
Haptoglobin and Its Related Protein, Zonulin—What Is Their Role in Spondyloarthropathy?

 ,
, 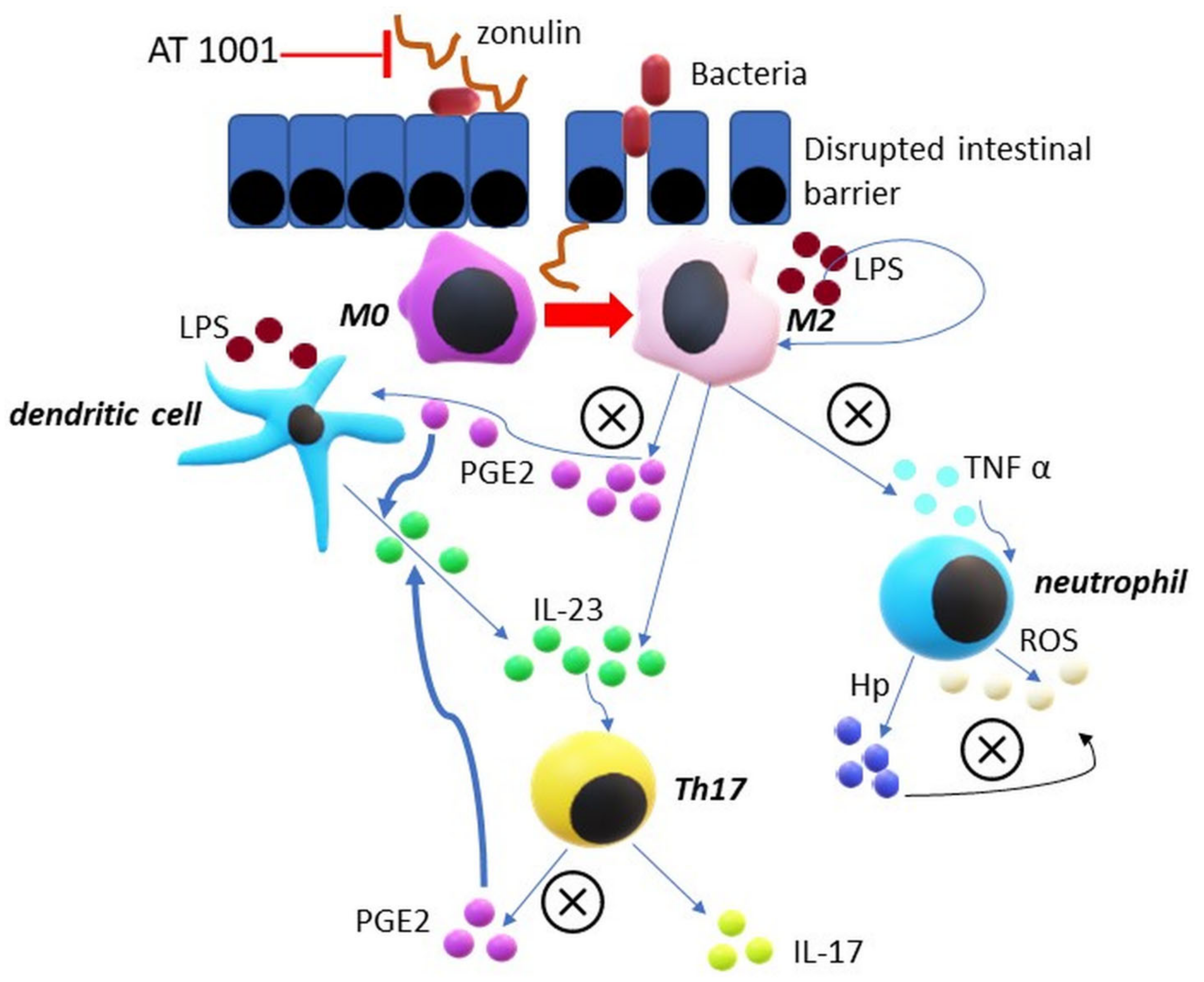{kind=link}
Abstract
:1. Introduction
2. Distribution of Haptoglobin Phenotypes in Spondyloarthropathies
3. The Role of Zonulin and Haptoglobin in Chronic Gut Inflammation
4. The Role of Haptoglobin in Inflammatory Pathways
5. Haptoglobin and Oxidative Stress
6. Conclusions and Research Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bakland, G.; Nossent, H.C. Epidemiology of Spondyloarthritis: A Review. Curr. Rheumatol. Rep. 2013, 15, 1–7. [Google Scholar] [CrossRef]
- Ambarus, C.; Yeremenko, N.; Tak, P.P.; Baeten, D. Pathogenesis of spondyloarthritis. Curr. Opin. Rheumatol. 2012, 24, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Solmaz, D.; Kozaci, D.; Sari, I.; Taylan, A.; Onen, F.; Akkoc, N.; Akar, S. Oxidative stress and related factors in patients with ankylosing spondylitis. Eur. J. Rheum. 2016, 3, 20–24. [Google Scholar] [CrossRef]
- Langlois, M.R.; Delanghe, J.R. Biological and clinical significance of haptoglobin polymorphism in humans. Clin. Chem. 1996, 42, 1589–1600. [Google Scholar] [CrossRef] [Green Version]
- Smeets, M.B.; Fontijn, J.; Kavelaars, A.; Pasterkamp, G.; De Kleijn, D.P. The acute phase protein haptoglobin is locally expressed in arthritic and oncological tissues. Int. J. Exp. Pathol. 2003, 84, 69–74. [Google Scholar] [CrossRef]
- De Kleijn, D.P.V.; Smeets, M.B.; Kemmeren, P.P.C.W.; Lim, S.K.; Van Middelaar, B.J.; Velema, E.; Schoneveld, A.; Pasterkamp, G.; Borst, C. Acute-phase protein haptoglobin is a cell migration factor involved in arterial restructuring. FASEB J. 2002, 16, 1123–1125. [Google Scholar] [CrossRef]
- Levy, A.P.; Asleh, R.; Blum, S.; Levy, N.S.; Miller-Lotan, R.; Kalet-Litman, S.; Anbinder, Y.; Lache, O.; Nakhoul, F.M.; Asaf, R.; et al. Haptoglobin: Basic and clinical aspects. Antioxid. Redox Signal. 2010, 12, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Delanghe, J.R.; Langlois, M.R. Haptoglobin polymorphism and body iron stores. Clin. Chem. Lab. Med. 2002, 40, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz-Sosnowska, A.; Mikołajew, M.; Garwolińska, H.; Sadowska-Wróblewska, M. Przydatność określenia poziomu haptoglobiny jako wskaźnika aktywności procesu zapalnego w surowicy chorych na zesztywniające zapalenie stawów kręgosłupa (Usefulness of serum haptoglobin level determinations as an indicator of the activity of the inflammatory process in patients with ankylosing spondylitis). Reumatologia 1978, 16, 363–368. [Google Scholar] [PubMed]
- Reynolds, T.L.; Khan, M.A.; Van Der Linden, S.; Cleveland, R.P. Differences in HLA-B27 positive and negative patients with ankylosing spondylitis: Study of clinical disease activity and concentrations of serum IgA, C reactive protein, and haptoglobin. Ann. Rheum. Dis. 1991, 50, 154–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inman, R.D.; Baraliakos, X.; Hermann, K.-G.A.; Braun, J.; Deodhar, A.; Van Der Heijde, D.; Xu, S.; Hsu, B. Serum biomarkers and changes in clinical/MRI evidence of golimumab-treated patients with ankylosing spondylitis: Results of the randomized, placebo-controlled GO-RAISE study. Arthritis Res. 2016, 18, 304. [Google Scholar] [CrossRef] [Green Version]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Alessandro, R.; Luchetti, M.M.; Milling, S.; Saieva, L.; Cypers, H.; Stampone, T.; Di Benedetto, P.; et al. Dysbiosis and zonulin upregulation alter gut epithelial and vascular barriers in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2017, 76, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Maeda, N.; Yang, F.; Barnett, D.R.; Bowman, B.H.; Smithies, O. Duplication within the haptoglobin Hp2 gene. Nat. Cell Biol. 1984, 309, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Quaye, I.K. Haptoglobin, inflammation and disease. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 735–742. [Google Scholar] [CrossRef]
- Boettger, L.M.; Salem, R.M.; Handsaker, R.E.; Peloso, G.M.; Kathiresan, S.; Hirschhorn, J.N.; McCarroll, S.A. Recurring exon deletions in the HP (haptoglobin) gene contribute to lower blood cholesterol levels. Nat. Gen. 2016, 48, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.-L.; Lee, K.-Y.; Cheng, T.-M.; Chuang, H.-C.; Wu, S.-M.; Feng, P.-H.; Liu, W.-T.; Chen, K.-Y.; Ho, S.-C. Relationships of haptoglobin phenotypes with systemic inflammation and the severity of chronic obstructive pulmonary disease. Sci. Rep. 2019, 9, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, A.P.; Hochberg, I.; Jablonski, K.; Resnick, H.E.; Lee, E.T.; Best, L.; Howard, B.V. Haptoglobin phenotype is an independent risk factor for cardiovascular disease in individuals with diabetes. J. Am. Coll. Cardiol. 2002, 40, 1984–1990. [Google Scholar] [CrossRef] [Green Version]
- Sertorio, J.T.; Lacchini, R.; Amaral, L.M.; Palei, A.C.T.; Cavalli, R.C.; Sandrim, V.C.; Duarte, G.; Tanus-Santos, J.E. Haptoglobin polymorphism affects nitric oxide bioavailability in preeclampsia. J. Hum. Hypertens. 2012, 27, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Márquez, L.; Shen, C.; Cleynen, I.; De Hertogh, G.; Van Steen, K.; Machiels, K.; Perrier, C.; Ballet, V.; Organe, S.; Ferrante, M.; et al. Effects of haptoglobin polymorphisms and deficiency on susceptibility to inflammatory bowel disease and on severity of murine colitis. Gut 2011, 61, 528–534. [Google Scholar] [CrossRef]
- Cohen, A.S.; Chasen, W. Serum haptoglobin types in rheumatoid spondylitis. Exp. Biol. Med. 1963, 114, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Sitton, N.G.; Dixon, J.S. Haptoglobin phenotypes. Ann. Rheum. Dis. 1983, 42, 356. [Google Scholar] [CrossRef] [Green Version]
- Baeten, D.; Møller, H.J.; Delanghe, J.; Veys, E.M.; Moestrup, S.K.; De Keyser, F. Association of CD163+ macrophages and local production of soluble CD163 with decreased lymphocyte activation in spondylarthropathy synovitis. Arthritis Rheum. 2004, 50, 1611–1623. [Google Scholar] [CrossRef]
- Soliev, T.S.; Arifzhanov, K.R.; Nabieva, D.A.; Mirakhmedova, K. Fenotipy gaptoglobina pri spondiloartritakh i u zdorovykh liudeĭ (Haptoglobin phenotype in patients with spondyloarthritis and healthy individuals). Klinicheskaia Labora-Tornaia Diagnostika 2002, 4, 41–42. [Google Scholar]
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E. Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease. Lancet 2000, 355, 1518–1519. [Google Scholar] [CrossRef]
- Tripathi, A.; Lammers, K.M.; Goldblum, S.; Shea-Donohue, T.; Netzel-Arnett, S.; Buzza, M.S.; Antalis, T.M.; Vogel, S.N.; Zhao, A.; Yang, S.; et al. Identification of human zonulin, a physiological modulator of tight junctions, as prehaptoglobin-2. Proc. Natl. Acad. Sci. USA 2009, 106, 16799–16804. [Google Scholar] [CrossRef] [Green Version]
- Fasano, A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2012, 1258, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhu, P.; Peng, J.; Li, K.; Du, J.; Gu, J.; Ou, Y. Identification of disease-associated proteins by proteomic approach in ankylosing spondylitis. Biochem. Biophys. Res. Commun. 2007, 357, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Mielants, H.; Veys, E.M.; Cuvelier, C.; De Vos, M.; Goemaere, S.; De Clercq, L.; Schatteman, L.; Elewaut, D. The evolution of spondyloarthropathies in relation to gut histology. II. Histological aspects. J. Rheumatol. 1995, 22, 2273–2278. [Google Scholar]
- Van Praet, L.; Bosch, F.E.V.D.; Jacques, P.; Carron, P.; Jans, L.; Colman, R.; Glorieus, E.; Peeters, H.; Mielants, H.; De Vos, M.; et al. Microscopic gut inflammation in axial spondyloarthritis: A multiparametric predictive model. Ann. Rheum. Dis. 2012, 72, 414–417. [Google Scholar] [CrossRef]
- Van Praet, L.; Jans, L.; Carron, P.; Jacques, P.; Glorieus, E.; Colman, R.; Cypers, H.; Mielants, H.; De Vos, M.; Cuvelier, C.; et al. Degree of bone marrow oedema in sacroiliac joints of patients with axial spondyloarthritis is linked to gut inflammation and male sex: Results from the GIANT cohort. Ann. Rheum. Dis. 2013, 73, 1186–1189. [Google Scholar] [CrossRef]
- Rehaume, L.M.; Matigian, N.; Mehdi, A.M.; Lachner, N.; Bowerman, K.L.; Daly, J.; Cao, K.-A.L.; Hugenholtz, P.; Thomas, R. IL-23 favours outgrowth of spondyloarthritis-associated pathobionts and suppresses host support for homeostatic microbiota. Ann. Rheum. Dis. 2019, 78, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Gheita, T.A.; El Gazzar, I.I.; El-Fishawy, H.S.; Aboul-Ezz, M.A.; Kenawy, S.A. Involvement of IL-23 in enteropathic arthritis patients with inflammatory bowel disease: Preliminary results. Clin. Rheumatol. 2014, 33, 713–717. [Google Scholar] [CrossRef]
- Ciccia, F.; Rizzo, A.; Triolo, G. Subclinical gut inflammation in ankylosing spondylitis. Curr. Opin. Rheum. 2016, 28, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, M.J.; Sowders, D.P.; DeLay, M.L.; Mohapatra, R.; Bai, S.; Smith, J.A.; Brandewie, J.R.; Taurog, J.D.; Colbert, R.A. HLA-B27 misfolding in transgenic rats is associated with activation of the unfolded protein response. J. Immunol. 2005, 175, 2438–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Li, J.; He, C.; Li, D.; Tong, W.; Zou, Y.; Xu, W. Role of HLA-B27 in the pathogenesis of ankylosing spondylitis. Mol. Med. Rep. 2017, 15, 1943–1951. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, J.T.; Asquith, M. The microbiome and HLA-B27-associated acute anterior uveitis. Nat. Rev. Rheumatol. 2018, 14, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, I.; Peters, T.J. Influence of anti-rheumatic drugs on gut permeability and on the gut associated lymphoid tissue. Baillière’s Clin. Rheum. 1996, 10, 165–176. [Google Scholar] [CrossRef]
- Nakamura, Y.K.; Janowitz, C.; Metea, C.; Asquith, M.; Karstens, L.; Rosenbaum, J.T.; Lin, P. Short chain fatty acids ameliorate immune-mediated uveitis partially by altering migration of lymphocytes from the intestine. Sci. Rep. 2017, 7, 11745. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Tena, C.; De La Barrera, C.A.; López-Vidal, Y.; Vázquez-Mellado, J.; Richaud-Patin, Y.; Amieva, R.I.; Llorente, L.; Martínez, A.; Zúñiga, J.; Cifuentes-Alvarado, M.; et al. Bacterial DNA in synovial fluid cells of patients with juvenile onset spondyloarthropathies. Rheumatology 2001, 40, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmi, M.; Rajala, P.; Jalkanen, S. Homing of mucosal leukocytes to joints. Distinct endothelial ligands in synovium mediate leukocyte-subtype specific adhesion. J. Clin. Investig. 1997, 99, 2165–2172. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccia, F.; Alessandro, R.; Rizzo, A.; Accardo-Palumbo, A.; Raimondo, S.; Raiata, F.; Guggino, G.; Giardina, A.; De Leo, G.; Sireci, G.; et al. Macrophage phenotype in the subclinical gut inflammation of patients with ankylosing spondylitis. Rheumatology 2013, 53, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeten, D.; Demetter, P.; Cuvelier, C.A.; Kruithof, E.; Van Damme, N.; De Vos, M.; Veys, E.M.; De Keyser, F. Macrophages expressing the scavenger receptor CD163: A link between immune alterations of the gut and synovial inflammation in spondyloarthropathy. J. Pathol. 2002, 196, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Taurog, J.D.; Richardson, J.A.; Croft, J.T.; Simmons, W.A.; Zhou, M.; Fernández-Sueiro, J.L.; Balish, E.; Hammer, R.E. The germfree state prevents development of gut and joint inflammatory disease in HLA-B27 transgenic rats. J. Exp. Med. 1994, 180, 2359–2364. [Google Scholar] [CrossRef]
- Lee, E.-K.; Kang, S.-M.; Paik, D.-J.; Kim, J.M.; Youn, J. Essential roles of Toll-like receptor-4 signaling in arthritis induced by type II collagen antibody and LPS. Int. Immunol. 2005, 17, 325–333. [Google Scholar] [CrossRef]
- Chang, J.; Voorhees, T.; Liu, Y.; Zhao, Y.; Chang, C.-H. Interleukin-23 production in dendritic cells is negatively regulated by protein phosphatase 2A. Proc. Natl. Acad. Sci. USA 2010, 107, 8340–8345. [Google Scholar] [CrossRef] [Green Version]
- Bernink, J.H.; Krabbendam, L.; Germar, K.; de Jong, E.; Gronke, K.; Kofoed-Nielsen, M.; Munneke, J.M.; Hazenberg, M.D.; Villaudy, J.; Buskens, C.J.; et al. Interleukin-12 and -23 control plasticity of CD127+ group 1 and group 3 innate lymphoid cells in the intestinal lamina propria. Immunology 2015, 43, 146–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Saieva, L.; Peralta, S.; Giardina, A.; Cannizzaro, A.; Sireci, G.; De Leo, G.; Alessandro, R.; et al. Type 3 innate lymphoid cells producing IL-17 and IL-22 are expanded in the gut, in the peripheral blood, synovial fluid and bone marrow of patients with ankylosing spondylitis. Ann. Rheum. Dis. 2015, 74, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- El Ghmati, S.M.; Van Hoeyveld, E.M.; Van Strijp, J.G.; Ceuppens, J.L.; Stevens, E.A. Identification of haptoglobin as an alternative ligand for CD11b/CD18. J. Immunol. 1996, 156, 2542–2552. [Google Scholar]
- Zeng, L.; Lindstrom, M.J.; Smith, J.A. Ankylosing spondylitis macrophage production of higher levels of interleukin-23 in response to lipopolysaccharide without induction of a significant unfolded protein response. Arthritis Rheum. 2011, 63, 3807–3817. [Google Scholar] [CrossRef]
- Arredouani, M.S.; Kasran, A.; Vanoirbeek, J.A.; Berger, F.G.; Baumann, H.; Ceuppens, J.L. Haptoglobin dampens endotoxin-induced inflammatory effects both in vitro and in vivo. Immunology 2005, 114, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Raju, S.M.; Kumar, A.P.; Yadav, A.N.; Rajkumar, K.; Mvs, S.; Burgula, S. Haptoglobin improves acute phase response and endotoxin tolerance in response to bacterial LPS. Immunol. Lett. 2019, 207, 17–27. [Google Scholar] [CrossRef]
- Hreggvidsdottir, H.S.; Noordenbos, T.; Baeten, D.L. Inflammatory pathways in spondyloarthritis. Mol. Immunol. 2014, 57, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Boniface, K.; Bak-Jensen, K.S.; Li, Y.; Blumenschein, W.M.; McGeachy, M.J.; McClanahan, T.K.; McKenzie, B.S.; Kastelein, R.A.; Cua, D.J.; Malefyt, R.D.W. Prostaglandin E2 regulates Th17 cell differentiation and function through cyclic AMP and EP2/EP4 receptor signaling. J. Exp. Med. 2009, 206, 535–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheibanie, A.F.; Khayrullina, T.; Safadi, F.F.; Ganea, D. Prostaglandin E2 exacerbates collagen-induced arthritis in mice through the inflammatory interleukin-23/interleukin-17 axis. Arthritis Rheum. 2007, 56, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, K.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular mechanisms underlying prostaglandin E2-exacerbated inflammation and immune diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Klasen, C.; Meyer, A.; Wittekind, P.S.; Waqué, I.; Nabhani, S.; Kofler, D.M. Prostaglandin receptor EP4 expression by Th17 cells is associated with high disease activity in ankylosing spondylitis. Arthritis Res. 2019, 21, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinath, A.; Guggino, G.; Sari, I.; Zeng, F.; Ciccia, F.; Haroon, N. Prostaglandin E2 and Its Receptor Subtype EP4 Are Involved in Ankylosing Spondylitis Disease Progression. Available online: https://acrabstracts.org/abstract/prostaglandin-e2-and-its-receptor-subtype-ep4-are-involved-in-ankylosing-spondylitis-disease-progression/ (accessed on 16 April 2020). [CrossRef]
- Li, M.; Thompson, D.D.; Paralkar, V.M. Prostaglandin E2 receptors in bone formation. Int. Orthop. 2007, 31, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Dougados, M.; Baeten, D. Spondyloarthritis. Lancet 2011, 377, 2127–2137. [Google Scholar] [CrossRef]
- Kroon, F.; Landewé, R.; Dougados, M.; Van Der Heijde, D. Continuous NSAID use reverts the effects of inflammation on radiographic progression in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2012, 71, 1623–1629. [Google Scholar] [CrossRef] [Green Version]
- Shim, B.-S. Increase in serum haptoglobin stimulated by prostaglandins. Nat. Cell Biol. 1976, 259, 326–327. [Google Scholar] [CrossRef]
- Arredouani, M.; Matthijs, P.; Van Hoeyveld, E.; Kasran, A.; Baumann, H.; Ceuppens, J.L.; Stevens, E. Haptoglobin directly affects T cells and suppresses T helper cell type 2 cytokine release. Immunology 2003, 108, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxid. Med. Cell. Longev. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Navid, F.; LiCausi, F.; Nguyen, B.; Cougnoux, A.; Violet, P.; Levine, M.; Colbert, R. Targeting the Oxidative Stress Pathway in Experimental Spondyloarthritis Reduces Pro-Inflammatory Response in Rat Macrophages and Modulates Their Metabolic Requirements (Abstract). Arthritis Rheumatol. 2019, 71. Available online: https://acrabstracts.org/abstract/targeting-the-oxidative-stress-pathway-in-experimental-spondyloarthritis-reduces-pro-inflammatory-response-in-rat-macrophages-and-modulates-their-metabolic-requirements/ (accessed on 14 December 2020).
- Coaccioli, S.; Panaccione, A.; Biondi, R.; Sabatini, C.; Landucci, P.; Del Giorno, R.; Fantera, M.; Mondo, A.M.; Di Cato, L.; Paladini, A.; et al. Evaluation of oxidative stress in rheumatoid and psoriatic arthritis and psoriasis. LA Clin. Ter. 2009, 160, 467–472. [Google Scholar]
- Pishgahi, A.; Abolhasan, R.; Danaii, S.; Amanifar, B.; Soltani-Zangbar, M.S.; Zamani, M.; Kamrani, A.; Ghorbani, F.; Mehdizadeh, A.; Kafil, H.S.; et al. Immunological and oxidative stress biomarkers in Ankylosing Spondylitis patients with or without metabolic syndrome. Cytokine 2020, 128, 155002. [Google Scholar] [CrossRef]
- Wang, L.; Gao, L.; Jin, D.; Wang, P.; Yang, B.; Deng, W.; Xie, Z.; Tang, Y.; Wu, Y.; Shen, H. The relationship of bone mineral density to oxidant/antioxidant status and inflammatory and bone turnover markers in a multicenter cross-sectional study of young men with ankylosing spondylitis. Calcif. Tissue Int. 2015, 97, 12–22. [Google Scholar] [CrossRef]
- Zhang, D.-Y.; Pan, Y.; Zhang, C.; Yan, B.-X.; Yu, S.-S.; Wu, D.-L.; Shi, M.-M.; Shi, K.; Cai, X.-X.; Zhou, S.-S.; et al. Wnt/β-catenin signaling induces the aging of mesenchymal stem cells through promoting the ROS production. Mol. Cell. Biochem. 2012, 374, 13–20. [Google Scholar] [CrossRef]
- Li, J.; Liu, S.; Cui, Y. Oxidative and antioxidative stress linked biomarkers in ankylosing spondylitis: A systematic review and meta-analysis. Oxid. Med. Cell. Longev. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Feijóo, M.; Túnez, I.; Tasset, I.; Montilla, P.; Ruiz, A.; Collantes, E. Infliximab reduces oxidative stress in ankylosing spondylitis. Clin. Exp. Rheum. 2009, 27, 167. [Google Scholar]
- Tseng, C.F.; Lin, C.C.; Huang, H.Y.; Liu, H.C.; Mao, S.J.T. Antioxidant role of human haptoglobin. Proteomics 2004, 4, 2221–2228. [Google Scholar] [CrossRef]
- Oh, S.-K.; Pavlotsky, N.; Tauber, A. specific binding of haptoglobin to human neutrophils and its functional consequences. J. Leukoc. Biol. 1990, 47, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Berkova, N.; Gilbert, C.; Goupil, S.; Yan, J.; Korobko, V.; Naccache, P.H. TNF-induced haptoglobin release from human neutrophils: Pivotal role of the TNF p55 receptor. J. Immunol. 1999, 162, 6226–6232. [Google Scholar]
- Ho, K.-J.; Chen, P.-Q.; Chang, C.-Y.; Lu, F.-J. The oxidative metabolism of circulating phagocytes in ankylosing spondylitis: Determination by whole blood chemiluminescence. Ann. Rheum. Dis. 2000, 59, 338–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaleghi, S.; Ju, J.M.; Lamba, A.; Murray, J.A. The potential utility of tight junction regulation in celiac disease: Focus on larazotide acetate. Ther. Adv. Gastroenterol. 2015, 9, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Leffler, D.A.; Kelly, C.P.; Green, P.H.; Fedorak, R.N.; Dimarino, A.; Perrow, W.; Rasmussen, H.; Wang, C.; Bercik, P.; Bachir, N.M.; et al. Larazotide acetate for persistent symptoms of celiac disease despite a gluten-free diet: A randomized controlled trial. Gastroenterology 2015, 148, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.P.; Green, P.H.R.; Murray, J.A.; Dimarino, A.; Colatrella, A.; Leffler, D.A.; Alexander, T.; Arsenescu, R.; Leon, F.; Jiang, J.G.; et al. Larazotide acetate in patients with coeliac disease undergoing a gluten challenge: A randomised placebo-controlled study. Aliment. Pharmacol. Ther. 2012, 37, 252–262. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmielińska, M.; Olesińska, M.; Romanowska-Próchnicka, K.; Szukiewicz, D. Haptoglobin and Its Related Protein, Zonulin—What Is Their Role in Spondyloarthropathy? J. Clin. Med. 2021, 10, 1131. https://doi.org/10.3390/jcm10051131
Chmielińska M, Olesińska M, Romanowska-Próchnicka K, Szukiewicz D. Haptoglobin and Its Related Protein, Zonulin—What Is Their Role in Spondyloarthropathy? Journal of Clinical Medicine. 2021; 10(5):1131. https://doi.org/10.3390/jcm10051131
Chicago/Turabian StyleChmielińska, Magdalena, Marzena Olesińska, Katarzyna Romanowska-Próchnicka, and Dariusz Szukiewicz. 2021. "Haptoglobin and Its Related Protein, Zonulin—What Is Their Role in Spondyloarthropathy?" Journal of Clinical Medicine 10, no. 5: 1131. https://doi.org/10.3390/jcm10051131