1. Introduction
The first diagnosis of coronavirus disease 2019 (COVID-19) cases caused by the severe acute respiratory syndrome coronavirus-type 2 (SARS-CoV-2) occurred in Wuhan, China, in December 2019, and its subsequent spread starting in spring 2020 marked the beginning of a challenging global pandemic and public health crisis, which is unprecedented in modern history [
1]. COVID-19 comprises a wide spectrum of clinical manifestations, ranging from moderate courses of infection with very mild symptoms to life-threatening viral pneumonia. The latter is more frequent in patients with preexisting conditions such as compromised immunity, and the elderly [
2]. SARS-CoV-2 infections do not necessarily cause COVID-19 and a relevant fraction (in some studies up to almost half of all infections) occur without symptoms [
3,
4]. Despite the recent development of highly effective vaccines, the understanding of sustained SARS-CoV-2 immunity in the convalescent unvaccinated population remains crucial for future decision making regarding the ongoing pandemic [
5,
6]. It is particularly relevant for countries with lower vaccine coverage. There is concern that waning immunity may eventually fail to prevent all re-infection. Others, and we, have studied the antibody response against the virus and its kinetics in various population groups [
3,
7,
8,
9,
10]. Several of these longitudinal studies report a biphasic decrease in both the levels of SARS-CoV-2 specific antibodies (IgA, IgG and neutralizing antibodies) with a plateau maintained over several months. Immunological memory depends not only on maintaining high antibody titers but also on the generation of specific T cells that immediately respond if the host reencounters the virus [
11]. In several countries, convalescent people are considered to be protected against reinfections and are treated similarly to vaccinated individuals with regard to exemptions from public health restrictions operative for non-protected people. This poses questions such as the following: How long will the protection last, particularly in asymptomatic or mildly ill individuals? Does the severity of symptoms influence cellular and humoral immune responses and their sustainability? Which factors influence the persistence of long-term immune responses? To address these questions, the cellular and humoral immune response was assessed in a total of 200 unvaccinated SARS-CoV-2 convalescents. Patients were recruited from the post-COVID-19 outpatient clinic of the Department of Infectious Diseases at the University Hospital Essen, since May 2020.
2. Materials and Methods
2.1. Study Subjects and Sampling
This cross-sectional, prospective study included SARS-CoV-2 unvaccinated convalescent individuals who presented to the post-COVID-19 outpatient clinic of the Department of Infectious Diseases of the University Hospital Essen in May 2020. To reach as many convalescents as possible, we drew attention to the study by the Health Department of the city of Essen and via the public media. Inclusion criteria were an age at least 18 years and previously confirmed detection of SARS-CoV-2 viral RNA in swab or sputum by reverse transcription polymerase chain reaction (RT-PCR); exclusion criterion was vaccination against SARS-CoV-2. Patients were grouped into categories according to their worst disease manifestation at the time of primary COVID-19, as defined by World Health Organization (WHO) criteria (asymptomatic or mild disease, i.e., WHO score 1–2; hospitalized with moderate disease, i.e., WHO score 3–4, severe disease requiring intensive care treatment, i.e., WHO score 5–7). Peripheral blood samples were collected from all patients for immunological analyses. The median interval between the PCR-confirmed SARS-CoV-2 infection and the presentation to the post-COVID-19 outpatient clinic was 5 months (range 1–11 months). The study was approved by the local ethics committee (approval no. 20-9374-BO) and was performed in accordance with ethics standards noted in the 1964 Declaration of Helsinki and its later amendments or comparable ethics standards. All patients provided informed consent for participation in the study.
2.2. T Cell ELISpot Assays for S- and M-Derived SARS-CoV-2 Peptides
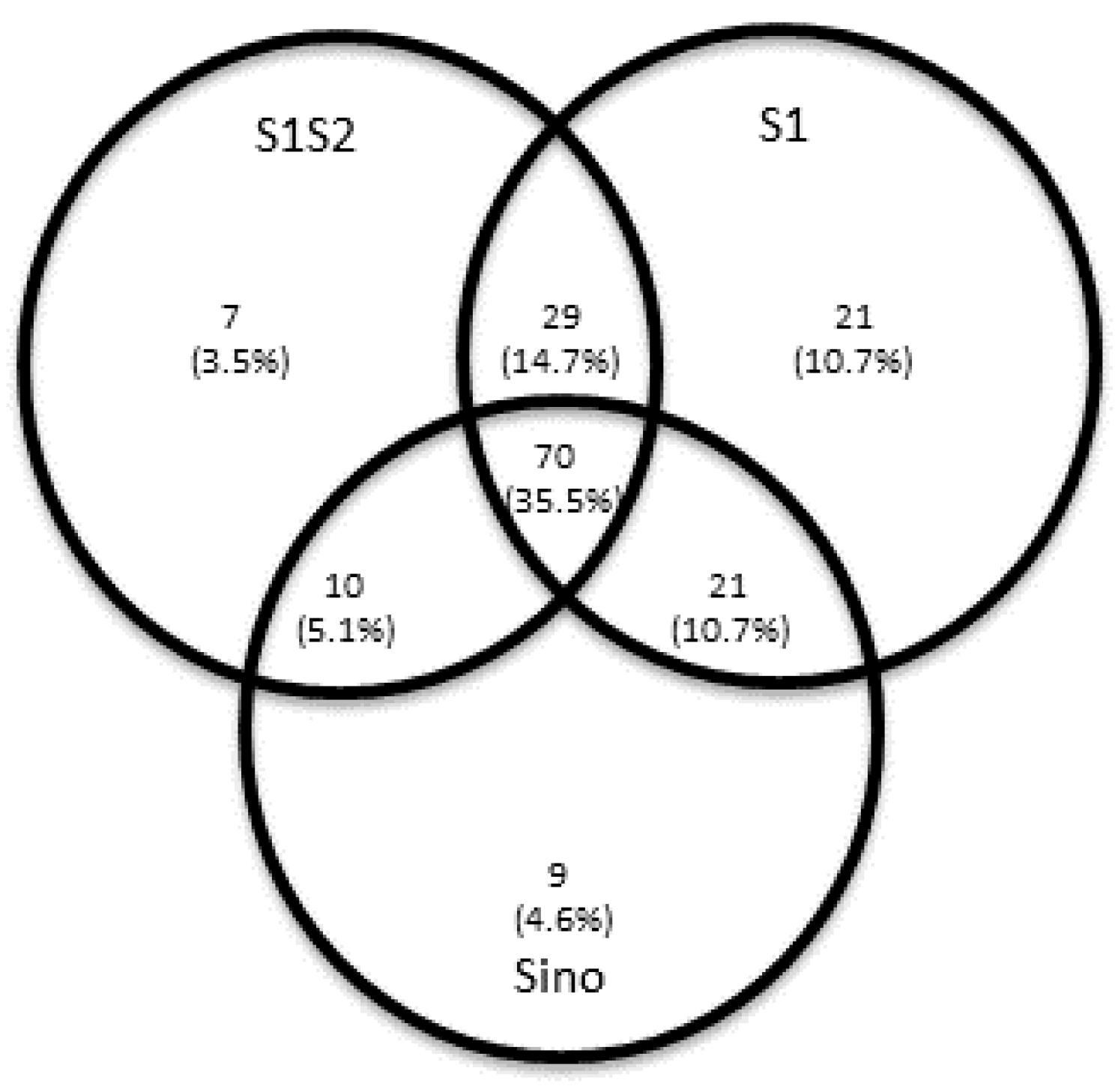
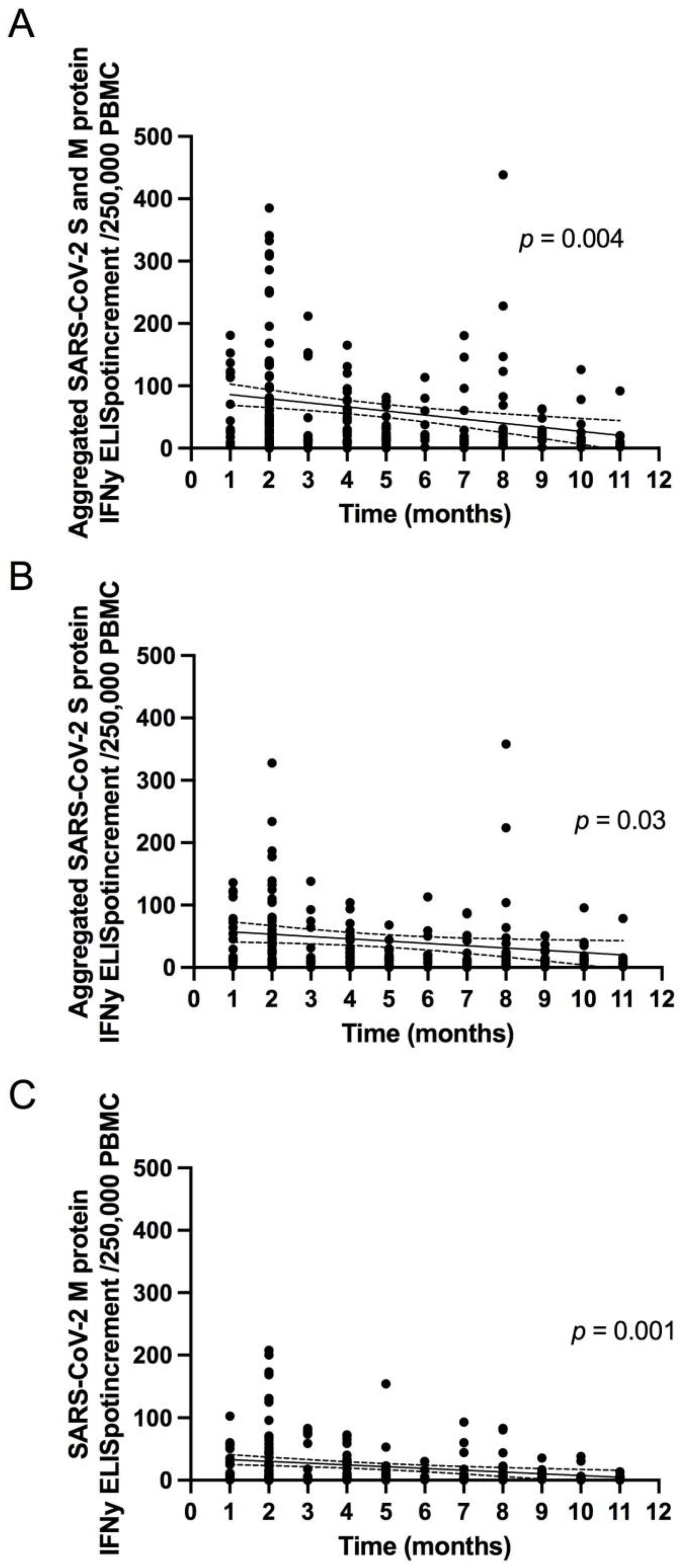
To explore the SARS-CoV-2 specific T cell response, we collected peripheral blood mononuclear cells (PBMCs) for interferon gamma (IFN-γ) enzyme-linked immunospot (ELISpot) assays. For 197 of 200 patients, ELISpot assays were performed applying recently described methodologies [
11,
12]. To cover a broad range of putative antigens for Spike (S) protein and Membrane (M) protein and to increase sensitivity, the following four peptide pools or proteins were used: (I) PepTivator SARS-CoV-2 protein S1/S2 (Miltenyi Biotec), (II) PepTivator SARS-CoV-2 protein S1 protein (Miltenyi Biotec), (III) an S1 protein antigen of SARS-CoV-2 (Sino Biological) or (IV) PepTivator SARS-CoV-2 membrane (M) protein (Miltenyi Biotec). The peptide mix (PepTivator) of the S1/S2 protein covers the immunodominant domains, whereas the peptide mix corresponding to the M protein covers the complete sequence of the glycoprotein. The peptide pools consist mainly of 15-mer sequences with an overlap of 11 amino acids. Per cell culture, 250,000 PBMCs were tested, and IFN-γ production was measured after 19 h, as previously described [
11,
13]. Spot numbers were evaluated by an ELISpot reader (AID Fluorospot; Autoimmun Diagnostika GmbH, Strassberg, Germany). Mean values of duplicate cell cultures were considered. SARS-CoV-2 specific spots were determined as stimulated minus non-stimulated (background) values (spot increment). A positive response was defined as a threefold increase in SARS-CoV-2 specific spots compared with background and at least three spots above background. This cut-off was set on the basis of the negative control values, as previously described. The ELISpot results were evaluated both for all S peptide pool/protein antigens and M peptide pools individually and in aggregated form (S plus M ELISpot results).
2.3. SARS-CoV-2 Specific Antibody Detection
SARS-CoV-2 specific antibodies were detected by two separate methods. Initially, for 84 patients, SARS-CoV-2 immunoglobulin (IgG) against the spike glycoprotein were detected by a Communauté Européenne (CE)-marked anti-SARS-CoV-2 IgG semi-quantitative enzyme-linked immunosorbent assay (ELISA) (Euroimmun, Luebeck, Germany) in accordance with the manufacturer’s instructions. The ELISA plates were coated with recombinant SARS-CoV-2 spike protein (S1 domain). Serum samples were automatically analyzed at a dilution of 1:101 with the Immunomat (Virion\Serion). The results were given as the ratio of patient sample to control sample. An antibody ratio higher than 1.1 was considered positive, a ratio of 0.8 to less than 1.1 was considered borderline, and a ratio lower than 0.8 was considered negative. According to the manufacturer’s instructions, an evaluation of a borderline result is not possible. For statistical analysis, only the clearly positive or negative results of these semi-quantitative analyses were taken into account and dichotomized with respect to the presence of SARS-CoV-2 IgG antibodies.
After a quantitative automatized anti-SARS-CoV-2 IgG chemiluminescent enzyme immunoassay (CLIA) (LIAISON SARS-CoV-2 TrimericS IgG assay, Diasorin, Saluggia, Italy) became available in February 2021, the detection method for SARS-CoV-2 IgG antibodies was changed, and 128 patients were assessed using this method (with 17 patients tested with both antibody detection methods). According to the manufacturer’s recommendations for CLIA, an arbitrary unit per milliliter (AU/mL) ratio of less than 13.0 was considered negative and a ratio of 13.0 or higher was considered positive. A conversion of AU/mL to binding antibody units (BAU/mL), which correlate with the WHO standard, is possible with the following equation: BAU/mL = 2.6*AU/mL. The upper limit of quantification of the CLIA test without dilution is 800.0 AU/mL (2080 BAU/mL).
2.4. Cells and Virus
Vero E6 cells were purchased from ATCC (Manassas, VA, USA; ATCC
® RL-1586™) and maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 1% fetal calf serum (FCS), penicillin (100 IU/mL), and streptomycin (100 μg/mL). SARS-CoV-2 was isolated from a nasopharyngeal swab of a patient suffering from COVID-19 in April 2020 as previously described [
14]. The virus was propagated on Vero E6 cells cultured in DMEM containing 10% (
v/
v) FCS and supplemented with penicillin (100 IU/mL), streptomycin (100 μg/mL), ciprofloxacin (10 μg/mL), and amphotericin B (2.5 μg/mL). Viral titers were determined by endpoint dilution assay and expressed as 50% tissue culture infective dose (TCID50)/mL.
2.5. Virus Neutralization Assay
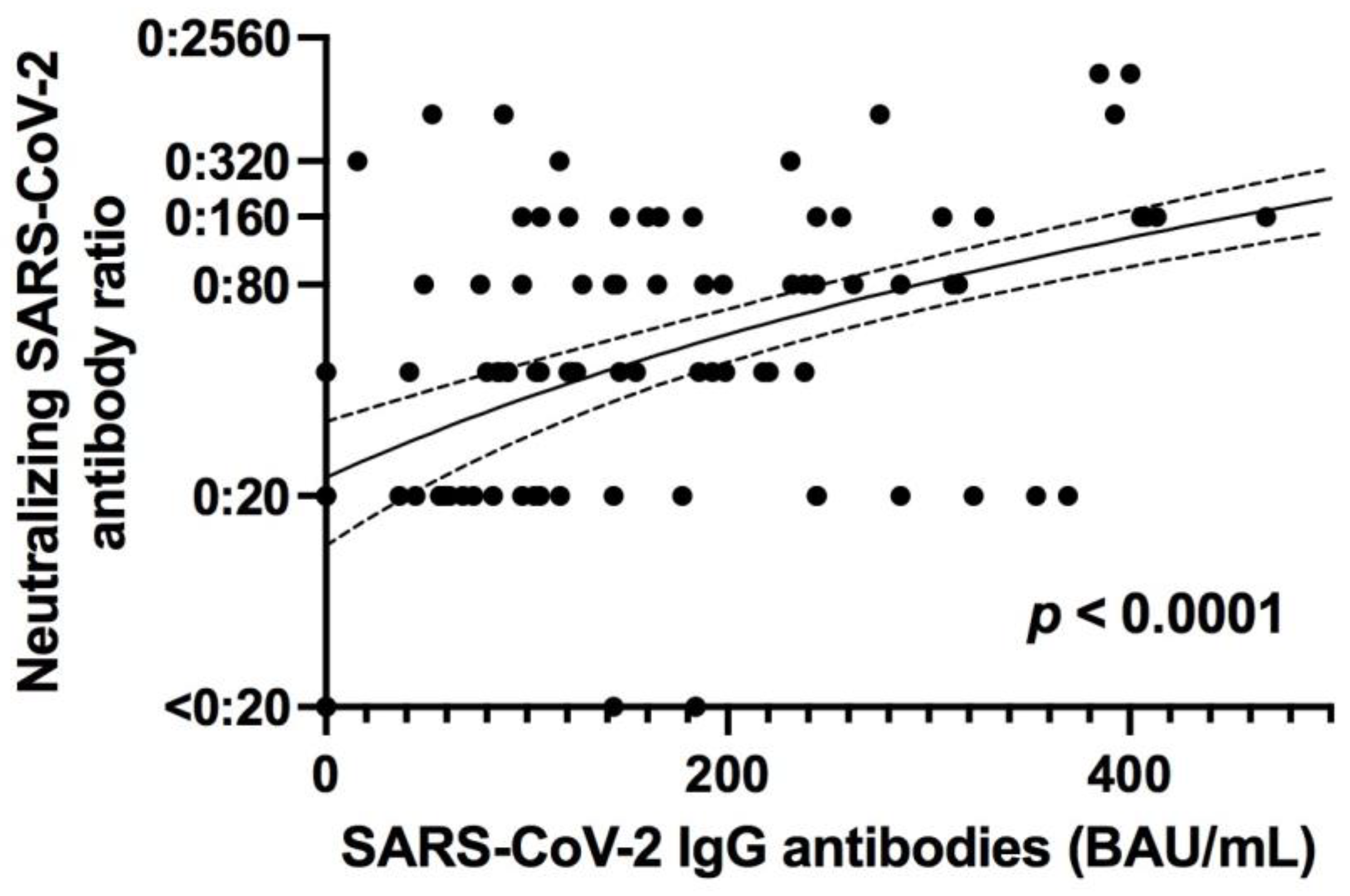
The neutralizing capability of antibodies against SARS-CoV-2 in patient serum was quantified using previously described methods [
11]. Serial dilutions (1:20–1:2560) of serum samples were incubated with 100 TCID50 of SARS-CoV-2 for 1 h at 37 °C and subsequently added to confluent Vero E6 cells cultured in 96-well microtiter plates. On day 2 after infection, cells were stained with crystal violet (Roth, Karlsruhe, Germany) and analyzed for the appearance of virus-induced cytopathic effects (CPE) by light microscopy. The neutralizing titer was defined as the reciprocal of the highest serum dilution at which no CPE breakthrough was observed in any of the triplicate cultures. SARS-CoV-2 specific neutralizing antibodies were evaluated in 176 out of 200 patients.
2.6. Statistics and Data Analysis
Statistical analyses were performed with IBM SPSS-Statistic 23 (SPSS Inc., Chicago, IL, USA) and GraphPad Prism 9 (GraphPad Software, San Diego, CA, USA) software. Data sets were analyzed with the Mann–Whitney U-test, the Kruskal–Wallis test, the Friedman test, or Spearman’s rank-order correlation tests. Univariate and multivariate linear regression models were used to estimate relationship between the independent variables. Two-sided p-values lower than 0.05 were considered significant.
4. Discussion
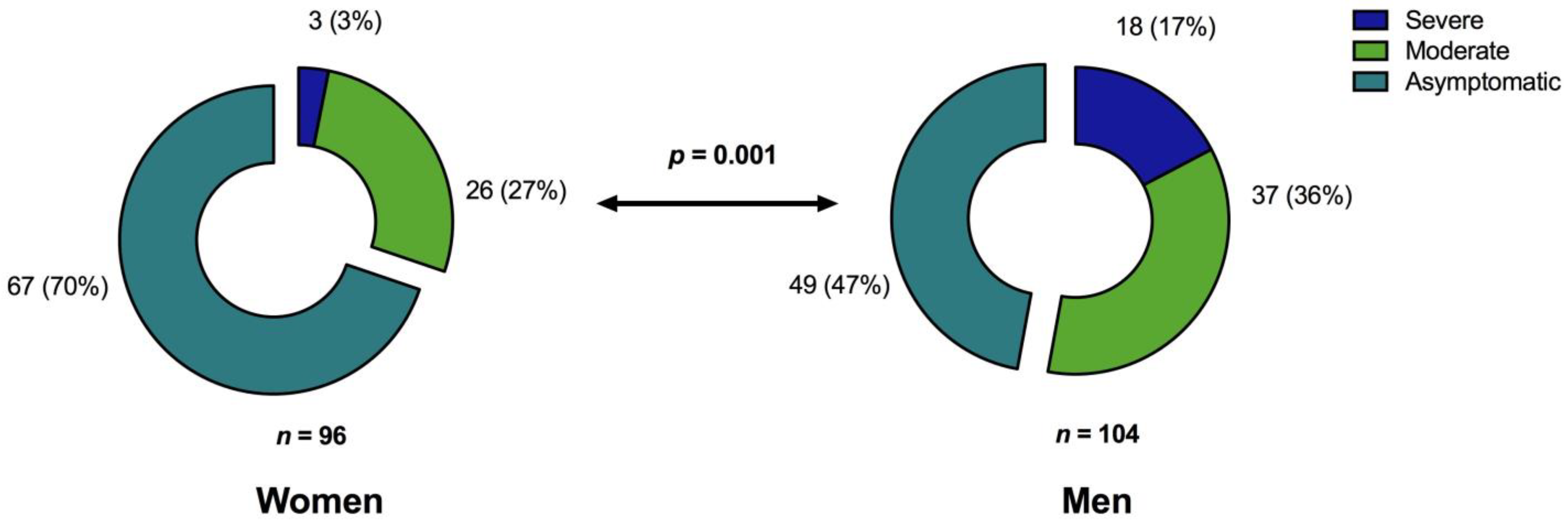
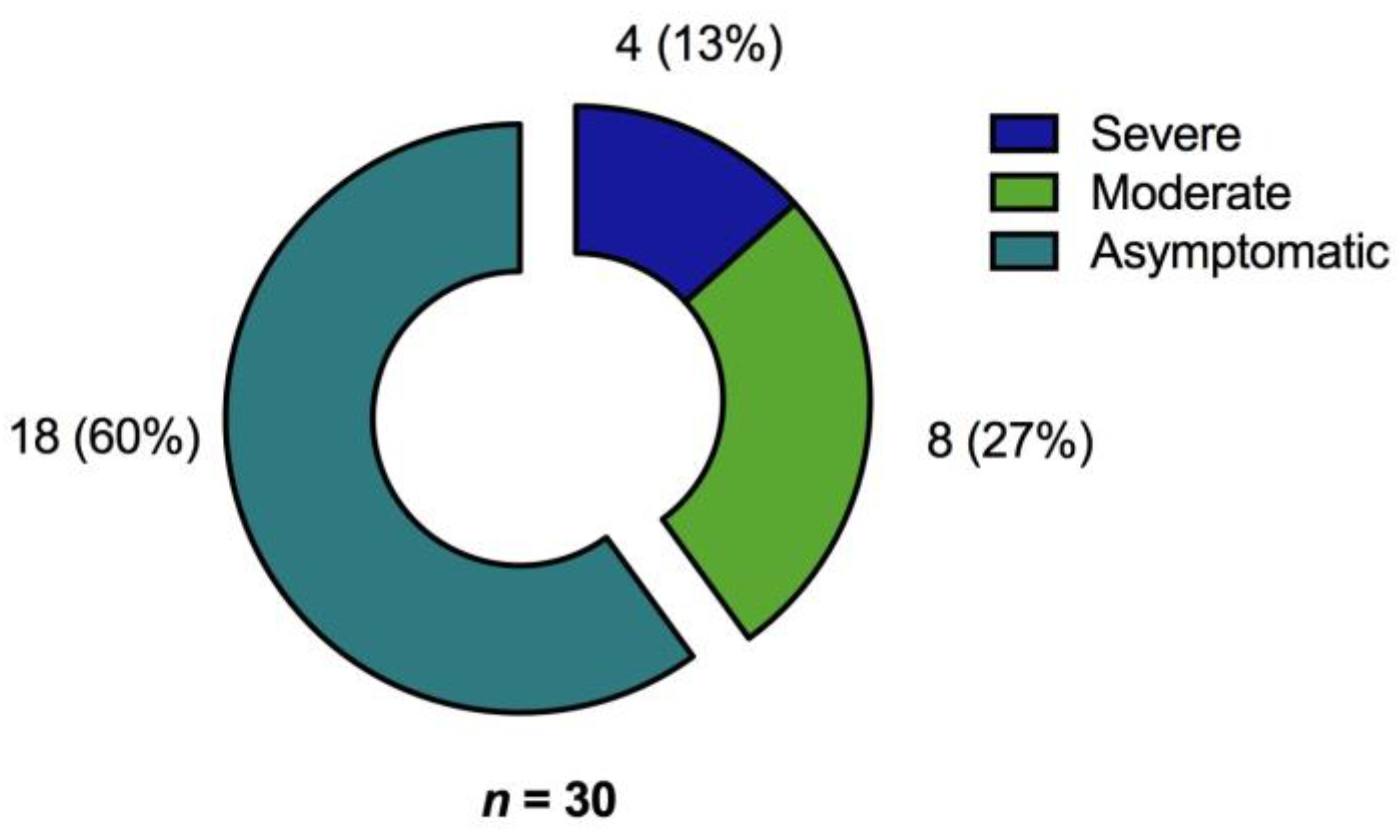
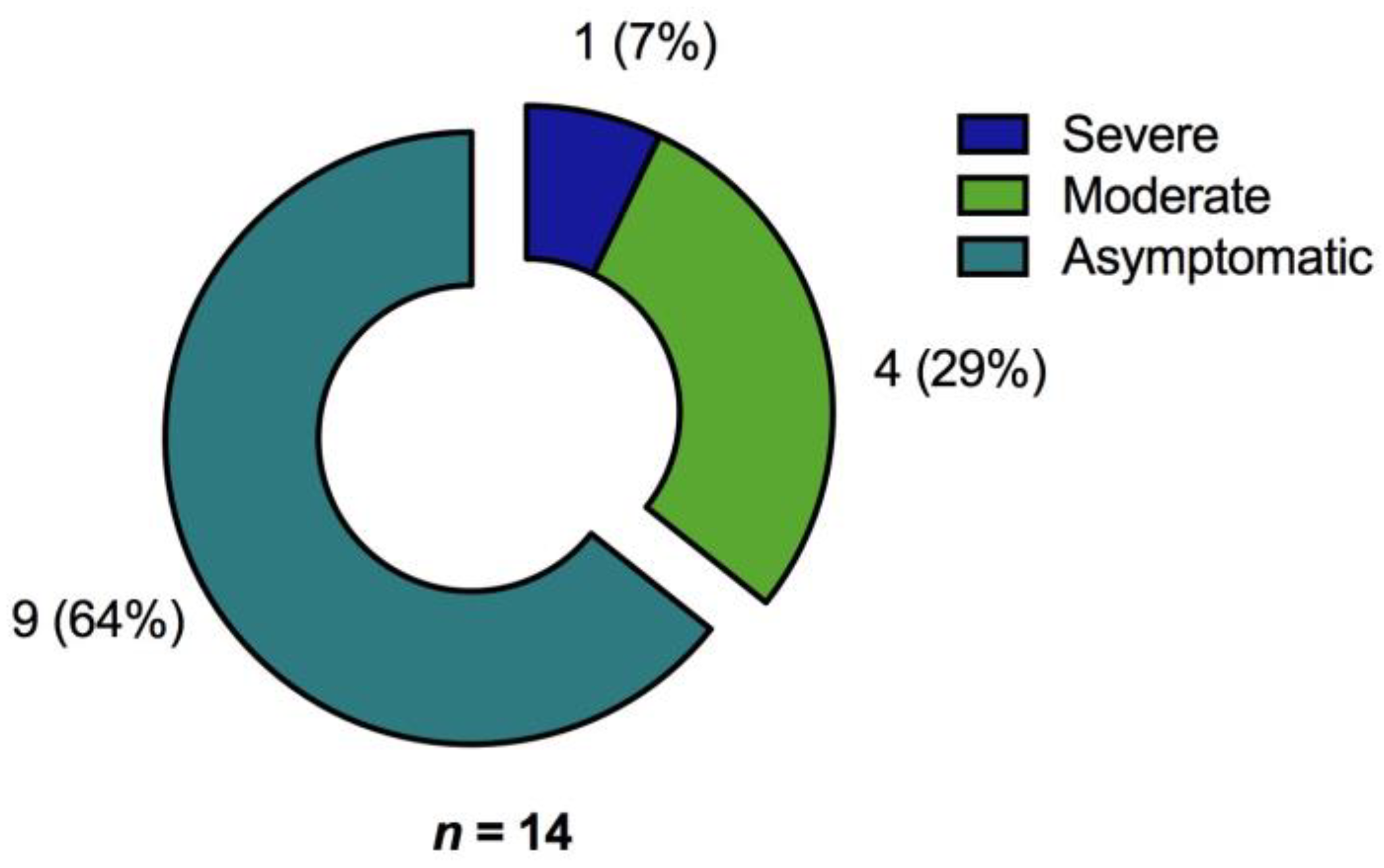
In a cohort of 200 unvaccinated COVID-19 convalescents, SARS-CoV-2 specific T cell- and humoral immunity was assessed up to almost one year after resolution of acute infection. The combined results of our study not only provide a comprehensive characterization of the immune responses after COVID-19 but also demonstrate that the severity of the initial disease predominantly affects cellular and humoral immunity against the Spike (S) protein even months after recovery from SARS-CoV-2 infection. Regarding Membrane (M) protein specific T cell immunity, patient age and time since SARS-CoV-2 infection were prognostic factors for long-term immunity. In addition, immunosuppression was an independent factor for the absence of T cell immunity. In our cohort, significant risk factors for hospitalization were older age, obesity, and male sex which is consistent with the findings of other studies [
15,
16,
17].
Knowledge of the extent and quality of the long-term immune system response to SARS-CoV-2 is crucial for further risk assessment regarding the ongoing pandemic. In addition, understanding the natural history of immunity can help estimate the duration of immunity after vaccination. Factors that trigger initial and long-term individual immunity still remain to be characterized. T cell specific responses are initially detected in almost all SARS-CoV-2 infections [
18,
19] and have been associated with control of primary SARS-CoV-2 infection [
19]. In our cohort, the main factor impairing the formation of T cell immune responses was an immunocompromised status of the patient. Thus, SARS-CoV-2-specific T cell responses (against the Spike (S) and Membrane (M) protein) were found significantly less frequently in immunocompromised patients, who are considered particularly at risk of severe forms of COVID-19 [
20,
21]. Consistent with our data, low T cell responses (and seroconversion rates) were observed in studies of various immunocompromised patient cohorts both during the natural course of SARS-CoV-2 infection and after vaccination [
12,
22,
23,
24,
25,
26]. However, because IFNγ ELISpot assays against three different viral Spike (S) peptide pools/proteins and one Membrane (M) protein pool provide the only read-out regarding T cell responses in our study, our findings may not provide comprehensive information about the complex overall T cell responses after primary SARS-CoV-2 infection [
24]. Few studies have focused on long-term T cell immunity after COVID-19 [
27,
28,
29]. Our results are consistent with those studies finding that T cell responses are detectable at least 6 months after initial infection, not only in symptomatic but also in asymptomatic subjects [
28].
Virus-specific antibodies develop in most patients infected with SARS-CoV-2 within 5 to 15 days of infection [
19,
30], and the S protein is the target of neutralizing SARS-CoV-2 antibodies [
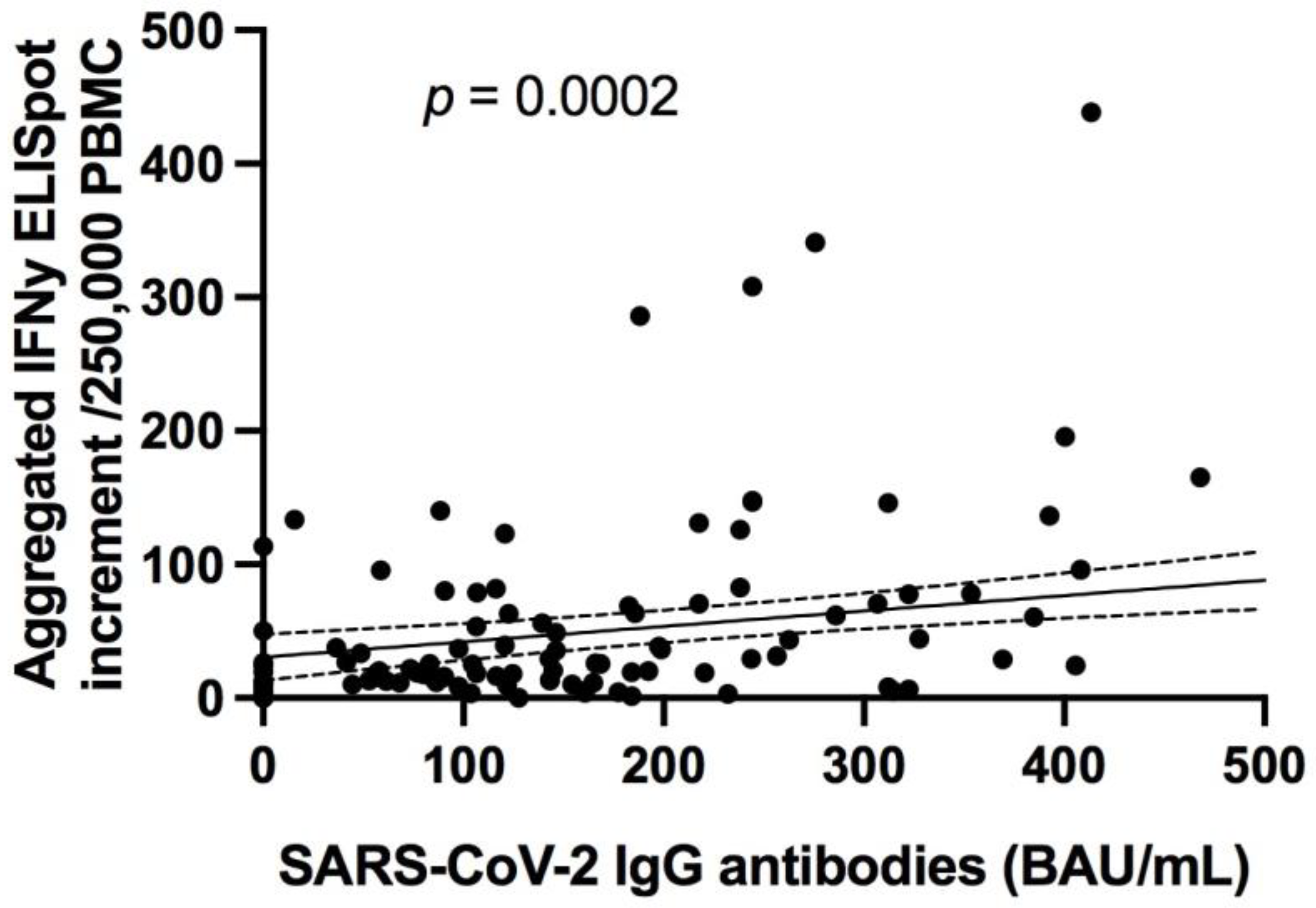
13]. Although the number of studies of the persistence of SARS-CoV-2-specific immunity is increasing, data on long-term immunity after SARS-CoV-2 infection are still scarce. In addition, most of these studies focus on the persistence of circulating anti-SARS-CoV-2 IgG antibodies. These studies demonstrate that seroconversion is maintained over several months, as seen in our cohort [
30,
31]. Classification of patients according to the severity of initial COVID-19 showed that those who had a SARS-CoV-2 infection with more severe clinical manifestations had a stronger Spike (S) protein specific T cell and antibody response. This finding is consistent with results from other groups showing stronger SARS-CoV-2 specific T cell responses in patients who had recovered from severe COVID-19, as well as from other coronavirus infections, such as SARS and MERS, in which patients with more severe symptoms have higher levels of specific memory T cells against the virus [
2,
32,
33]. Similar to the stronger T cell response observed in patients with acute COVID-19, stronger long-term T cell responses probably reflect a more severe course of disease with a stronger immunogenic environment created by higher viral load and inflammatory bystander activation [
34,
35]. In addition, neutralizing antibody titers and specific anti-S protein total antibody titers have been found to correlate with the severity of COVID-19 in our cohort and others [
19,
34,
36].
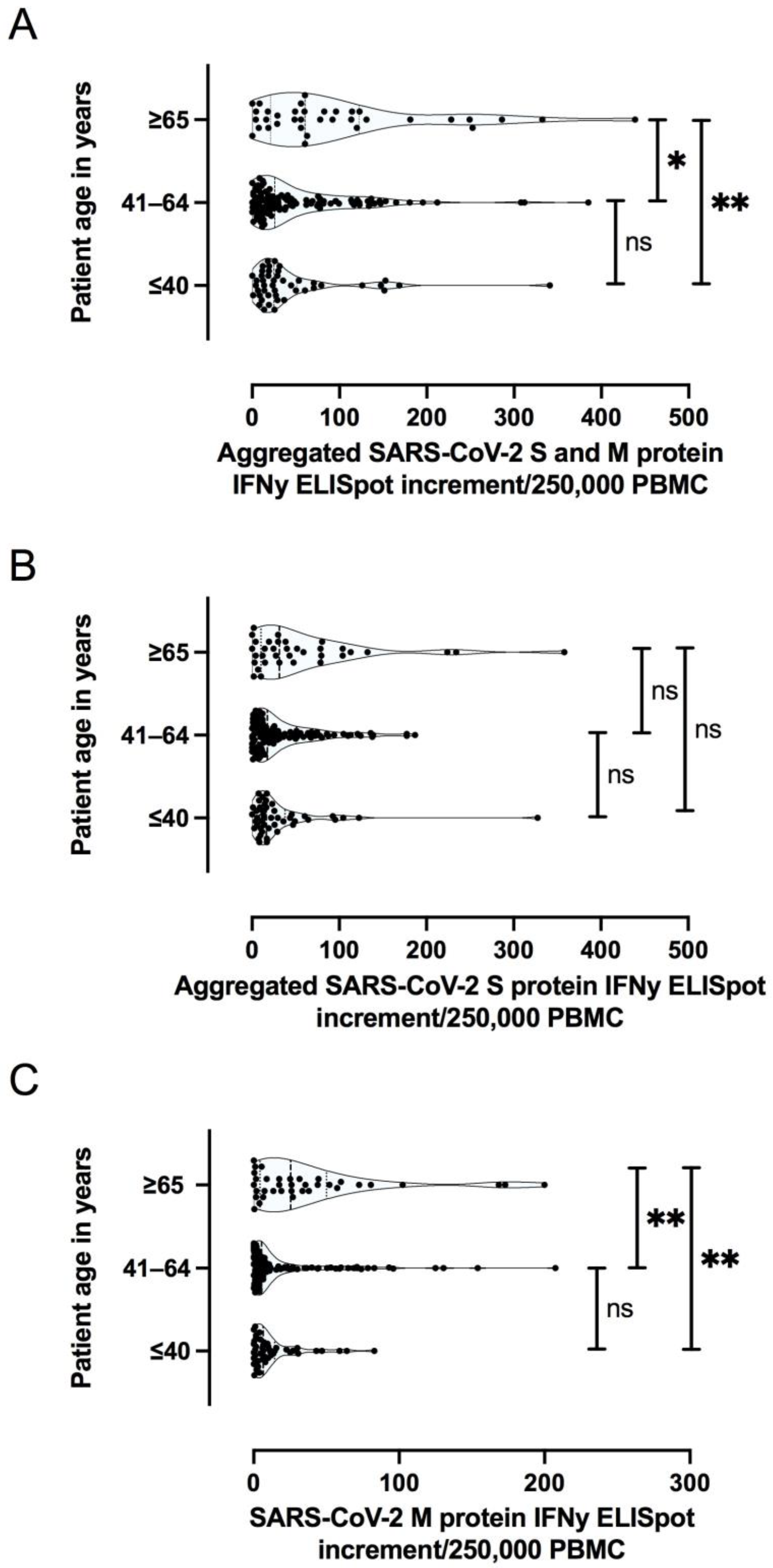
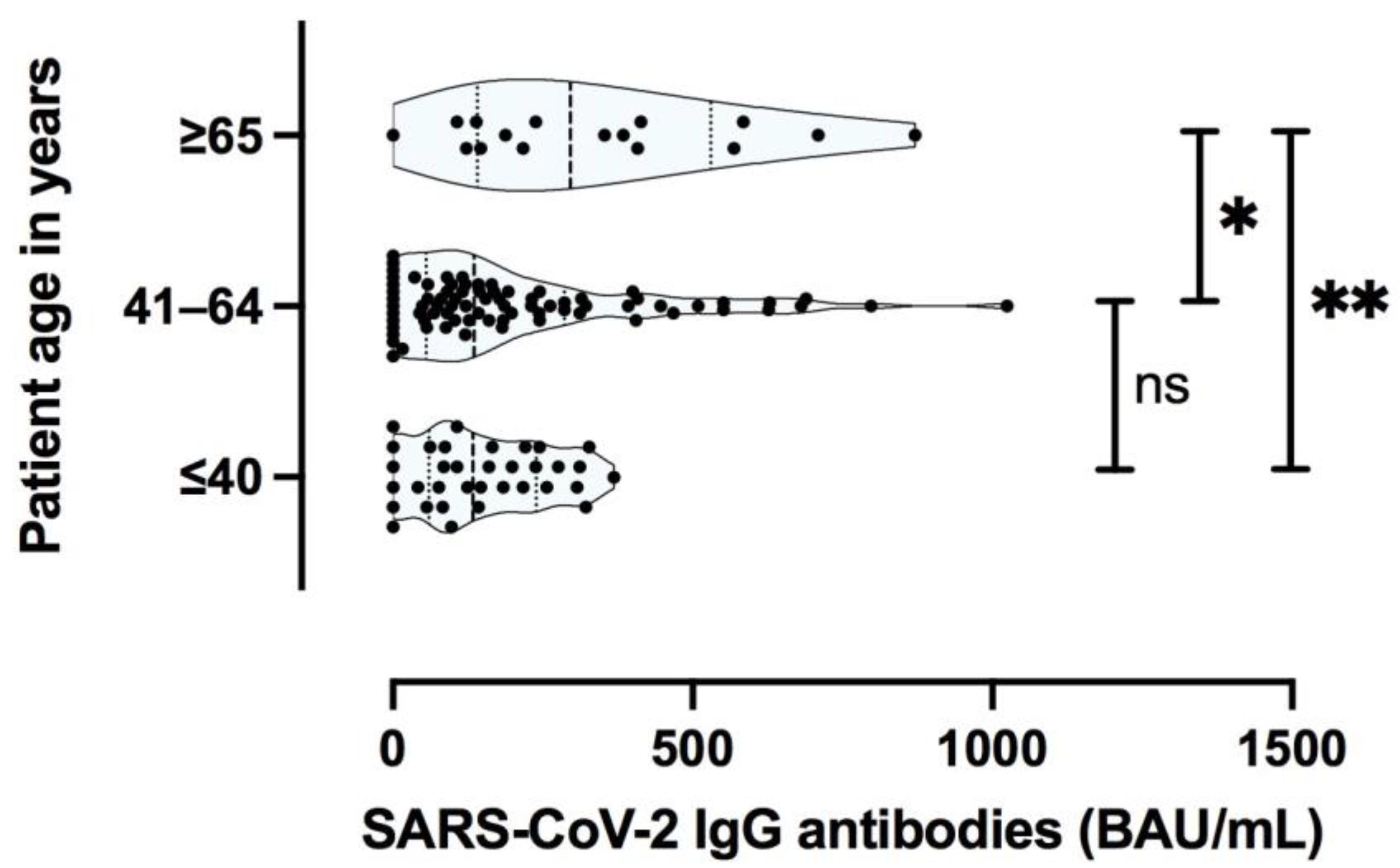
In addition to COVID-19 severity, patient age and concomitant diseases have been shown to affect SARS-CoV-2 specific cellular immunity during the acute phase of the disease [
37]. Typically, immune senescence predisposes older patients to infection and may hinder the development of protective immunity after immunization. Although the sample size in our study was limited, older participants showed a stronger T cell specific and antibody-specific immune response after COVID-19. Importantly, the age of the patient was an independent factor for the magnitude of T cell response (especially against the Membrane protein) and antibody responses, independent of the initial severity of COVID-19. This finding is in line with one previous study showing that even in advanced age, an immunological memory against SARS-CoV-2 was developed and was still present in most patients months after recovery [
27,
38,
39,
40,
41]. Another potential explanation may be that older patients may had more repetitive exposures to seasonal coronaviruses that can induce cross-reactive T cells and antibodies recognizing regions of the Membrane (M) or Spike (S) proteins, which are conserved among several coronaviruses [
42,
43]. Especially, the Membrane (M) protein accounting for the overall shape of the viral envelope is the most abundant structural protein of the coronavirus family. Studies have shown that different coronavirus M proteins share the same overall basic structural characteristics and are most subject to evolutionary constraints [
44,
45,
46]. A recent study of SARS-CoV-2 sequences confirmed this finding, showing that missense mutations in the Membrane protein gene were relatively uncommon [
47]. Studies have shown that the Membrane protein can elicit similar immune responses as the S protein, making it a potential candidate for vaccines [
48,
49,
50].
The current study has some limitations, mainly because longitudinal data for individual subjects are missing. In addition, all patients in this study survived their SARS-CoV-2 infection and thus were more likely to develop virus-specific immunity than patients who died as consequence of infection [
37]. Nevertheless, this current prospective cross-sectional study describes the long-term persistence of cellular and humoral immunity to SARS-CoV-2, which is likely to be present for several months in the vast majority of adults after COVID-19. These characteristics are encouraging with respect to the longevity of cellular immunity to this novel virus and likely contribute to the relatively low rates of re-infection observed to date.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}