
The Role of Dendritic Cells during Physiological and Pathological Dentinogenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
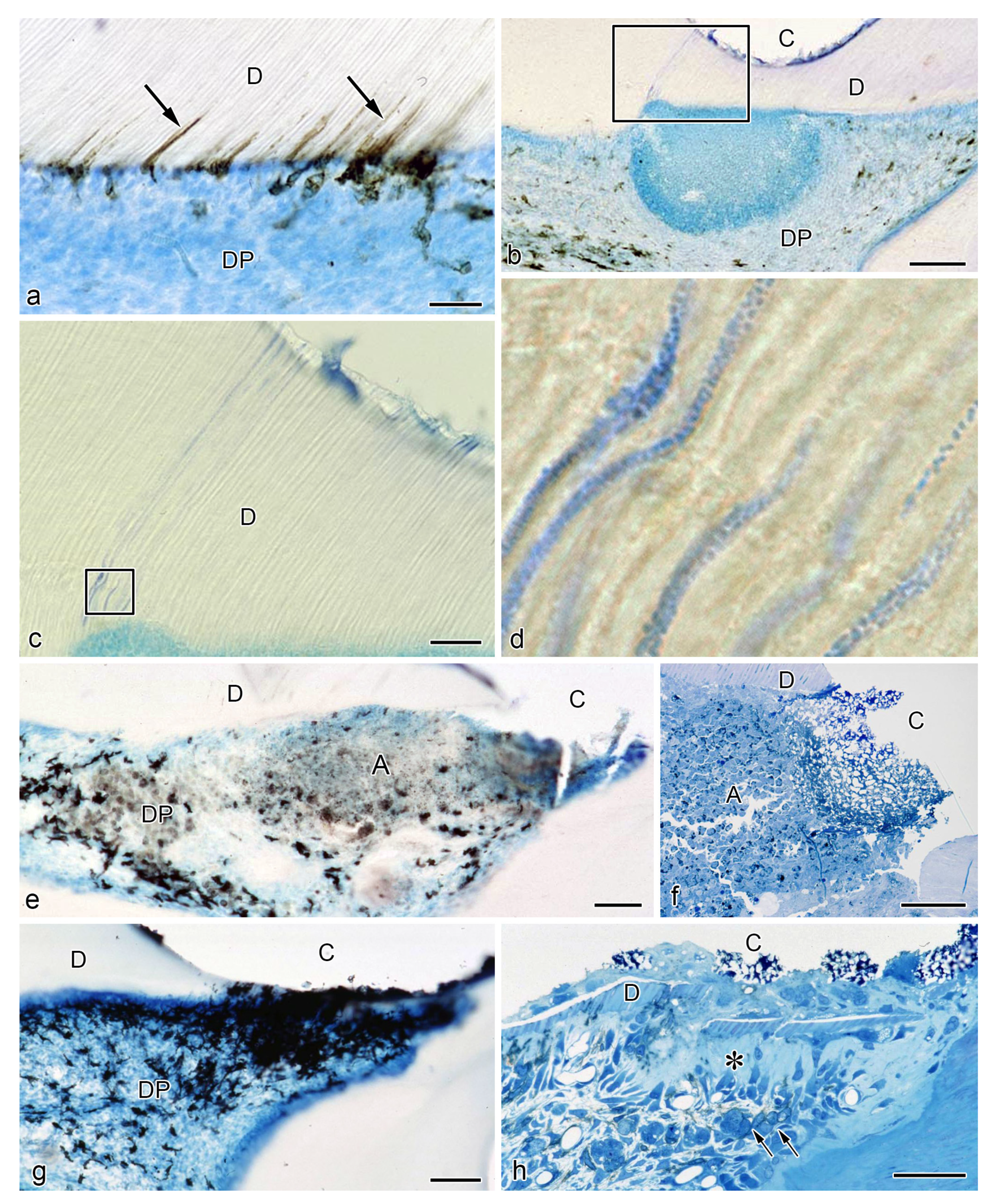{kind=link}
{kind=link}
{kind=link}
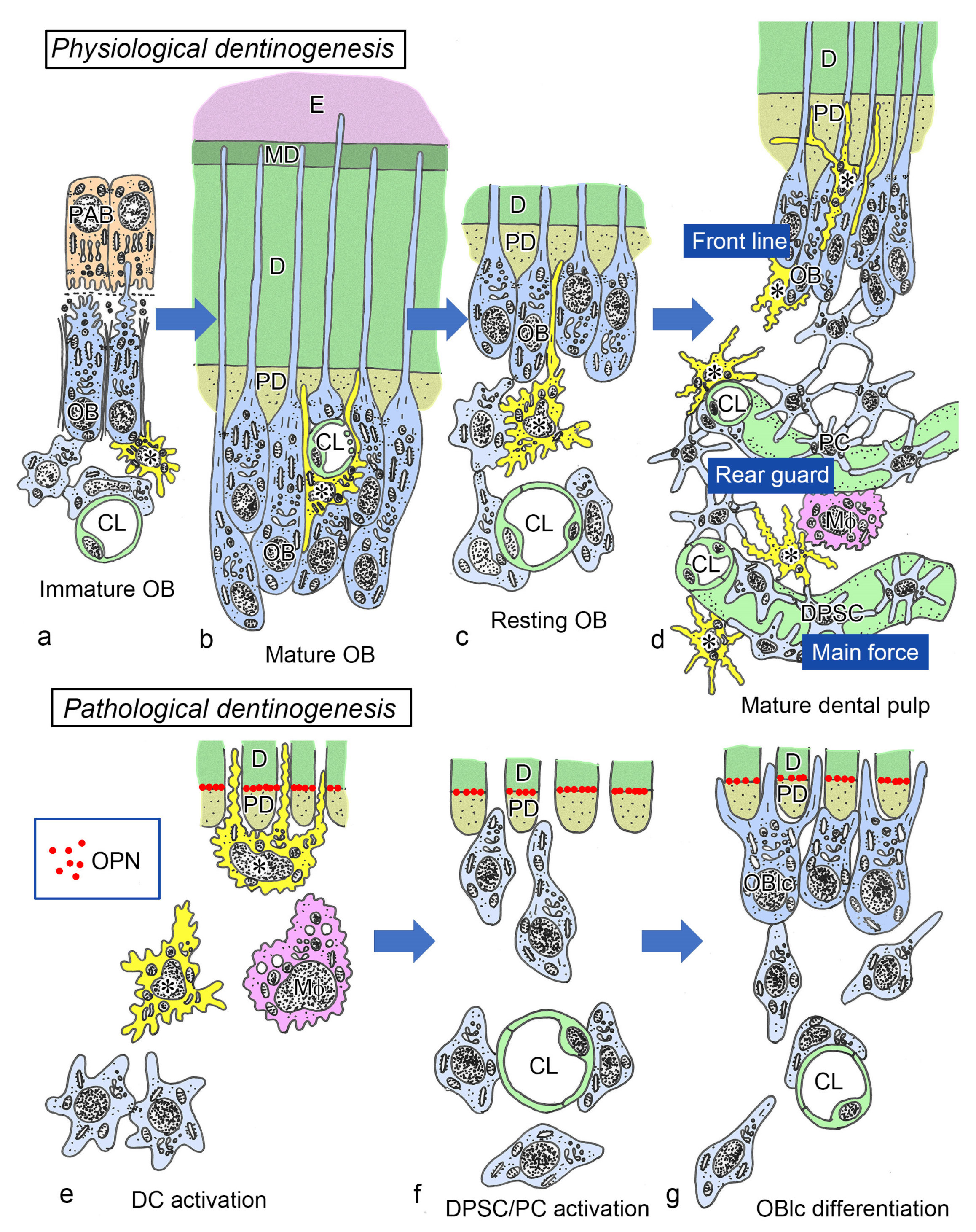{kind=link}
Abstract
:1. Introduction
2. Key Functions of DCs
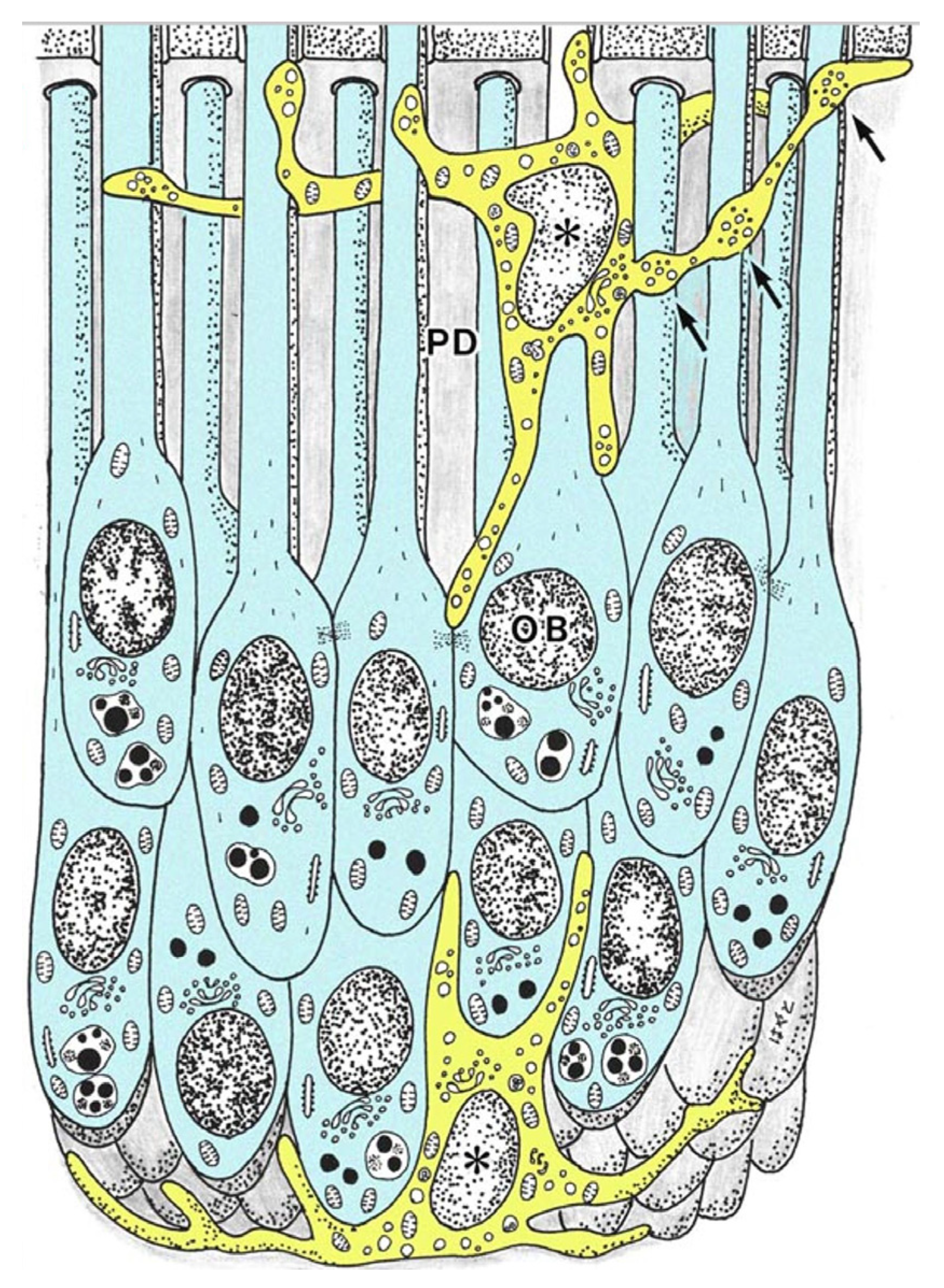
3. Relationship between DCs and Odontoblasts during Physiological Dentinogenesis
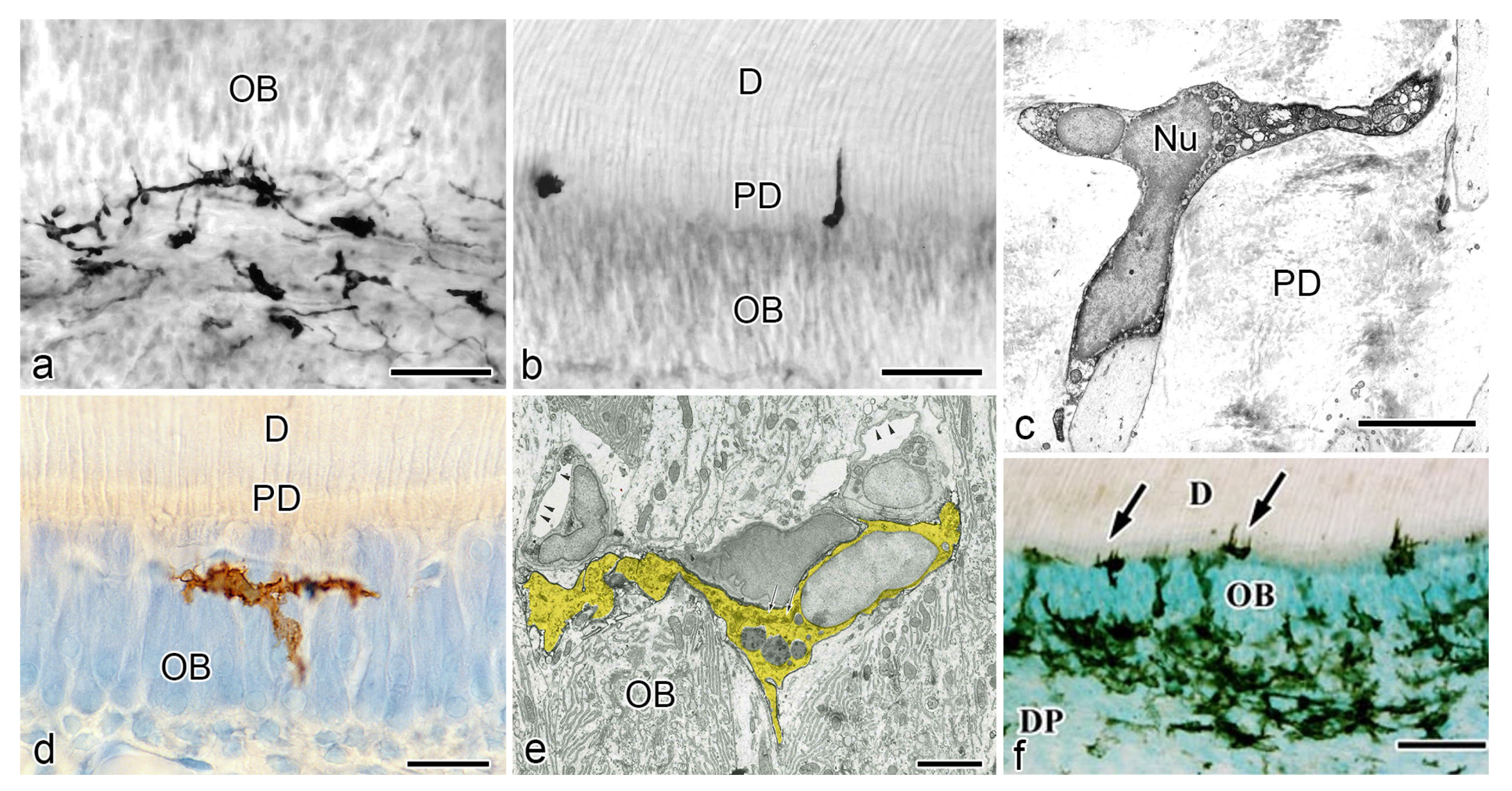
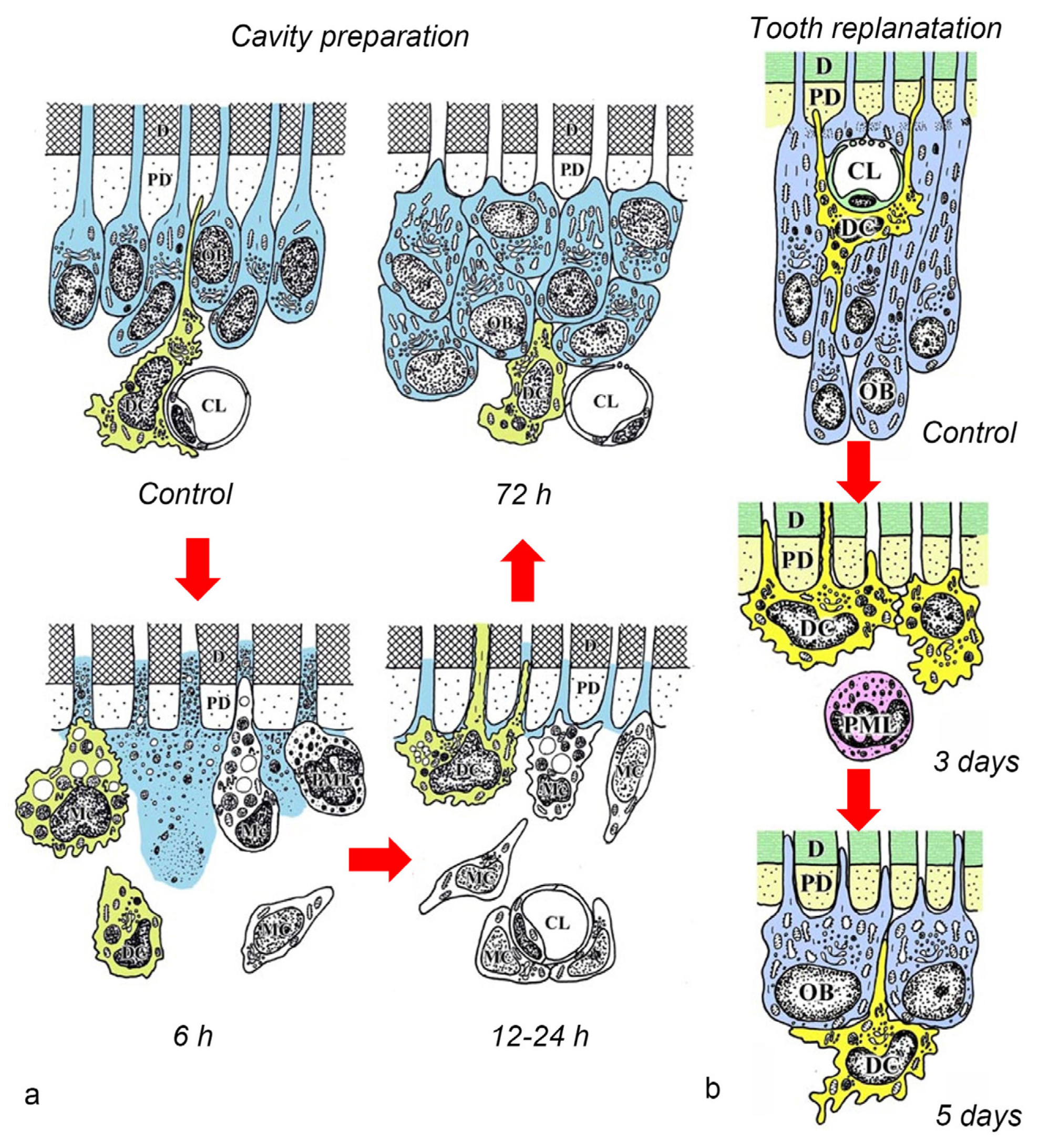
4. Relationship between DCs and Odontoblasts or Newly Differentiated Odontoblast-like Cells after Noninfected Exogenous Injuries
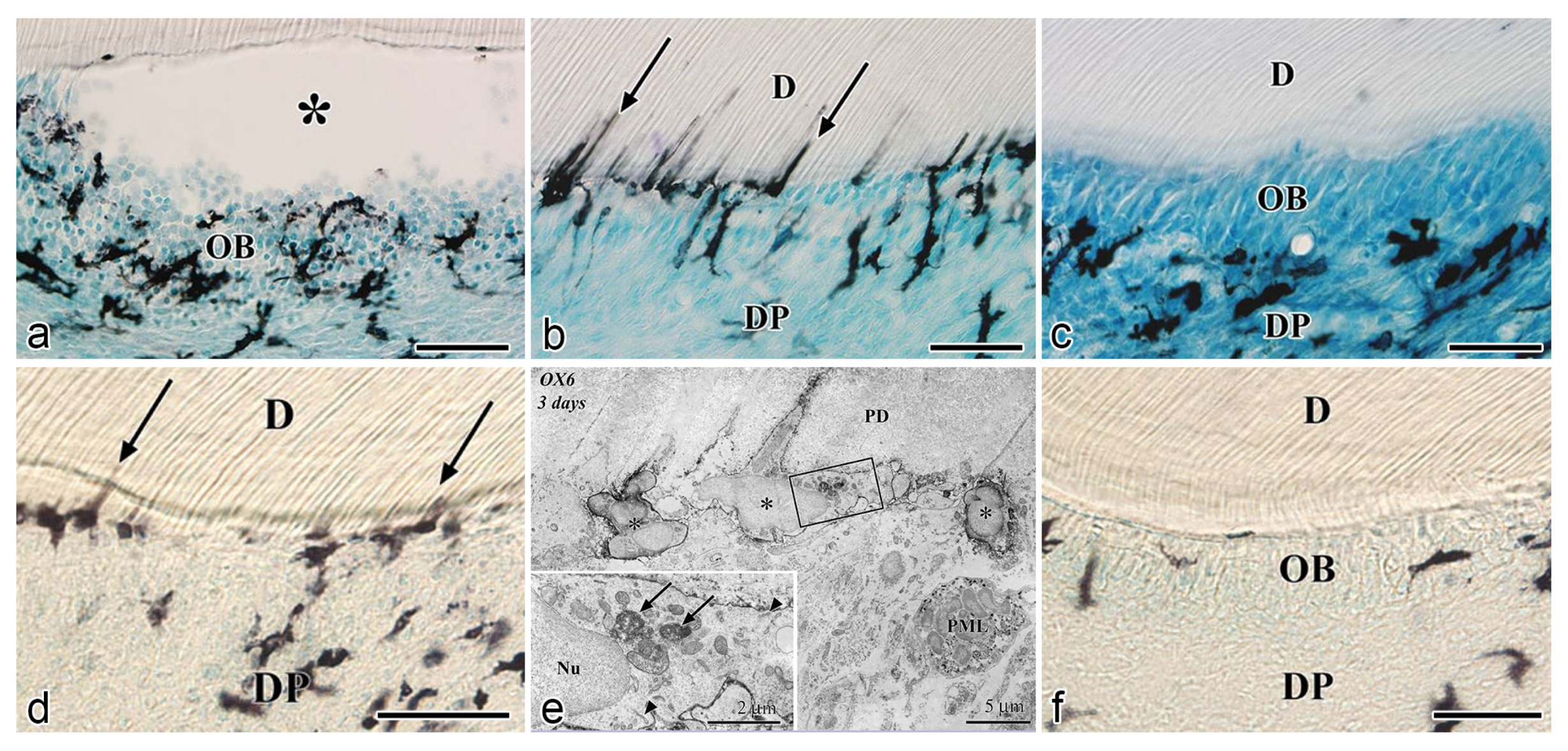
5. Responses of DCs to Bacterial Infection in the Dental Pulp
6. Relationship between DCs and Osteopontin (OPN) during Pathological Dentinogenesis
7. Relationship between DCs and Stem/Progenitor Cells in the Dental Pulp after Injuries
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nanci, A. Ten Cate’s Oral Histology: Development, Structure, and Function, 9th ed.; Elsevier: St. Louis, MO, USA, 2018; p. 344. [Google Scholar]
- Berkovitz, B.K.B.; Holland, G.R.; Moxham, B.J.; Berkovitz, B.K.B. Oral Anatomy, Histology and Embryology, 5th ed.; Elsevier: Edinburgh, UK, 2018; pp. 1–462. [Google Scholar]
- Chiego, D.J. Essentials of Oral Histology and Embryology: A Clinical Approach, 5th ed.; Elsevier: St. Louis, MO, USA, 2018. [Google Scholar]
- Rajan, S.; Ljunggren, A.; Manton, D.J.; Bjorkner, A.E.; McCullough, M. Post-mitotic odontoblasts in health, disease, and regeneration. Arch. Oral. Biol. 2020, 109, 104591. [Google Scholar] [CrossRef]
- Yumoto, H.; Hirao, K.; Hosokawa, Y.; Kuramoto, H.; Takegawa, D.; Nakanishi, T.; Matsuo, T. The roles of odontoblasts in dental pulp innate immunity. Jpn. Dent. Sci. Rev. 2018, 54, 105–117. [Google Scholar] [CrossRef]
- Couve, E.; Osorio, R.; Schmachtenberg, O. The amazing odontoblast: Activity, autophagy, and aging. J. Dent. Res. 2013, 92, 765–772. [Google Scholar] [CrossRef]
- Hargreaves, K.M.; Goodis, H.E.; Tay, F.R. Seltzer and Bender’s Dental Pulp, 2nd ed.; Quintessence Publishing: Hanover Park, IL, USA, 2012; p. 501. [Google Scholar]
- Orchardson, R.; Cadden, S.W. An update on the physiology of the dentine-pulp complex. Dent. Update 2001, 28, 200–209. [Google Scholar] [CrossRef]
- Mjor, I.A.; Sveen, O.B.; Heyeraas, K.J. Pulp-dentin biology in restorative dentistry. Part 1: Normal structure and physiology. Quintessence Int. 2001, 32, 427–446. [Google Scholar]
- Casasco, A.; Casasco, M.; Calligaro, A.; Ferrieri, G.; Brambilla, E.; Strohmenger, L.; Alberici, R.; Mazzini, G. Cell proliferation in developing human dental pulp. A combined flow cytometric and immunohistochemical study. Eur. J. Oral Sci. 1997, 105, 609–613. [Google Scholar] [CrossRef]
- Piattelli, A.; Rubini, C.; Fioroni, M.; Ciavarelli, L.; De Fazio, P. bcl-2, p53, and MIB-1 in human adult dental pulp. J. Endod. 2000, 26, 225–227. [Google Scholar] [CrossRef]
- Vermelin, L.; Lecolle, S.; Septier, D.; Lasfargues, J.J.; Goldberg, M. Apoptosis in human and rat dental pulp. Eur. J. Oral Sci. 1996, 104, 547–553. [Google Scholar] [CrossRef]
- Franquin, J.C.; Remusat, M.; Abou Hashieh, I.; Dejou, J. Immunocytochemical detection of apoptosis in human odontoblasts. Eur. J. Oral Sci. 1998, 106 (Suppl. 1), 384–387. [Google Scholar] [CrossRef]
- Nishikawa, S.; Sasaki, F. Apoptosis of dental pulp cells and their elimination by macrophages and MHC class II-expressing dendritic cells. J. Histochem. Cytochem. 1999, 47, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Satchell, P.G.; Gutmann, J.L.; Witherspoon, D.E. Apoptosis: An introduction for the endodontist. Int. Endod. J. 2003, 36, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Kenmotsu, S.; Nakasone, N.; Nakakura-Ohshima, K.; Ohshima, H. Cell dynamics in the pulpal healing process following cavity preparation in rat molars. Histochem. Cell Biol. 2008, 130, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Heyeraas, K.J.; Sveen, O.B.; Mjor, I.A. Pulp-dentin biology in restorative dentistry. Part 3: Pulpal inflammation and its sequelae. Quintessence Int. 2001, 32, 611–625. [Google Scholar] [PubMed]
- Bjorndal, L.; Mjor, I.A. Pulp-dentin biology in restorative dentistry. Part 4: Dental caries--characteristics of lesions and pulpal reactions. Quintessence Int. 2001, 32, 717–736. [Google Scholar]
- Mjor, I.A.; Odont, D. Pulp-dentin biology in restorative dentistry. Part 2: Initial reactions to preparation of teeth for restorative procedures. Quintessence Int. 2001, 32, 537–551. [Google Scholar]
- Farges, J.C.; Keller, J.F.; Carrouel, F.; Durand, S.H.; Romeas, A.; Bleicher, F.; Lebecque, S.; Staquet, M.J. Odontoblasts in the dental pulp immune response. J. Exp. Zool. B Mol. Dev. Evol. 2009, 312B, 425–436. [Google Scholar] [CrossRef]
- Farges, J.C.; Alliot-Licht, B.; Baudouin, C.; Msika, P.; Bleicher, F.; Carrouel, F. Odontoblast control of dental pulp inflammation triggered by cariogenic bacteria. Front. Physiol. 2013, 4, 326. [Google Scholar] [CrossRef] [Green Version]
- Farges, J.C.; Alliot-Licht, B.; Renard, E.; Ducret, M.; Gaudin, A.; Smith, A.J.; Cooper, P.R. Dental pulp defence and repair mechanisms in dental caries. Mediat. Inflamm. 2015, 2015, 230251. [Google Scholar] [CrossRef] [Green Version]
- Tziafas, D. Mechanisms controlling secondary initiation of dentinogenesis: A review. Int. Endod. J. 1994, 27, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Ruch, J.V.; Lesot, H.; Begue-Kirn, C. Odontoblast differentiation. Int. J. Dev. Biol. 1995, 39, 51–68. [Google Scholar]
- Smith, A.J.; Cassidy, N.; Perry, H.; Begue-Kirn, C.; Ruch, J.V.; Lesot, H. Reactionary dentinogenesis. Int. J. Dev. Biol. 1995, 39, 273–280. [Google Scholar]
- Magloire, H.; Romeas, A.; Melin, M.; Couble, M.L.; Bleicher, F.; Farges, J.C. Molecular regulation of odontoblast activity under dentin injury. Adv. Dent. Res. 2001, 15, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Lesot, H. Induction and regulation of crown dentinogenesis: Embryonic events as a template for dental tissue repair? Crit. Rev. Oral Biol. Med. 2001, 12, 425–437. [Google Scholar] [CrossRef]
- Tjaderhane, L. The mechanism of pulpal wound healing. Aust. Endod. J. 2002, 28, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Arana-Chavez, V.E.; Massa, L.F. Odontoblasts: The cells forming and maintaining dentine. Int. J. Biochem. Cell. Biol. 2004, 36, 1367–1373. [Google Scholar] [CrossRef]
- Saito, K.; Nakatomi, M.; Ida-Yonemochi, H.; Ohshima, H. Osteopontin is essential for type I collagen secretion in reparative dentin. J. Dent. Res. 2016, 95, 1034–1041. [Google Scholar] [CrossRef]
- Angelova Volponi, A.; Zaugg, L.K.; Neves, V.; Liu, Y.; Sharpe, P.T. Tooth repair and regeneration. Curr. Oral Health Rep. 2018, 5, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Neves, V.C.M.; Sharpe, P.T. Regulation of reactionary dentine formation. J. Dent. Res. 2018, 97, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Tziafas, D. Characterization of odontoblast-like cell phenotype and reparative dentin formation in vivo: A comprehensive literature review. J. Endod. 2019, 45, 241–249. [Google Scholar] [CrossRef]
- Dammaschke, T.; Stratmann, U.; Fischer, R.J.; Sagheri, D.; Schafer, E. Proliferation of rat molar pulp cells after direct pulp capping with dentine adhesive and calcium hydroxide. Clin. Oral Investig. 2011, 15, 577–587. [Google Scholar] [CrossRef]
- Quispe-Salcedo, A.; Ida-Yonemochi, H.; Ohshima, H. Use of a triple antibiotic solution affects the healing process of intentionally delayed replanted teeth in mice. J. Oral Biosci. 2013, 55, 91–100. [Google Scholar] [CrossRef]
- Quispe-Salcedo, A.; Ida-Yonemochi, H.; Ohshima, H. Effects of a triple antibiotic solution on pulpal dynamics after intentionally delayed tooth replantation in mice. J. Endod. 2014, 40, 1566–1572. [Google Scholar] [CrossRef]
- Quispe-Salcedo, A.; Sato, T.; Matsuyama, J.; Ida-Yonemochi, H.; Ohshima, H. Responses of oral-microflora-exposed dental pulp to capping with a triple antibiotic paste or calcium hydroxide cement in mouse molars. Regen. Ther. 2020, 15, 216–225. [Google Scholar] [CrossRef]
- Saito, K.; Nakatomi, M.; Ohshima, H. Dynamics of bromodeoxyuridine label-retaining dental pulp cells during pulpal healing after cavity preparation in mice. J. Endod. 2013, 39, 1250–1255. [Google Scholar] [CrossRef]
- Ohshima, H.; Kawahara, I.; Maeda, T.; Takano, Y. The relationship between odontoblasts and immunocompetent cells during dentinogenesis in rat incisors: An immunohistochemical study using OX6-monoclonal antibody. Arch. Histol. Cytol. 1994, 57, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, H.; Sato, O.; Kawahara, I.; Maeda, T.; Takano, Y. Responses of immunocompetent cells to cavity preparation in rat molars: An immunohistochemical study using OX6-monoclonal antibody. Connect. Tissue Res. 1995, 32, 303–311. [Google Scholar] [CrossRef]
- Kannari, N.; Ohshima, H.; Maeda, T.; Noda, T.; Takano, Y. Class II MHC antigen-expressing cells in the pulp tissue of human deciduous teeth prior to shedding. Arch. Histol. Cytol. 1998, 61, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Rungvechvuttivittaya, S.; Okiji, T.; Suda, H. Responses of macrophage-associated antigen-expressing cells in the dental pulp of rat molars to experimental tooth replantation. Arch. Oral Biol. 1998, 43, 701–710. [Google Scholar] [CrossRef]
- Shimizu, A.; Nakakura-Ohshima, K.; Noda, T.; Maeda, T.; Ohshima, H. Responses of immunocompetent cells in the dental pulp to replantation during the regeneration process in rat molars. Cell Tissue Res. 2000, 302, 221–233. [Google Scholar] [CrossRef]
- Ohshima, H.; Nakakura-Ohshima, K.; Takeuchi, K.; Hoshino, M.; Takano, Y.; Maeda, T. Pulpal regeneration after cavity preparation, with special reference to close spatio-relationships between odontoblasts and immunocompetent cells. Microsc. Res. Tech. 2003, 60, 483–490. [Google Scholar] [CrossRef]
- Nakakura-Ohshima, K.; Watanabe, J.; Kenmotsu, S.; Ohshima, H. Possible role of immunocompetent cells and the expression of heat shock protein-25 in the process of pulpal regeneration after tooth injury in rat molars. J. Electron. Microsc. 2003, 52, 581–591. [Google Scholar] [CrossRef]
- Suzuki, T.; Nomura, S.; Maeda, T.; Ohshima, H. An immunocytochemical study of pulpal responses to cavity preparation by laser ablation in rat molars by using antibodies to heat shock protein (Hsp) 25 and class II MHC antigen. Cell Tissue Res. 2004, 315, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, E.; Nakakura-Ohshima, K.; Nomura, S.; Ohshima, H. Pulpal responses to cavity preparation in aged rat molars. Cell Tissue Res. 2006, 326, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kenmotsu, S.; Nakakura-Ohshima, K.; Takahashi, N.; Ohshima, H. Responses of infected dental pulp to α TCP-containing antimicrobials in rat molars. Arch. Histol. Cytol. 2011, 73, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Nakatomi, M.; Ida-Yonemochi, H.; Kenmotsu, S.; Ohshima, H. The expression of GM-CSF and osteopontin in immunocompetent cells precedes the odontoblast differentiation following allogenic tooth transplantation in mice. J. Histochem. Cytochem. 2011, 59, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshima, H.; Maeda, T.; Takano, Y. Acid phosphatase activity in the class II MHC antigen-expressing cells in the rat incisor pulp. Dent. Jpn. 1997, 33, 8–14. [Google Scholar]
- Ohshima, H.; Takano, Y.; Sato, O.; Kawahara, I.; Maeda, T. Responses of class II MHC antigen-expressing cells to cavity preparation. In Dentin/Pulp Complex, Proceedings of the International Conference on Dentin Pulp Complex, Chiba, Japan, 1-4 July 1995; Quintessence Pub. Co.: Tokyo, Japan; Chicago, IL, USA, 1996; pp. 316–318. [Google Scholar]
- Lhuillier, C.; Galluzzi, L. Preface—Dendritic cells: Master regulators of innate and adaptive immunity. Int. Rev. Cell. Mol. Biol. 2019, 348, ix–xiv. [Google Scholar] [CrossRef]
- Devi, K.S.; Anandasabapathy, N. The origin of DCs and capacity for immunologic tolerance in central and peripheral tissues. Semin. Immunopathol. 2017, 39, 137–152. [Google Scholar] [CrossRef]
- Tsukamoto-Tanaka, H.; Ikegame, M.; Takagi, R.; Harada, H.; Ohshima, H. Histochemical and immunocytochemical study of hard tissue formation in dental pulp during the healing process in rat molars after tooth replantation. Cell Tissue Res. 2006, 325, 219–229. [Google Scholar] [CrossRef]
- Ogawa, R.; Saito, C.; Jung, H.S.; Ohshima, H. Capacity of dental pulp differentiation after tooth transplantation. Cell Tissue Res. 2006, 326, 715–724. [Google Scholar] [CrossRef]
- Takamori, Y.; Suzuki, H.; Nakakura-Ohshima, K.; Cai, J.; Cho, S.W.; Jung, H.S.; Ohshima, H. Capacity of dental pulp differentiation in mouse molars as demonstrated by allogenic tooth transplantation. J. Histochem. Cytochem. 2008, 56, 1075–1086. [Google Scholar] [CrossRef]
- Unno, H.; Suzuki, H.; Nakakura-Ohshima, K.; Jung, H.S.; Ohshima, H. Pulpal regeneration following allogenic tooth transplantation into mouse maxilla. Anat. Rec. 2009, 292, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Mutoh, N.; Nakatomi, M.; Ida-Yonemochi, H.; Nakagawa, E.; Tani-Ishii, N.; Ohshima, H. Responses of BrdU label-retaining dental pulp cells to allogenic tooth transplantation into mouse maxilla. Histochem. Cell Biol. 2011, 136, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Quispe-Salcedo, A.; Ida-Yonemochi, H.; Ohshima, H. The effects of enzymatically synthesized glycogen on the pulpal healing process of extracted teeth following intentionally delayed replantation in mice. J. Oral Biosci. 2015, 57, 124–130. [Google Scholar] [CrossRef]
- Kuratate, M.; Yoshiba, K.; Shigetani, Y.; Yoshiba, N.; Ohshima, H.; Okiji, T. Immunohistochemical analysis of nestin, osteopontin, and proliferating cells in the reparative process of exposed dental pulp capped with mineral trioxide aggregate. J. Endod. 2008, 34, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Shigetani, Y.; Suzuki, H.; Ohshima, H.; Yoshiba, K.; Yoshiba, N.; Okiji, T. Odontoblast response to cavity preparation with Er:YAG laser in rat molars: An immunohistochemical study. Odontology 2013, 101, 186–192. [Google Scholar] [CrossRef]
- Nakakura-Ohshima, K.; Quispe-Salcedo, A.; Sano, H.; Hayasaki, H.; Ohshima, H. The effects of reducing the root length by apicoectomy on dental pulp revascularization following tooth replantation in mice. Dent. Traumatol. 2021. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Ida-Yonemochi, H.; Nakakura-Ohshima, K.; Ohshima, H. The relationship between cell proliferation and differentiation and mapping of putative dental pulp stem/progenitor cells during mouse molar development by chasing BrdU-labeling. Cell Tissue Res. 2012, 348, 95–107. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Ida-Yonemochi, H.; Suzuki, H.; Nakakura-Ohshima, K.; Jung, H.S.; Honda, M.J.; Ishii, Y.; Watanabe, N.; Ohshima, H. Mapping of BrdU label-retaining dental pulp cells in growing teeth and their regenerative capacity after injuries. Histochem. Cell Biol. 2010, 134, 227–241. [Google Scholar] [CrossRef]
- Saito, K.; Ishikawa, Y.; Nakakura-Ohshima, K.; Ida-Yonemochi, H.; Nakatomi, M.; Kenmotsu, S.; Ohshima, H. Differentiation capacity of BrdU label-retaining dental pulp cells during pulpal healing following allogenic transplantation in mice. Biomed. Res. 2011, 32, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammaschke, T. Rat molar teeth as a study model for direct pulp capping research in dentistry. Lab. Anim. 2010, 44, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Andreasen, J.O.; Andersson, L. Critical considerations when planning experimental in vivo studies in dental traumatology. Dent. Traumatol. 2011, 27, 275–280. [Google Scholar] [CrossRef]
- Kim, S.; Shin, S.J.; Song, Y.; Kim, E. In vivo experiments with dental pulp stem cells for pulp-dentin complex regeneration. Mediat. Inflamm. 2015, 2015, 409347. [Google Scholar] [CrossRef] [Green Version]
- Balan, S.; Saxena, M.; Bhardwaj, N. Dendritic cell subsets and locations. Int. Rev. Cell. Mol. Biol. 2019, 348, 1–68. [Google Scholar] [CrossRef]
- Steinman, R.M.; Cohn, Z.A. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp. Med. 1973, 137, 1142–1162. [Google Scholar] [CrossRef]
- Patel, V.I.; Metcalf, J.P. Identification and characterization of human dendritic cell subsets in the steady state: A review of our current knowledge. J. Investig. Med. 2016, 64, 833–847. [Google Scholar] [CrossRef]
- Nudel, I.; Elnekave, M.; Furmanov, K.; Arizon, M.; Clausen, B.E.; Wilensky, A.; Hovav, A.H. Dendritic cells in distinct oral mucosal tissues engage different mechanisms to prime CD8+ T cells. J. Immunol. 2011, 186, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Geginat, J.; Nizzoli, G.; Paroni, M.; Maglie, S.; Larghi, P.; Pascolo, S.; Abrignani, S. Immunity to pathogens taught by specialized human dendritic cell subsets. Front. Immunol. 2015, 6, 527. [Google Scholar] [CrossRef]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Constantino, J.; Gomes, C.; Falcao, A.; Neves, B.M.; Cruz, M.T. Dendritic cell-based immunotherapy: A basic review and recent advances. Immunol. Res. 2017, 65, 798–810. [Google Scholar] [CrossRef]
- Cabeza-Cabrerizo, M.; Cardoso, A.; Minutti, C.M.; Pereira da Costa, M.; Reis, E.S.C. Dendritic cells revisited. Annu. Rev. Immunol. 2021, 39, 131–166. [Google Scholar] [CrossRef]
- Keselowsky, B.G.; Lewis, J.S. Dendritic cells in the host response to implanted materials. Semin. Immunol. 2017, 29, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Van Brussel, I.; Berneman, Z.N.; Cools, N. Optimizing dendritic cell-based immunotherapy: Tackling the complexity of different arms of the immune system. Mediat. Inflamm. 2012, 2012, 690643. [Google Scholar] [CrossRef]
- Jontell, M.; Gunraj, M.N.; Bergenholtz, G. Immunocompetent cells in the normal dental pulp. J. Dent. Res. 1987, 66, 1149–1153. [Google Scholar] [CrossRef]
- Okiji, T.; Jontell, M.; Belichenko, P.; Bergenholtz, G.; Dahlstrom, A. Perivascular dendritic cells of the human dental pulp. Acta Physiol. Scand. 1997, 159, 163–169. [Google Scholar] [CrossRef]
- Yoshiba, N.; Yoshiba, K.; Nakamura, H.; Iwaku, M.; Ozawa, H. Immunohistochemical localization of HLA-DR-positive cells in unerupted and erupted normal and carious human teeth. J. Dent. Res. 1996, 75, 1585–1589. [Google Scholar] [CrossRef]
- Ohshima, H.; Maeda, T.; Takano, Y. The distribution and ultrastructure of class II MHC-positive cells in human dental pulp. Cell Tissue Res. 1999, 295, 151–158. [Google Scholar] [CrossRef]
- Sotirovska Ivkovska, A.; Zabokova-Bilbilova, E.; Georgiev, Z.; Bajraktarova Valjakova, E.; Ivkovski, L. Immunohistochemical study on antigen-presenting cells in healthy and carious human teeth. Bratisl. Lek. Listy 2018, 119, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudin, A.; Renard, E.; Hill, M.; Bouchet-Delbos, L.; Bienvenu-Louvet, G.; Farges, J.C.; Cuturi, M.C.; Alliot-Licht, B. Phenotypic analysis of immunocompetent cells in healthy human dental pulp. J. Endod. 2015, 41, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Nakasone, N.; Hashimoto, E.; Sakai, H.; Nakakura-Ohshima, K.; Harada, H. The eternal tooth germ is formed at the apical end of continuously growing teeth. Arch. Oral. Biol. 2005, 50, 153–157. [Google Scholar] [CrossRef]
- Kawashima, N.; Okiji, T.; Kosaka, T.; Suda, H. Kinetics of macrophages and lymphoid cells during the development of experimentally induced periapical lesions in rat molars: A quantitative immunohistochemical study. J. Endod. 1996, 22, 311–316. [Google Scholar] [CrossRef]
- Zhang, J.; Kawashima, N.; Suda, H.; Nakano, Y.; Takano, Y.; Azuma, M. The existence of CD11c+ sentinel and F4/80+ interstitial dendritic cells in dental pulp and their dynamics and functional properties. Int. Immunol. 2006, 18, 1375–1384. [Google Scholar] [CrossRef] [Green Version]
- Jontell, M.; Bergenholtz, G.; Scheynius, A.; Ambrose, W. Dendritic cells and macrophages expressing class II antigens in the normal rat incisor pulp. J. Dent. Res. 1988, 67, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Okiji, T.; Kawashima, N.; Kosaka, T.; Matsumoto, A.; Kobayashi, C.; Suda, H. An immunohistochemical study of the distribution of immunocompetent cells, especially macrophages and Ia antigen-expressing cells of heterogeneous populations, in normal rat molar pulp. J. Dent. Res. 1992, 71, 1196–1202. [Google Scholar] [CrossRef]
- Tsuruga, E.; Sakakura, Y.; Yajima, T.; Shide, N. Appearance and distribution of dendritic cells and macrophages in dental pulp during early postnatal morphogenesis of mouse mandibular first molars. Histochem. Cell Biol. 1999, 112, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Bergenholtz, G.; Nagaoka, S.; Jontell, M. Class II antigen expressing cells in experimentally induced pulpitis. Int. Endod. J. 1991, 24, 8–14. [Google Scholar] [CrossRef]
- Fristad, I.; Heyeraas, K.J.; Kvinnsland, I.H.; Jonsson, R. Recruitment of immunocompetent cells after dentinal injuries in innervated and denervated young rat molars: An immunohistochemical study. J. Histochem. Cytochem. 1995, 43, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, C.; Kimura, K.; Nakayama, T.; Toyoshima, K.; Terashita, M. Primary and secondary induction of apoptosis in odontoblasts after cavity preparation of rat molars. J. Dent. Res. 2001, 80, 1530–1534. [Google Scholar] [CrossRef]
- Ohshima, H.; Nakakura-Ohshima, K.; Maeda, T. Expression of heat-shock protein 25 immunoreactivity in the dental pulp and enamel organ during odontogenesis in the rat molar. Connect. Tissue Res. 2002, 43, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Inoue, H.; Matsuura, H.; Mukae, F.; Ishikawa, H.; Hirano, H.; Tamura, N. Age-related changes in the immunoreactivity of the monocyte/macrophage system in rat molar pulp after cavity preparation. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2002, 94, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.L.; Liewehr, F.R. Update on the adaptive immune responses of the dental pulp. J. Endod. 2007, 33, 773–781. [Google Scholar] [CrossRef]
- Shah, D.; Lynd, T.; Ho, D.; Chen, J.; Vines, J.; Jung, H.D.; Kim, J.H.; Zhang, P.; Wu, H.; Jun, H.W.; et al. Pulp-dentin tissue healing response: A discussion of current biomedical approaches. J. Clin. Med. 2020, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Kamal, A.M.; Okiji, T.; Kawashima, N.; Suda, H. Defense responses of dentin/pulp complex to experimentally induced caries in rat molars: An immunohistochemical study on kinetics of pulpal Ia antigen-expressing cells and macrophages. J. Endod. 1997, 23, 115–120. [Google Scholar] [CrossRef]
- Izumi, T.; Kobayashi, I.; Okamura, K.; Matsuo, K.; Kiyoshima, T.; Ishibashi, Y.; Inoue, H.; Sakai, H. An immunohistochemical study of HLA-DR and alpha 1-antichymotrypsin-positive cells in the pulp of human non-carious and carious teeth. Arch. Oral Biol. 1996, 41, 627–630. [Google Scholar] [CrossRef]
- Yoshiba, N.; Yoshiba, K.; Iwaku, M.; Ozawa, H. Immunohistochemical localizations of class II antigens and nerve fibers in human carious teeth: HLA-DR immunoreactivity in Schwann cells. Arch. Histol. Cytol. 1998, 61, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, K.; Okiji, T.; Suda, H. Co-increase of nerve fibers and HLA-DR- and/or factor-XIIIa-expressing dendritic cells in dentinal caries-affected regions of the human dental pulp: An immunohistochemical study. J. Dent. Res. 1999, 78, 1596–1608. [Google Scholar] [CrossRef] [PubMed]
- Yoshiba, K.; Yoshiba, N.; Iwaku, M. Class II antigen-presenting dendritic cell and nerve fiber responses to cavities, caries, or caries treatment in human teeth. J. Dent. Res. 2003, 82, 422–427. [Google Scholar] [CrossRef]
- Tanabe, K.; Yoshiba, K.; Yoshiba, N.; Iwaku, M.; Ozawa, H. Immunohistochemical study on pulpal response in rat molars after cavity preparation by Er:YAG laser. Eur. J. Oral Sci. 2002, 110, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, A.; Scutera, S.; Sozzani, S.; Musso, T. Role of osteopontin in dendritic cell shaping of immune responses. Cytokine Growth Factor Rev. 2019, 50, 19–28. [Google Scholar] [CrossRef]
- Weber, G.F. The phylogeny of osteopontin-analysis of the protein sequence. Int. J. Mol. Sci. 2018, 19, 2557. [Google Scholar] [CrossRef] [Green Version]
- Butler, W.T. The nature and significance of osteopontin. Connect. Tissue Res. 1989, 23, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.F.; Berse, B.; Van de Water, L.; Papadopoulos-Sergiou, A.; Perruzzi, C.A.; Manseau, E.J.; Dvorak, H.F.; Senger, D.R. Expression and distribution of osteopontin in human tissues: Widespread association with luminal epithelial surfaces. Mol. Biol. Cell 1992, 3, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.T.; Ritchie, H. The nature and functional significance of dentin extracellular matrix proteins. Int. J. Dev. Biol. 1995, 39, 169–179. [Google Scholar] [PubMed]
- Kawamura, K.; Iyonaga, K.; Ichiyasu, H.; Nagano, J.; Suga, M.; Sasaki, Y. Differentiation, maturation, and survival of dendritic cells by osteopontin regulation. Clin. Diagn. Lab. Immunol. 2005, 12, 206–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scutera, S.; Salvi, V.; Lorenzi, L.; Piersigilli, G.; Lonardi, S.; Alotto, D.; Casarin, S.; Castagnoli, C.; Dander, E.; D’Amico, G.; et al. Adaptive regulation of osteopontin production by dendritic cells through the bidirectional interaction with mesenchymal stromal cells. Front. Immunol. 2018, 9, 1207. [Google Scholar] [CrossRef]
- Inoue, M.; Shinohara, M.L. Intracellular osteopontin (iOPN) and immunity. Immunol. Res. 2011, 49, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittling, S.R.; Singh, R. Osteopontin in immune-mediated diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef] [Green Version]
- Rittling, S.R. Osteopontin in macrophage function. Expert Rev. Mol. Med. 2011, 13, e15. [Google Scholar] [CrossRef]
- Matsuo, K.; Ray, N. Osteoclasts, mononuclear phagocytes, and c-Fos: New insight into osteoimmunology. Keio J. Med. 2004, 53, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Blengio, F.; Raggi, F.; Pierobon, D.; Cappello, P.; Eva, A.; Giovarelli, M.; Varesio, L.; Bosco, M.C. The hypoxic environment reprograms the cytokine/chemokine expression profile of human mature dendritic cells. Immunobiology 2013, 218, 76–89. [Google Scholar] [CrossRef]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Miner. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.W.; Feng, J.; Daltoe, F.; Fatscher, R.; Gentleman, E.; Gentleman, M.M.; Sharpe, P.T. Perivascular stem cells at the tip of mouse incisors regulate tissue regeneration. J. Bone Miner. Res. 2016, 31, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Vidovic-Zdrilic, I.; Vijaykumar, A.; Mina, M. Activation of alphaSMA expressing perivascular cells during reactionary dentinogenesis. Int. Endod. J. 2019, 52, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Ethokic, J.M.; Tomic, S.Z.; Colic, M.J. Cross-talk between mesenchymal stem/stromal cells and dendritic cells. Curr. Stem Cell Res. Ther. 2016, 11, 51–65. [Google Scholar] [PubMed]
- Zhang, W.; Dang, K.; Huai, Y.; Qian, A. Osteoimmunology: The regulatory roles of T lymphocytes in osteoporosis. Front. Endocrinol. 2020, 11, 465. [Google Scholar] [CrossRef]
- Negrescu, A.M.; Cimpean, A. The state of the art and prospects for osteoimmunomodulatory biomaterials. Materials 2021, 14, 1357. [Google Scholar] [CrossRef]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The conceptual framework unifying the immune and skeletal systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef]
- Okamoto, K.; Takayanagi, H. Osteoimmunology. Cold Spring Harb. Perspect. Med. 2019, 9. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhu, Q.L.; Huang, P.; Yu, Q.; Wang, Z.H.; Cooper, P.R.; Smith, A.J.; He, W. CpG ODN-induced matrix metalloproteinase-13 expression is mediated via activation of the ERK and NF-kappaB signalling pathways in odontoblast cells. Int. Endod. J. 2013, 46, 666–674. [Google Scholar] [CrossRef]
- Hoshi, K.; Yamazaki, T.; Sugiyama, Y.; Tsukakoshi, K.; Tsugawa, W.; Sode, K.; Ikebukuro, K. G-quadruplex structure improves the immunostimulatory effects of CpG oligonucleotides. Nucleic Acid Ther. 2019, 29, 224–229. [Google Scholar] [CrossRef]
- Hanagata, N. CpG oligodeoxynucleotide nanomedicines for the prophylaxis or treatment of cancers, infectious diseases, and allergies. Int. J. Nanomed. 2017, 12, 515–531. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quispe-Salcedo, A.; Ohshima, H. The Role of Dendritic Cells during Physiological and Pathological Dentinogenesis. J. Clin. Med. 2021, 10, 3348. https://doi.org/10.3390/jcm10153348
Quispe-Salcedo A, Ohshima H. The Role of Dendritic Cells during Physiological and Pathological Dentinogenesis. Journal of Clinical Medicine. 2021; 10(15):3348. https://doi.org/10.3390/jcm10153348
Chicago/Turabian StyleQuispe-Salcedo, Angela, and Hayato Ohshima. 2021. "The Role of Dendritic Cells during Physiological and Pathological Dentinogenesis" Journal of Clinical Medicine 10, no. 15: 3348. https://doi.org/10.3390/jcm10153348