.; Almroth, G.; Skogh, T.; Tengvall, P. Solid-phase classical complement activation by C-reactive protein (CRP) is inhibited by fluid-phase CRP-C1q interaction. Biochem. Biophys. Res. Commun. 2007, 352, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babay, Z.; Al-Wakeel, J.; Addar, M.; Mittwalli, A.; Tarif, N.; Hammad, D.; Ali, N.; Al-Askar, A.; Choudhary, A.R. Serum cystatin C in pregnant women: Reference values, reliable and superior diagnostic accuracy. Clin. Exp. Obstet. Gynecol. 2005, 32, 175–179. [Google Scholar] [PubMed]
- Strevens, H.; Wide-Swensson, D.; Torffvit, O.; Grubb, A. Serum cystatin C for assessment of glomerular filtration rate in pregnant and non-pregnant women. Indications of altered filtration process in pregnancy. Scand. J. Clin. Lab. Investig. 2002, 62, 141–147. [Google Scholar] [CrossRef]
- Strevens, H.; Wide-Swensson, D.; Hansen, A.; Horn, T.; Ingemarsson, I.; Larsen, S.; Willner, J.; Olsen, S. Glomerular endotheliosis in normal pregnancy and pre-eclampsia. BJOG 2003, 110, 831–836. [Google Scholar] [CrossRef]
- Strevens, H.; Wide-Swensson, D.; Grubb, A.; Hansen, A.; Horn, T.; Ingemarsson, I.; Larsen, S.; Nyengaard, J.R.; Torffvit, O.; Willner, J.; et al. Serum cystatin C reflects glomerular endotheliosis in normal, hypertensive and pre-eclamptic pregnancies. BJOG 2003, 110, 825–830. [Google Scholar] [CrossRef]
- Pomeroy, C.; Mitchell, J.; Eckert, E.; Raymond, N.; Crosby, R.; Dalmasso, A.P. Effect of body weight and caloric restriction on serum complement proteins, including Factor D/adipsin: Studies in anorexia nervosa and obesity. Clin. Exp. Immunol. 1997, 108, 507–515. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean age, years (SD) | 30.0 (3.8) |
| Caucasian ethnicity, n (%) | 92 (92) |
| Mean body mass index at inclusion, kg/m2 (SD) | 22.0 (1.8) |
| Mean gestation duration in weeks + days (range) | 40 + 1 (37 + 4 − 41 + 4) |
| Multiparous, n (%) | 39 (39) |
| Births in January–March, n (%) | 25 (25) |
| Births in April–June, n (%) | 23 (23) |
| Births in July–September, n (%) | 27 (27) |
| Births in October–December, n (%) | 25 (25) |
| Mean birth weight, g (SD) | 3572.2 (399.5) |
| Female newborns, n (%) | 50 (50) |
| Routine Laboratory Analyses (mean (SD)) | |
| T1: P-albumin (g/L) | 42.9 (4.3) |
| T2: P-albumin (g/L) | 34.7 (3.6) |
| Partus: P-albumin (g/L) | 34.1 (4.5) |
| T1: P-cystatin C (mg/L) | 0.74 (0.10) |
| T2: P-cystatin C (mg/L) | 0.84 (0.11) |
| Partus: P-cystatin C (mg/L) | 1.3 (0.28) |
| T1: eGFR (mL/min/1.73 m2) | 116.8 (19.2) |
| T2: eGFR (mL/min/1.73 m2) | 101.3 (17.3) |
| Partus: eGFR (mL/min/1.73 m2) | 60.1 (13.8) |
| Complement | Sampling Occassion | Mean | SD | Min | Max | Percentiles | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 3rd | 5th | 95th | 97th | 99th | ||||||
| C1q (mg/L) Ref. 70–300 | T1 | 84.6 | 22.3 | 47.8 | 205.8 | 47.8 | 50.3 | 52.1 | 112.2 | 122.8 | 205.1 |
| T2 | 69.8 | 15.5 | 34.1 | 113.9 | 34.1 | 39.8 | 45.8 | 95.7 | 103.9 | 113.9 | |
| Partus | 81.5 | 19.7 | 38.0 | 134.9 | 38.0 | 44.5 | 46.9 | 121.8 | 123.9 | 134.8 | |
| UCB | 58.2 | 16.4 | 19.9 | 112.4 | 19.9 | 34.2 | 36.7 | 85.4 | 102.3 | 112.4 | |
| C3 (g/L) Ref. 0.67–1.29 | T1 | 0.95 | 0.17 | 0.43 | 1.3 | 0.43 | 0.65 | 0.67 | 1.2 | 1.2 | 1.3 |
| T2 | 1.1 | 0.16 | 0.47 | 1.6 | 0.47 | 0.81 | 0.82 | 1.3 | 1.4 | 1.6 | |
| Partus | 1.2 | 0.21 | 0.57 | 1.9 | 0.57 | 0.69 | 0.79 | 1.5 | 1.6 | 1.9 | |
| UCB | 0.74 | 0.15 | 0.49 | 1.2 | 0.49 | 0.52 | 0.54 | 1.0 | 1.1 | 1.2 | |
| C4 (g/L) Ref. 0.13–0.32 | T1 | 0.19 | 0.05 | 0.08 | 0.4 | 0.08 | 0.10 | 0.11 | 0.29 | 0.30 | 0.36 |
| T2 | 0.19 | 0.05 | 0.09 | 0.4 | 0.09 | 0.11 | 0.13 | 0.30 | 0.33 | 0.35 | |
| Partus | 0.28 | 0.09 | 0.06 | 0.6 | 0.06 | 0.11 | 0.13 | 0.41 | 0.49 | 0.55 | |
| UCB | 0.13 | 0.04 | 0.05 | 0.3 | 0.05 | 0.08 | 0.09 | 0.20 | 0.21 | 0.31 | |
| C3d (g/L) Ref. < 5.3 | T1 | 4.4 | 0.98 | 1.6 | 6.5 | 1.6 | 2.3 | 2.6 | 6.0 | 6.1 | 6.5 |
| T2 | 4.4 | 1.0 | 2.5 | 7.7 | 2.5 | 2.8 | 2.9 | 6.1 | 6.4 | 7.7 | |
| Partus | 4.9 | 1.9 | 2.2 | 20.1 | 2.2 | 2.8 | 2.9 | 7.6 | 8.1 | 20.1 | |
| UCB | 4.4 | 2.6 | 0.01 | 24.0 | 0.02 | 1.2 | 1.7 | 8.2 | 8.4 | 23.9 | |
| C3d/C3 ratio Ref. < 5.3 | T1 | 4.7 | 1.2 | 1.4 | 8.9 | 1.4 | 2.4 | 3.0 | 6.7 | 7.4 | 8.9 |
| T2 | 4.3 | 1.2 | 2.4 | 10.4 | 2.4 | 2.8 | 2.9 | 6.2 | 6.7 | 10.4 | |
| Partus | 4.1 | 1.7 | 1.9 | 17.2 | 1.9 | 2.6 | 2.7 | 6.6 | 7.8 | 17.2 | |
| UCB | 6.0 | 3.5 | 0.01 | 32.1 | 0.03 | 1.8 | 2.6 | 9.9 | 14.0 | 32.0 | |
| Comp-Lement | Sampling Occasion | Body Mass Index (Mothers’) | P-Cystatin C | Mothers’ Age | Newborns’ Weight | Parity | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| T1 | T2 | Partus | T1 | T2 | Partus | |||||
| C1q (mg/L) | T1 | n.s. | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | p = 0.02 |
| T2 | n.t. | n.s. | n.t. | n.t. | n.s. | n.t. | n.s. | n.s. | n.s. | |
| Partus | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. | n.s. | |
| UCB | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.s. | n.s. | |
| C3 (g/L) | T1 | n.s. | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. |
| T2 | n.t. | r = 0.21; p = 0.04 | n.t. | n.t. | n.s. | n.t. | n.s. | n.s. | n.s. | |
| Partus | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. | n.s. | |
| UCB | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | r = 0.22; p = 0.03 | n.s. | |
| C4 (g/L) | T1 | n.s. | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. |
| T2 | n.t. | r = 0.23; p = 0.03 | n.t. | n.t. | n.s. | n.t. | r = −0.24; p = 0.02 | n.s. | n.s. | |
| Partus | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. | n.s. | |
| UCB | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.s. | n.s. | |
| C3d (g/L) | T1 | r = 0.24; p = 0.02 | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. |
| T2 | n.t. | n.s. | n.t. | n.t. | n.s. | n.t. | n.s. | r = 0.20; p = 0.05 | n.s. | |
| Partus | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. | n.s. | |
| UCB | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.s. | n.s. | |
| C3d/C3 ratio | T1 | n.s. | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. |
| T2 | n.t. | n.s. | n.t. | n.t. | n.s. | n.t. | n.s. | n.s. | n.s. | |
| Partus | n.t. | n.t. | n.s. | n.t. | n.t. | n.s. | n.s. | n.s. | n.s. | |
| UCB | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.s. | n.s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, M.; Compagno, M.; Pihl, S.; Strevens, H.; Persson, B.; Wetterö, J.; Nilsson, B.; Sjöwall, C. Variation of Complement Protein Levels in Maternal Plasma and Umbilical Cord Blood during Normal Pregnancy: An Observational Study. J. Clin. Med. 2022, 11, 3611. https://doi.org/10.3390/jcm11133611
Saleh M, Compagno M, Pihl S, Strevens H, Persson B, Wetterö J, Nilsson B, Sjöwall C. Variation of Complement Protein Levels in Maternal Plasma and Umbilical Cord Blood during Normal Pregnancy: An Observational Study. Journal of Clinical Medicine. 2022; 11(13):3611. https://doi.org/10.3390/jcm11133611
Chicago/Turabian StyleSaleh, Muna, Michele Compagno, Sofia Pihl, Helena Strevens, Barbro Persson, Jonas Wetterö, Bo Nilsson, and Christopher Sjöwall. 2022. "Variation of Complement Protein Levels in Maternal Plasma and Umbilical Cord Blood during Normal Pregnancy: An Observational Study" Journal of Clinical Medicine 11, no. 13: 3611. https://doi.org/10.3390/jcm11133611