
Comparison of Glaucoma-Relevant Transcriptomic Datasets Identifies Novel Drug Targets for Retinal Ganglion Cell Neuroprotection


Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA-Sequencing Comparison and Identification of Compounds via the Comparative Toxicogenomics Database
2.2. Animal Strain and Husbandry
2.3. Retina Axotomy Explant Model
2.4. Immunofluorescent Labelling
2.5. Analysis of Retinal Ganglion Cell Degeneration
2.6. Statistical Analysis
3. Results
3.1. Comparison of Glaucoma-Relevant Transcriptomic Datasets Identifies Common Genes for Therapeutic Targeting
3.2. Retinal Explant Model Provides a Platform to Rapidly Test Candidate Neuroprotective Therapeutics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stein, J.D.; Khawaja, A.P.; Weizer, J.S. Glaucoma in adults-screening, diagnosis, and management: A review. JAMA J. Am. Med. Assoc. 2021, 325, 164–174. [Google Scholar] [CrossRef]
- Tham, Y.-C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.-Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24974815 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Yasuda, M.; Tanaka, Y.; Ryu, M.; Tsuda, S.; Nakazawa, T. Rna sequence reveals mouse retinal transcriptome changes early after axonal injury. PLoS ONE 2014, 9, e93258. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24676137 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Yasuda, M.; Tanaka, Y.; Omodaka, K.; Nishiguchi, K.M.; Nakamura, O.; Tsuda, S.; Nakazawa, T. Transcriptome profiling of the rat retina after optic nerve transection. Sci. Rep. 2016, 6, 28736. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27353354 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Morgan, E.J.; Tribble, J.R. Microbead models in glaucoma. Exp. Eye Res. 2015, 141, 9–14. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26116904 (accessed on 26 August 2021). [CrossRef]
- Tribble, R.J.; Otmani, A.; Kokkali, E.; Lardner, E.; Morgan, J.E.; Williams, P.A. Retinal ganglion cell degeneration in a rat magnetic bead model of ocular hypertensive glaucoma. Transl. Vis. Sci. Technol. 2021, 10, 21. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33510960 (accessed on 26 August 2021). [CrossRef]
- Williams, P.A.; Marsh-Armstrong, N.; Howell, G.R.; Bosco, A.; Danias, J.; Simon, J.; Di Polo, A.; Kuehn, M.; Przedborski, S.; Raff, M.; et al. Neuroinflammation in glaucoma: A new opportunity. Exp. Eye Res. 2017, 157, 20–27. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28242160 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Williams, A.P.; Harder, J.M.; Foxworth, N.E.; Cochran, K.E.; Philip, V.M.; Porciatti, V.; Smithies, O.; John, S.W. Vitamin b. Science 2017, 355, 756–760. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28209901 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Tribble, R.J.; Otmani, A.; Sun, S.; Ellis, S.A.; Cimaglia, G.; Vohra, R.; Jöe, M.; Lardner, E.; Venkataraman, A.P.; Domínguez-Vicent, A.; et al. Nicotinamide provides neuroprotection in glaucoma by protecting against mitochondrial and metabolic dysfunction. Redox Biol. 2021, 43, 101988. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33932867 (accessed on 26 August 2021). [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22743772 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Bull, D.N.; Johnson, T.V.; Welsapar, G.; DeKorver, N.W.; Tomarev, S.I.; Martin, K.R. Use of an adult rat retinal explant model for screening of potential retinal ganglion cell neuroprotective therapies. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3309–3320. Available online: https://www.ncbi.nlm.nih.gov/pubmed/21345987 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Williams, A.P.; Howell, G.R.; Barbay, J.M.; Braine, C.E.; Sousa, G.L.; John, S.W.; Morgan, J.E. Retinal ganglion cell dendritic atrophy in dba/2j glaucoma. PLoS ONE 2013, 8, e72282. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23977271 (accessed on 26 August 2021). [CrossRef]
- Donahue, J.R.; Moller-Trane, R.; Nickells, R.W. Meta-analysis of transcriptomic changes in optic nerve injury and neurodegenerative models reveals a fundamental response to injury throughout the central nervous system. Mol. Vis. 2017, 23, 987–1005. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29386873 (accessed on 26 August 2021).
- Wang, J.; Struebing, F.L.; Geisert, E.E. Commonalities of optic nerve injury and glaucoma-induced neurodegeneration: Insights from transcriptome-wide studies. Exp. Eye Res. 2021, 207, 108571. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33844961 (accessed on 26 August 2021). [CrossRef]
- Biermann, J.; Grieshaber, P.; Goebel, U.; Martin, G.; Thanos, S.; di Giovanni, S.; Lagrèze, W.A. Valproic acid-mediated neuroprotection and regeneration in injured retinal ganglion cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 526–534. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19628741 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Kimura, A.; Guo, X.; Noro, T.; Harada, C.; Tanaka, K.; Namekata, K.; Harada, T. Valproic acid prevents retinal degeneration in a murine model of normal tension glaucoma. Neurosci. Lett. 2015, 588, 108–113. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25555796 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Lindsey, D.J.; Weinreb, R.N. Survival and differentiation of purified retinal ganglion cells in a chemically defined microenvironment. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3640–3648. Available online: https://www.ncbi.nlm.nih.gov/pubmed/7916337 (accessed on 26 August 2021).
- Allen, S.R.; Olsen, T.W.; Sayeed, I.; Cale, H.A.; Morrison, K.C.; Oumarbaeva, Y.; Lucaciu, I.; Boatright, J.H.; Pardue, M.T.; Stein, D.G. Progesterone treatment in two rat models of ocular ischemia. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2880–2891. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26024074 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Jackson, C.A.; Roche, S.L.; Byrne, A.M.; Ruiz-Lopez, A.M.; Cotter, T.G. Progesterone receptor signalling in retinal photoreceptor neuroprotection. J. Neurochem. 2016, 136, 63–77. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26447367 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Jiang, M.; Ma, X.; Zhao, Q.; Li, Y.; Xing, Y.; Deng, Q.; Shen, Y. The neuroprotective effects of novel estrogen receptor gper1 in mouse retinal ganglion cell degeneration. Exp. Eye Res. 2019, 189, 107826. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31586450 (accessed on 26 August 2021). [CrossRef]
- Means, C.J.; Lopez, A.A.; Koulen, P. Estrogen protects optic nerve head astrocytes against oxidative stress by preventing caspase-3 activation, tau dephosphorylation at ser. Cell Mol. Neurobiol. 2021, 41, 449–458. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32385548 (accessed on 26 August 2021). [CrossRef]
- Hao, M.; Li, Y.; Lin, W.; Xu, Q.; Shao, N.; Zhang, Y.; Kuang, H. Estrogen prevents high-glucose-induced damage of retinal ganglion cells via mitochondrial pathway. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 83–90. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25216739 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Prokai-Tatrai, K.; Xin, H.; Nguyen, V.; Szarka, S.; Blazics, B.; Prokai, L.; Koulen, P. 17β-estradiol eye drops protect the retinal ganglion cell layer and preserve visual function in an in vivo model of glaucoma. Mol. Pharm. 2013, 10, 3253–3261. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23841874 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Li, H.; Wang, B.; Zhu, C.; Feng, Y.; Wang, S.; Shahzad, M.; Hu, C.; Mo, M.; Du, F.; Yu, X. 17β-estradiol impedes bax-involved mitochondrial apoptosis of retinal nerve cells induced by oxidative damage via the phosphatidylinositol 3-kinase/akt signal pathway. J. Mol. Neurosci. 2013, 50, 482–493. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23361188 (accessed on 26 August 2021). [CrossRef]
- Yu, X.; Rajala, R.V.; McGinnis, J.F.; Li, F.; Anderson, R.E.; Yan, X.; Li, S.; Elias, R.V.; Knapp, R.R.; Zhou, X.; et al. Involvement of insulin/phosphoinositide 3-kinase/akt signal pathway in 17 beta-estradiol-mediated neuroprotection. J. Biol. Chem. 2004, 279, 13086–13094. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14711819 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Newman-Casey, A.P.; Talwar, N.; Nan, B.; Musch, D.C.; Pasquale, L.R.; Stein, J.D. The potential association between postmenopausal hormone use and primary open-angle glaucoma. JAMA Ophthalmol. 2014, 132, 298–303. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24481323 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Kumar, M.D.; Perez, E.; Cai, Z.Y.; Aoun, P.; Brun-Zinkernagel, A.M.; Covey, D.F.; Simpkins, J.W.; Agarwal, N. Role of nonfeminizing estrogen analogues in neuroprotection of rat retinal ganglion cells against glutamate-induced cytotoxicity. Free Radic. Biol. Med. 2005, 38, 1152–1163. Available online: https://www.ncbi.nlm.nih.gov/pubmed/15808412 (accessed on 26 August 2021). [CrossRef]
- Zhou, X.; Li, F.; Ge, J.; Sarkisian, S.R.; Tomita, H.; Zaharia, A.; Chodosh, J.; Cao, W. Retinal ganglion cell protection by 17-beta-estradiol in a mouse model of inherited glaucoma. Dev. Neurobiol. 2007, 67, 603–616. Available online: https://www.ncbi.nlm.nih.gov/pubmed/17443811 (accessed on 26 August 2021). [CrossRef]
- Merrell, B.J.; McMurry, J.P. Folic Acid. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, January 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554487/ (accessed on 26 August 2021).
- Cianciulli, A.; Salvatore, R.; Porro, C.; Trotta, T.; Panaro, M.A. Folic acid is able to polarize the inflammatory response in lps activated microglia by regulating multiple signaling pathways. Mediat. Inflamm. 2016, 2016, 5240127. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27738387 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Li, W.; Ma, Y.; Li, Z.; Lv, X.; Wang, X.; Zhou, D.; Luo, S.; Wilson, J.X.; Huang, G. Folic acid decreases astrocyte apoptosis by preventing oxidative stress-induced telomere attrition. Int. J. Mol. Sci. 2019, 21, 62. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31861819 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Meng, Q.Q.; Lei, W.; Chen, H.; Feng, Z.C.; Hu, L.Q.; Zhang, X.L.; Li, S. Combined rosiglitazone and forskolin have neuroprotective effects in sd rats after spinal cord injury. PPAR Res. 2018, 2018, 3897478. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30034460 (accessed on 26 August 2021). [CrossRef] [PubMed]
- He, X.; Feng, L.; Meng, H.; Wang, X.; Liu, S. Rosiglitazone protects dopaminergic neurons against lipopolysaccharide-induced neurotoxicity through inhibition of microglia activation. Int. J. Neurosci. 2012, 122, 532–540. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22524690 (accessed on 26 August 2021). [CrossRef]
- Li, P.; Xu, X.; Zheng, Z.; Zhu, B.; Shi, Y.; Liu, K. Protective effects of rosiglitazone on retinal neuronal damage in diabetic rats. Curr. Eye Res. 2011, 36, 673–679. Available online: https://www.ncbi.nlm.nih.gov/pubmed/21599458 (accessed on 26 August 2021). [CrossRef] [PubMed]
- Yin, C.J.; Zhang, L.; Ma, N.X.; Wang, Y.; Lee, G.; Hou, X.Y.; Lei, Z.F.; Zhang, F.Y.; Dong, F.P.; Wu, G.Y.; et al. Chemical conversion of human fetal astrocytes into neurons through modulation of multiple signaling pathways. Stem. Cell Rep. 2019, 12, 488–501. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30745031 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Ueda, K.; Nakahara, T.; Mori, A.; Sakamoto, K.; Ishii, K. Protective effects of tgf-β inhibitors in a rat model of nmda-induced retinal degeneration. Eur. J. Pharm. 2013, 699, 188–193. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23220705 (accessed on 26 August 2021). [CrossRef]
- Tappeiner, C.; Maurer, E.; Sallin, P.; Bise, T.; Enzmann, V.; Tschopp, M. Inhibition of the tgfβ pathway enhances retinal regeneration in adult zebrafish. PLoS ONE 2016, 11, e0167073. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27880821 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Zolezzi, M.J.; Silva-Alvarez, C.; Ordenes, D.; Godoy, J.A.; Carvajal, F.J.; Santos, M.J.; Inestrosa, N.C. Peroxisome proliferator-activated receptor (ppar) γ and pparα agonists modulate mitochondrial fusion-fission dynamics: Relevance to reactive oxygen species (ros)-related neurodegenerative disorders? PLoS ONE 2013, 8, e64019. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23675519 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Yakisich, S.J.; Lindblom, I.O.; Siden, A.; Cruz, M.H. Rapid inhibition of ongoing dna synthesis in human glioma tissue by genistein. Oncol. Rep. 2009, 22, 569–574. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19639205 (accessed on 26 August 2021). [CrossRef] [Green Version]
- Markovits, J.; Larsen, A.K.; Ségal-Bendirdjian, E.; Fossé, P.; Saucier, J.M.; Gazit, A.; Levitzki, A.; Umezawa, K.; Jacquemin-Sablon, A. Inhibition of dna topoisomerases i and ii and induction of apoptosis by erbstatin and tyrphostin derivatives. Biochem. Pharmacol. 1994, 48, 549–560. Available online: https://www.ncbi.nlm.nih.gov/pubmed/8068042 (accessed on 26 August 2021). [CrossRef]
- Pereira, P.S.; Medina, S.V.; Araujo, E.G. Cholinergic activity modulates the survival of retinal ganglion cells in culture: The role of m1 muscarinic receptors. Int. J. Dev. Neurosci. 2001, 19, 559–567. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11600318 (accessed on 26 August 2021). [CrossRef]
- Santos, C.R.; Araujo, E.G. Cyclic amp increases the survival of ganglion cells in mixed retinal cell cultures in the absence of exogenous neurotrophic molecules, an effect that involves cholinergic activity. Braz. J. Med. Biol. Res. 2001, 34, 1585–1593. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11717712 (accessed on 26 August 2021). [CrossRef] [PubMed] [Green Version]
- Pereira, S.F.P.; de Araujo, E.G. Chronic depolarization induced by veratridine increases the survival of rat retinal ganglion cells ‘in vitro’. Int. J. Dev. Neurosci. 2000, 18, 773–780. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11154846 (accessed on 26 August 2021). [CrossRef]



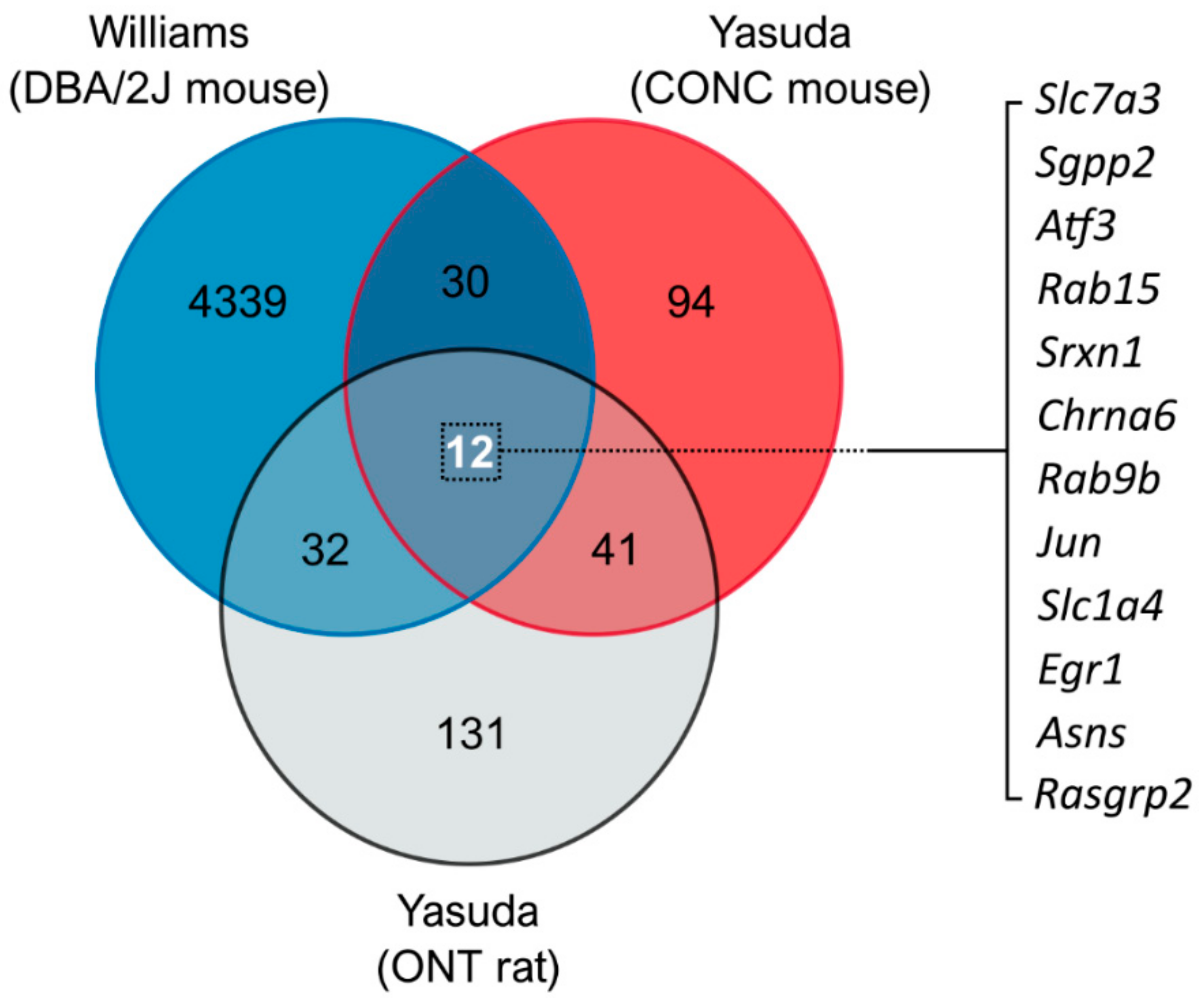{kind=link}
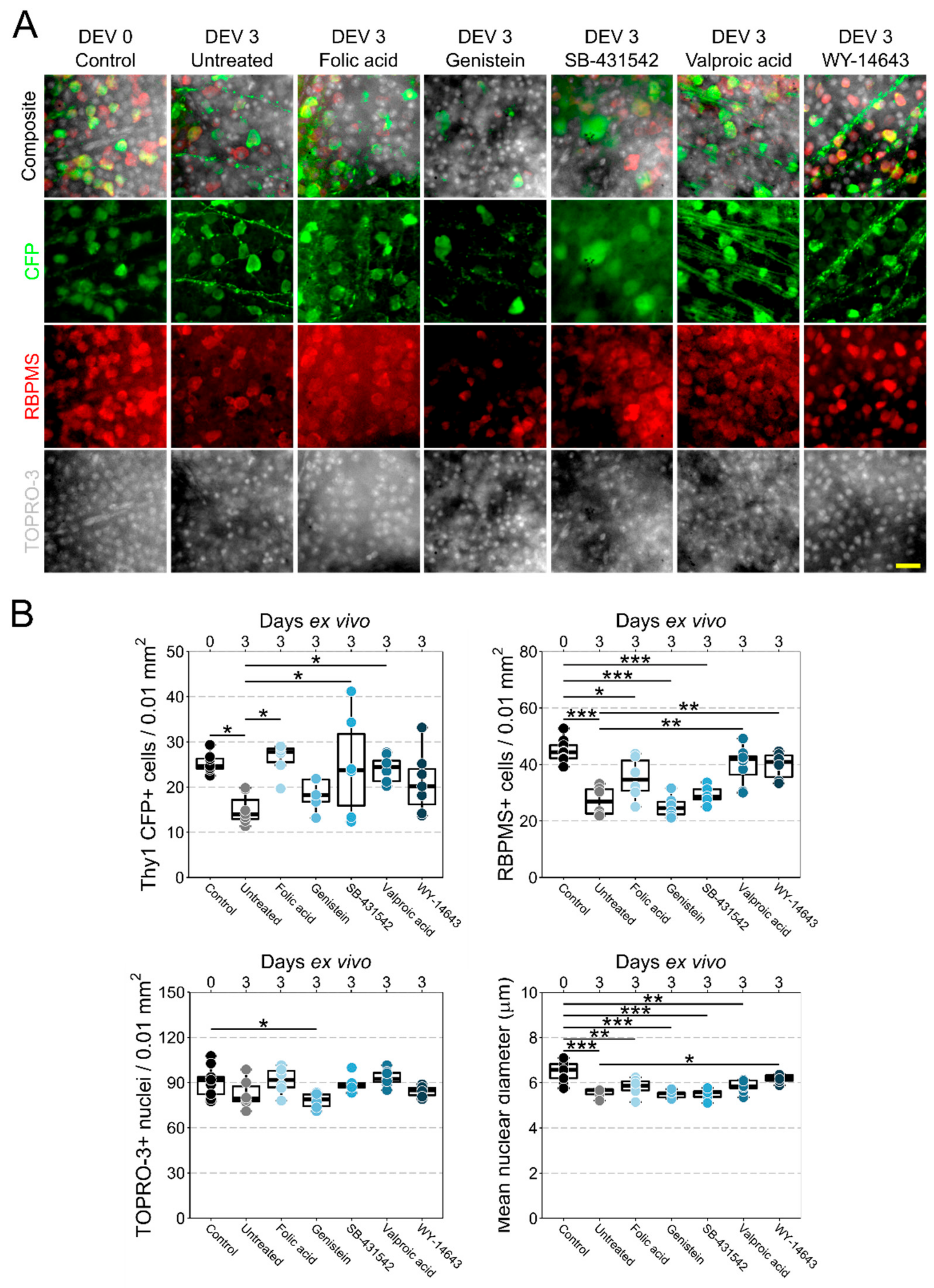{kind=link}
| Antibody | Target | Host | Stock Conc. | Working Conc. | Dilution | Details |
|---|---|---|---|---|---|---|
| anti-RBPMS | RNA-binding protein, RGC specific in the retina | Rabbit | 660 µg/mL | 1.32 µg/mL | 1:500 | Novusbio NBP2-20112 |
| anti-GFP | XFPs (e.g., CFP) | Chicken | 10 mg/mL | 20 µg/mL | 1:500 | Abcam ab13970 |
| Goat-anti Rabbit Alexa Fluor 568 | Rabbit primary antibody | Goat | 2 mg/mL | 4 µg/mL | 1:500 | Invitrogen A11011 |
| Goat-anti Chick Alexa Fluor 488 | Chick primary antibody | Goat | 2 mg/mL | 4 µg/mL | 1:500 | Invitrogen A11039 |
| Ingenuity Canonical Pathways | −log(p) * | Molecules | Pathway Hits (%) | |
|---|---|---|---|---|
| D2 † vs. Mouse CONC | Complement System | 6.22 | ITGAM,C1QA,C1QC,C1QB | 10.8 |
| Neuroinflammation Signaling Pathway | 5.87 | JUN,AGER,TYROBP,HLA-A,KCNJ5,ATF4,CX3CR1 | 2.25 | |
| Dendritic Cell Maturation | 4.59 | TYROBP,HLA-A,FCER1G,ATF4,PLCB1 | 2.59 | |
| PI3K Signaling in B Lymphocytes | 4.04 | ATF3,JUN,ATF4,PLCB1 | 3.08 | |
| GNRH Signaling | 3.87 | JUN,EGR1,ATF4,PLCB1 | 2.78 | |
| Role of NFAT in Regulation of the Immune Response | 3.44 | JUN,HLA-A,FCER1G,PLCB1 | 2.15 | |
| OX40 Signaling Pathway | 3.21 | JUN,HLA-A,FCER1G | 3.3 | |
| Type I Diabetes Mellitus Signaling | 2.96 | HLA-A,FCER1G,PTPRN | 2.7 | |
| Cytotoxic T Lymphocyte-Mediated Apoptosis of Target Cells | 2.81 | HLA-A,FCER1G | 6.25 | |
| Phagosome Formation | 2.77 | ITGAM,FCER1G,PLCB1 | 2.31 | |
| D2 † vs. Rat ONT | Glutamate Receptor Signaling | 3.72 | GRIN2A,SLC1A4,GRIK3 | 5.26 |
| Amyotrophic Lateral Sclerosis Signaling | 2.87 | PRPH,GRIN2A,GRIK3 | 2.7 | |
| Asparagine Biosynthesis I | 2.71 | ASNS | 100 | |
| MIF Regulation of Innate Immunity | 2.49 | JUN,PLA2G5 | 4.65 | |
| Serotonin Receptor Signaling | 2.49 | HTR5A,HTR1D | 4.65 | |
| ATM Signaling | 1.8 | JUN,GADD45G | 2.04 | |
| Sphingosine and Sphingosine-1-Phosphate Metabolism | 1.76 | SGPP2 | 11.1 | |
| IGF-1 Signaling | 1.73 | JUN,YWHAH | 1.89 | |
| p53 Signaling | 1.7 | JUN,GADD45G | 1.8 | |
| Neuroinflammation Signaling Pathway | 1.64 | GRIN2A,JUN,PLA2G5 | 0.965 | |
| Mouse CONC vs. Rat ONT | AMPK Signaling | 2.76 | RAB9B,CHRNA6,CDKN1A,CHRNB3 | 1.85 |
| Asparagine Biosynthesis I | 2.62 | ASNS | 100 | |
| PI3K Signaling in B Lymphocytes | 2.43 | ATF3,JUN,ATF5 | 2.31 | |
| CXCR4 Signaling | 2.14 | RHOQ,JUN,EGR1 | 1.82 | |
| eNOS Signaling | 2.09 | CHRNA6,SLC7A1,CHRNB3 | 1.74 | |
| Heme Degradation | 2.02 | HMOX1 | 25 | |
| IL-8 Signaling | 1.93 | HMOX1,RHOQ,JUN | 1.52 | |
| IL-10 Signaling | 1.93 | HMOX1,JUN | 2.9 | |
| Tetrahydrofolate Salvage from 5,10-Methenyltetrahydrofolate | 1.93 | MTHFD2 | 20 | |
| Serine Biosynthesis | 1.93 | PHGDH | 1.85 | |
| Three-way comparison | Asparagine Biosynthesis I | 3.25 | ASNS | 100 |
| PI3K Signaling in B Lymphocytes | 2.63 | ATF3,JUN | 1.54 | |
| GNRH Signaling | 2.55 | JUN,EGR1 | 1.39 | |
| CXCR4 Signaling | 2.43 | JUN,EGR1 | 1.21 | |
| B Cell Receptor Signaling | 2.31 | JUN,EGR1 | 1.05 | |
| Sphingosine and Sphingosine-1-Phosphate Metabolism | 2.3 | SGPP2 | 11.1 | |
| AMPK Signaling | 2.2 | RAB9B,CHRNA6 | 0.926 | |
| IL-17A Signaling in Gastric Cells | 1.86 | JUN | 4 | |
| TNFR2 Signaling | 1.78 | JUN | 3.33 | |
| 4-1BB Signaling in T Lymphocytes | 1.75 | JUN | 3.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enz, T.J.; Tribble, J.R.; Williams, P.A. Comparison of Glaucoma-Relevant Transcriptomic Datasets Identifies Novel Drug Targets for Retinal Ganglion Cell Neuroprotection. J. Clin. Med. 2021, 10, 3938. https://doi.org/10.3390/jcm10173938
Enz TJ, Tribble JR, Williams PA. Comparison of Glaucoma-Relevant Transcriptomic Datasets Identifies Novel Drug Targets for Retinal Ganglion Cell Neuroprotection. Journal of Clinical Medicine. 2021; 10(17):3938. https://doi.org/10.3390/jcm10173938
Chicago/Turabian StyleEnz, Tim J., James R. Tribble, and Pete A. Williams. 2021. "Comparison of Glaucoma-Relevant Transcriptomic Datasets Identifies Novel Drug Targets for Retinal Ganglion Cell Neuroprotection" Journal of Clinical Medicine 10, no. 17: 3938. https://doi.org/10.3390/jcm10173938