
CD28 Gene Polymorphisms in the Promoter Region Are Associated with Transfusion Reactions: A Functional Study

Abstract
:1. Introduction
2. Experimental Section
2.1. Subjects
2.2. DNA Extraction
2.3. PCR Amplification and Sequencing
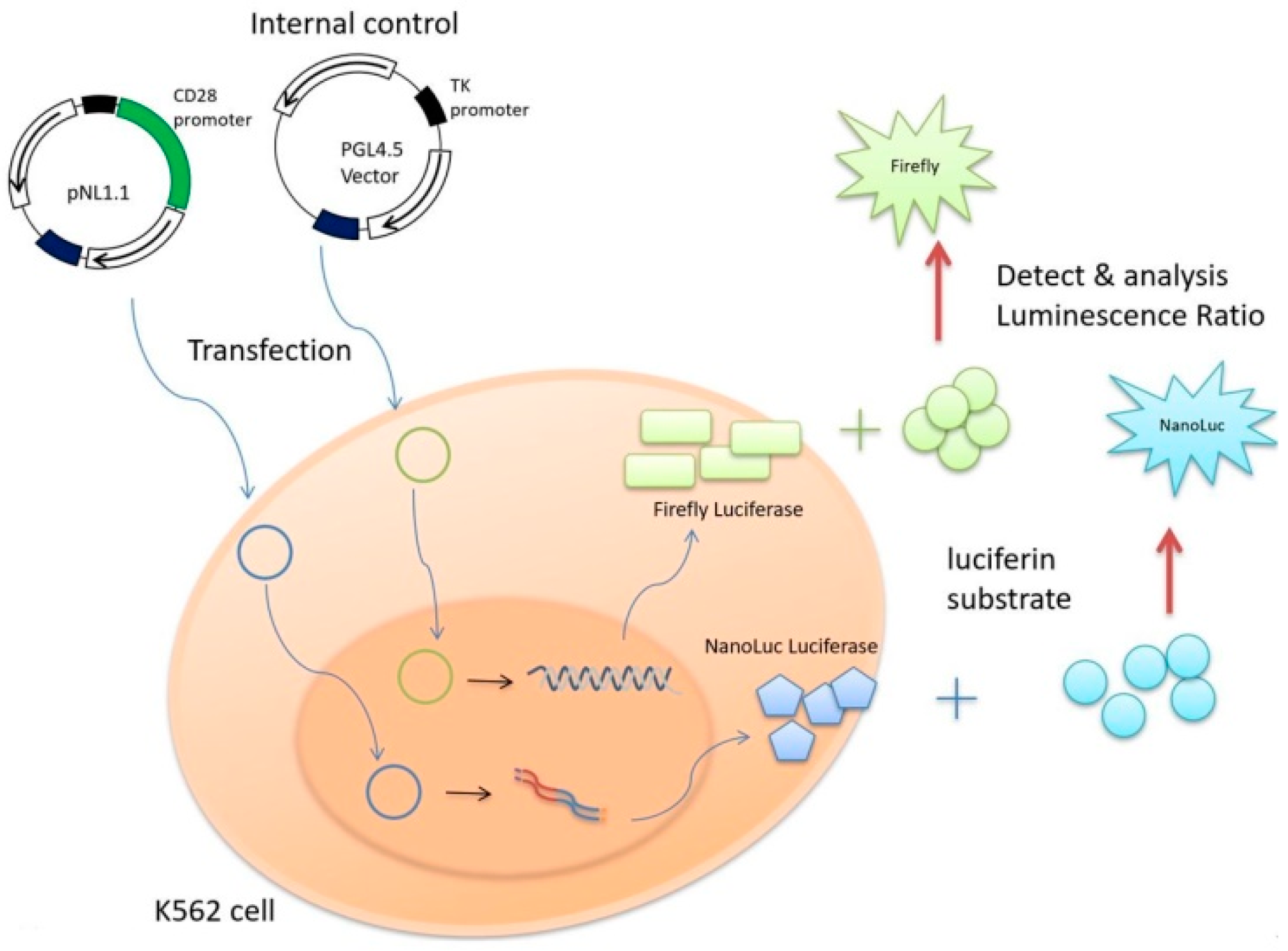
2.4. Promoter-Reporter Constructs
2.5. Cell Culture and Transient Transfections
2.6. Luciferase Reporter Assay
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blood Transfusions for People with Cancer. American Cancer Society. Available online: https://www.cancer.org/treatment/treatments-and-side-effects/treatment-types/blood-transfusion-and-donation/what-are-transfusions.html (accessed on 21 September 2020).
- Dean, L. Blood Groups and Red Cell Antigens [Internet]; Blood Transfusions and the Immune System; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2005; Chapter 3. Available online: https://www.ncbi.nlm.nih.gov/books/NBK2265/ (accessed on 11 September 2020).
- Ortolano, G.A.; Russell, R.L.; Angelbeck, J.A.; Schaer, J.; Wenz, B. Contamination control in nursing with filtration: Part 2: Emerging rationale for bedside (final) filtration of prestorage leukocyte-reduced blood products. J. Infus. Nurs. 2004, 27, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Mempel, W.; Böck, M. Substitution of thrombocyte concentrates in polytransfused patients. Beitr. Infusionsther. 1993, 31, 81–85. [Google Scholar]
- Rajesh, K.; Harsh, S.; Amarjit, K. Effects of Prestorage Leukoreduction on the Rate of Febrile Nonhemolytic Transfusion Reactions to Red Blood Cells in a Tertiary Care Hospital. Ann. Med. Health Sci. Res. 2015, 5, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.P.; Lin, W.T.; Wang, W.T.; Chiueh, T.S. The Influence of CD28 Gene Polymorphism in Transfusion Reaction after Transfusing Leukoreduced Blood Components. J. Clin. Med. 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.H.; Lin, W.T.; Wang, W.T.; Chiueh, T.S.; Chen, D.P. Association of CTLA4 Gene Polymorphism with Transfusion Reaction after Infusion of Leukoreduced Blood Component. J. Clin. Med. 2019, 8, 1961. [Google Scholar] [CrossRef] [Green Version]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Chapter 8: T Cell-Mediated Immunity. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. Available online: https://www.ncbi.nlm.nih.gov/books/NBK10762/ (accessed on 30 June 2020).
- Budd, R.C.; Fortner, K.A. T Lymphocytes. In Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Firestein, G.S., Budd, R.C., Gabriel, S.E., McInnes, I.B., O’Dell, J.R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Chapter 12; pp. 189–206. [Google Scholar]
- Zajac, A.J.; Blattman, J.N.; Murali-Krishna, K.; Sourdive, D.J.; Suresh, M.; Altman, J.D.; Ahmed, R. Viral Immune Evasion Due to Persistence of Activated T Cells without Effector Function. J. Exp. Med. 1998, 188, 2205–2213. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H. Mechanisms of costimulation. Immunol. Rev. 2009, 229, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porciello, N.; Tuosto, L. CD28 costimulatory signals in T lymphocyte activation: Emerging functions beyond a qualitative and quantitative support to TCR signaling. Cytokine Growth Factor Rev. 2016, 28, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylichenko, O.; Bondarenko, V.; Harnett, D.; Schor, I.E.; Males, M.; Viales, R.R.; Furlongl, E.E.M. The degree of enhancer or promoter activity is reflected by the levels and directionality of eRNA transcription. Genes Dev. 2018, 32, 42–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Chung, J.H.; Kang, S.; Kim, S.K.; Cho, B.S.; Kim, S.D.; Hahn, W.H. Association of CTLA4, CD28 and ICOS gene polymorphisms with clinicopathologic characteristics of childhood IgA nephropathy in Korean population. J. Genet. 2011, 90, 151–155. [Google Scholar] [CrossRef]
- Ferjeni, Z.; Bouzid, D.; Fourati, H.; Stayoussef, M.; Abida, O.; Kammoun, T.; Hachicha, M.; Penha-Gonçalves, C.; Masmoudi, H. Association of TCR/CD3, PTPN22, CD28 and ZAP70 gene polymorphisms with type 1 diabetes risk in Tunisian population: Family based association study. Immunol. Lett. 2015, 163, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Sun, J. Interactive Effects of Snps Located Within CD28/B7Pathway and Environment on Susceptibility to Recurrent Spontaneous Abortion. Cell Physiol. Biochem. 2017, 43, 2185–2199. [Google Scholar] [CrossRef] [Green Version]
- Tupikowski, K.; Partyka, A.; Kolodziej, A.; Dembowski, J.; Debinski, P.; Halon, A.; Zdrojowy, R.; Frydecka, I.; Karabon, L. CTLA-4 and CD28 genes’ polymorphisms and renal cell carcinoma susceptibility in the Polish population—A prospective study. Tissue Antigens 2015, 86, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Devin, M. Associations between Genetic Variation in the T-Cell Signaling Pathways and CD4+ T-Cell Count Recovery after ART Initiation in Southern African Cohort. Master’s Thesis, School of Molecular and Cell Biology, University of the Witwatersrand Johannesburg, Johannesburg, South Africa, 2018. Available online: https://hdl.handle.net/10539/27071 (accessed on 21 September 2020).
- Beyersdorf, N.; Kerkau, T.; Hünig, T. CD28 co-stimulation in T-cell homeostasis: A recent perspective. Immunotargets Ther. 2015, 4, 111–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adcock, I.M.; Caramori, G. Chapter 31: Transcription Factors. In Asthma and COPD, 2nd ed.; Barnes, P., Drazen, J., Thomson, N., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 373–380. [Google Scholar]
- Mitsis, T.; Efthimiadou, A.; Bacopoulou, F.; Vlachakis, D.; Chrousos, G.P.; Eliopoulos, E. Transcription factors and evolution: An integral part of gene expression (Review). World Acad. Sci. J. 2020, 2, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Fortelny, N.; Overall, C.M.; Pavlidis, P.; Freue, G.V.C. Can we predict protein from mRNA levels? Nature 2017, 547, E19–E20. [Google Scholar] [CrossRef]
- Science/Biology Library/Gene Regulation/Gene Regulation in Eukaryotes/Regulation after Transcription. Khan Academy. Available online: https://www.khanacademy.org/science/biology/gene-regulation/gene-regulation-in-eukaryotes/a/regulation-after-transcription (accessed on 7 July 2020).
- Chen, Q.Y.; Jackson, N. Human CD1D gene has TATA boxless dual promoters: An SP1-binding element determines the function of the proximal promoter. J. Immunol. 2004, 172, 5512–5521. [Google Scholar] [CrossRef]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Garcon, F.; Patton, D.T.; Emery, J.L.; Hirsch, E.; Rottapel, R.; Sasaki, T.; Okkenhaug, K. CD28 provides T-cell costimulation and enhances PI3K activity at the immune synapse independently of its capacity to interact with the p85/p110 heterodimer. Blood 2008, 111, 1464–1471. [Google Scholar] [CrossRef]
- Schneider, H.; Cai, Y.C.; Prasad, K.V.; Shoelson, S.E.; Rudd, C.E. T cell antigen CD28 binds to the GRB-2/SOS complex, regulators of p21ras. Eur. J. Immunol. 1995, 25, 1044–1050. [Google Scholar] [CrossRef]
- Marengère, L.E.; Okkenhaug, K.; Clavreul, A.; Couez, D.; Gibson, S.; Mills, G.B.; Mak, T.W.; Rottapel, R. The SH3 domain of Itk/Emt binds to proline-rich sequences in the cytoplasmic domain of the T cell costimulatory receptor CD28. J. Immunol. 1997, 159, 3220–3229. [Google Scholar] [PubMed]
- Landolin, J.M.; Johnson, D.S.; Trinklein, N.D.; Aldred, S.F.; Medina, C.; Shulha, H.; Weng, Z.; Myers, R.M. Sequence features that drive human promoter function and tissue specificity. Genome Res. 2010, 20, 890–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, L.B.; van Dijk, D.; Sloot, P.M.; Kaandorp, J.A.; Segal, E. Promoter sequence determines the relationship between expression level and noise. PLoS Biol. 2013, 11, e1001528. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| SNP | Mutagenesis Primer Sequence | SNP Position |
| rs1879877 (G > T) | F: 5′-TGGAATAACCCTCTCTTCAAAGGGCCTGGGA-3′ R: 5′-TCCCAGGCCCTTTGAAGAGAGGGTTATTCCA-3′ | Promoter -1198 |
| rs3181096 (C > T) | F: 5′-CTCCTTTTGTGCCCTATTATTTAACCTTGAGGG-3′ R: 5′-CCCTCAAGGTTAAATAATAGGGCACAAAAGGAG-3′ | Promoter -1106 |
| rs3181097 (G > A) | F: 5′-ACCAAGGGGCTTTTGATTGCCTTACTGTCCCA-3′ R: 5′-TGGGACAGTAAGGCAATCAAAAGCCCCTTGGT-3′ | Promoter -1059 |
| rs3181098 (G > A) | F: 5′-GTAACTCCTTTAAACATTTATGCAGATGTTTCCC-3′ R:5′-GGGAAACATCTGCATAAATGTTTAAAGGAGTTAC-3′ | Promoter -820 |
| No. (%) | |
|---|---|
| Average age of patients | 54.7 |
| Gender (female:male) | 30:10 |
| Blood type | |
| A type | 10 (25) |
| B type | 9 (22.5) |
| AB type | 3 (7.5) |
| O type | 18 (45) |
| Type of blood products | |
| Leukocyte-poor platelet | 19 (47.5) |
| Leukocyte-poor RBC | 21 (52.5) |
| Type of transfusion reaction | |
| Febrile non-hemolytic transfusion reaction (FNHTR) | 29 (72.5) |
| Allergic reaction | 11 (27.5) |
| SNP | Allele | Minor Allele Frequency | HWE p-Value | Odds Ratio (95% CI.) | p-Value a | |
|---|---|---|---|---|---|---|
| Patient | Control | |||||
| rs1879877 | G/T | 0.413 | 0.338 | 0.952 | 1.378 (0.725–2.621) | 0.327 |
| rs3181096 | C/T | 0.263 | 0.213 | 0.983 | 1.319 (0.635–2.741) | 0.457 |
| rs3181097 | G/A | 0.425 | 0.488 | 0.085 | 0.703 (0.377–1.311) | 0.267 |
| rs3181098 | G/A | 0.263 | 0.213 | 0.983 | 1.319 (0.635–2.741) | 0.457 |
| SNP | Genotype | Patient | Control | Model | Odds Ratio (95% CI) | p-Value |
|---|---|---|---|---|---|---|
| n = 40 | n = 40 | |||||
| rs1879877 | GG | 10 | 5 | A | 0.328 | |
| GT | 13 | 17 | D | 1.107 (0.457–2.679) | 0.822 | |
| TT | 17 | 18 | R | 2.333 (0.718–7.587) | 0.152 | |
| rs3181096 | CC | 24 | 25 | A | 0.479 | |
| CT | 11 | 13 | D | 1.111 (0.452–2.733) | 0.818 | |
| TT | 5 | 2 | R | 2.714 (0.494–14.901) | 0.432 | |
| rs3181097 | GG | 17 | 6 | A | 0.003 * | |
| AG | 12 | 27 | D | 0.287 (0.103–0.803) | 0.015 * | |
| AA | 11 | 7 | R | 2.149 (0.707–6.530) | 0.172 | |
| rs3181098 | GG | 24 | 25 | A | 0.479 | |
| AG | 11 | 13 | D | 2.715 (0.494–14.901) | 0.432 | |
| AA | 5 | 2 | R | 1.111 (0.452–2.733) | 0.818 |
| Test 1 | Test 2 | Test 3 | Test 4 | Test 5 | Average | p-Value | |
|---|---|---|---|---|---|---|---|
| Wild Type | 1 | 1 | 1 | 1 | 1 | 1 | ref |
| rs1878877G > T | 0.85 | 0.98 | 0.91 | 1.1 | 0.98 | 0.964 | 0.438 |
| rs3181096C > T | 1.05 | 1.11 | 0.86 | 1.17 | 0.85 | 1.008 | 0.908 |
| rs3181097G > A | 0.8 | 0.86 | 0.68 | 0.81 | 0.74 | 0.778 | 0.002 * |
| rs3181098G > A | 0.75 | 1.18 | 0.7 | 1.08 | 0.75 | 0.892 | 0.336 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Y.-H.; Lin, W.-T.; Wang, W.-T.; Chen, D.-P. CD28 Gene Polymorphisms in the Promoter Region Are Associated with Transfusion Reactions: A Functional Study. J. Clin. Med. 2021, 10, 871. https://doi.org/10.3390/jcm10040871
Wen Y-H, Lin W-T, Wang W-T, Chen D-P. CD28 Gene Polymorphisms in the Promoter Region Are Associated with Transfusion Reactions: A Functional Study. Journal of Clinical Medicine. 2021; 10(4):871. https://doi.org/10.3390/jcm10040871
Chicago/Turabian StyleWen, Ying-Hao, Wei-Tzu Lin, Wei-Ting Wang, and Ding-Ping Chen. 2021. "CD28 Gene Polymorphisms in the Promoter Region Are Associated with Transfusion Reactions: A Functional Study" Journal of Clinical Medicine 10, no. 4: 871. https://doi.org/10.3390/jcm10040871