
The Interplay between TRPM7 and MagT1 in Maintaining Endothelial Magnesium Homeostasis
 , , , , , and
, , , , , and 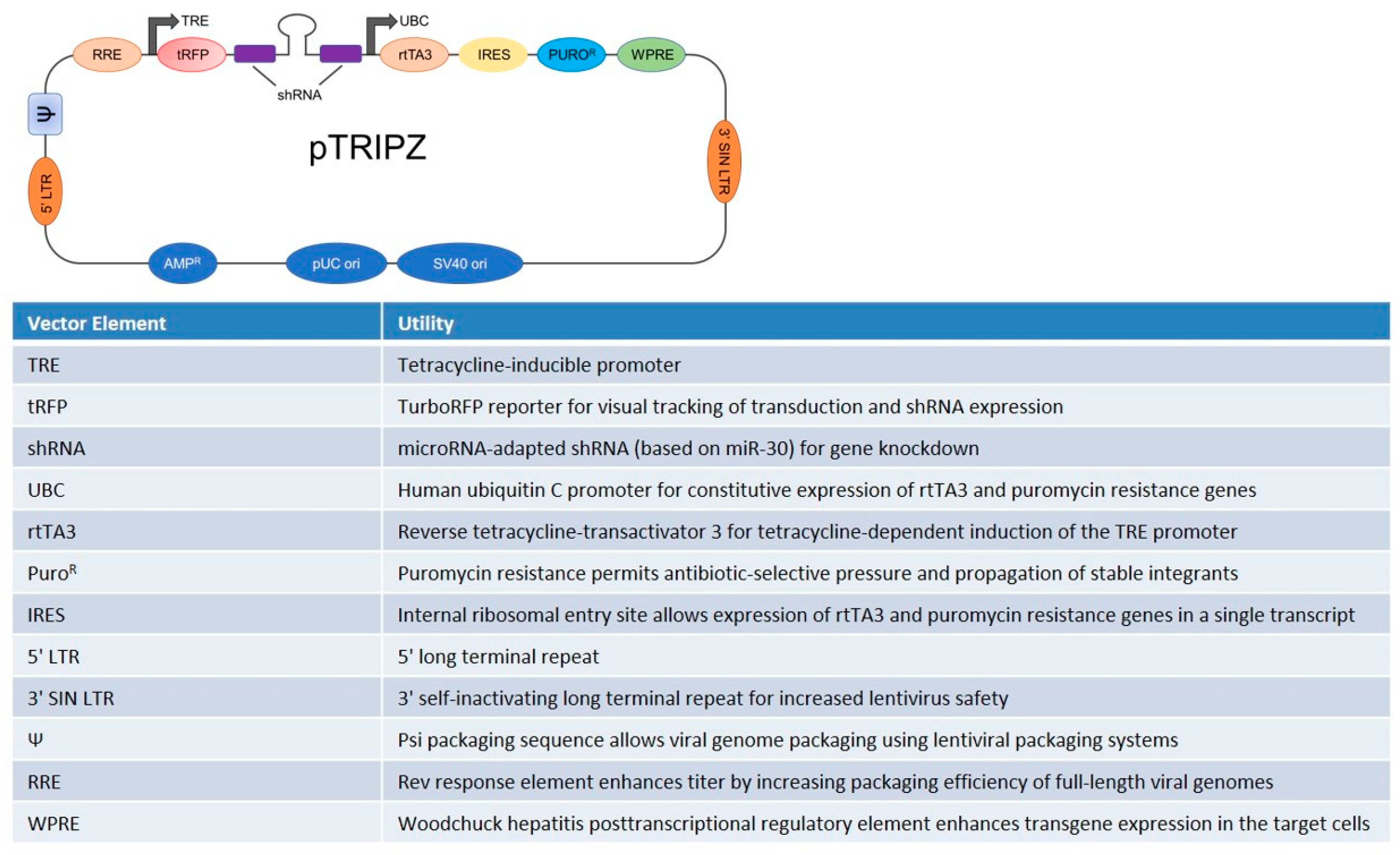{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Generation of Stably Transfected HUVEC with pTRIPZ Lentiviral Vector Containing TRPM7 shRNA
2.3. Cell Proliferation and Reactive Oxygen Species Production
2.4. Western Blot
2.5. Quantification of Total Intracellular Mg Concentration
2.6. Real-Time PCR
2.7. Confocal Microscopy
2.8. Atomic Force Microscopy Measurements
2.9. X-ray Fluorescence Microscopy and Scanning Transmission Microscopy Measurements
2.10. Elemental Quantification
2.11. Statistical Analysis
3. Results
3.1. Generation of TRPM7-Silencing HUVEC
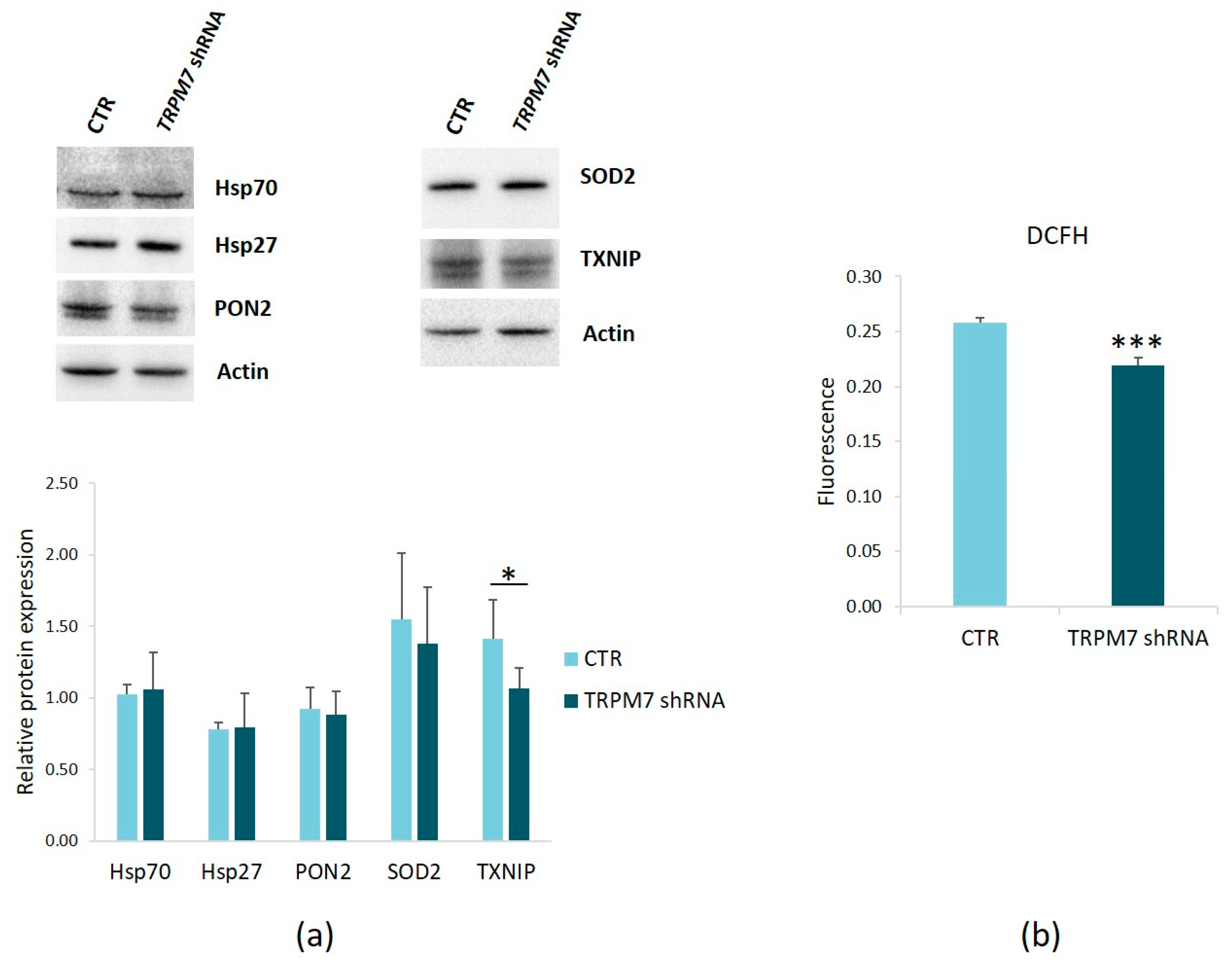
3.2. TRPM7 Silencing Decreases ROS Production in HUVEC
3.3. TRPM7 Silencing Increases HUVEC Proliferation
3.4. TRPM7 Silencing Does Not Modulate Total Mg
3.5. TRPM7 Silencing Modulates the Levels of MagT1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rooney, M.R.; Alonso, A.; Folsom, A.R.; Michos, E.D.; Rebholz, C.M.; Misialek, J.R.; Chen, L.Y.; Dudley, S.; Lutsey, P.L. Serum magnesium and the incidence of coronary artery disease over a median 27 years of follow-up in the Atherosclerosis Risk in Communities (ARIC) Study and a meta-analysis. Am. J. Clin. Nutr. 2020, 111, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, A.; Naghshi, S.; Sadeghi, O.; Larijani, B.; Esmaillzadeh, A. Total, Dietary, and Supplemental Magnesium Intakes and Risk of All-Cause, Cardiovascular, and Cancer Mortality: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Adv. Nutr. 2021, 12, 1196–1210. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, L.; Fedele, G.; Castiglioni, S.; Maier, J.A. Magnesium Deficiency Induces Lipid Accumulation in Vascular Endothelial Cells via Oxidative Stress-The Potential Contribution of EDF-1 and PPARγ. Int. J. Mol. Sci. 2021, 22, 1050. [Google Scholar] [CrossRef] [PubMed]
- Ozen, M.; Xie, H.; Shin, N.; Al Yousif, G.; Clemens, J.; McLane, M.W.; Lei, J.; Burd, I. Magnesium sulfate inhibits inflammation through P2X7 receptors in human umbilical vein endothelial cells. Pediatr. Res. 2020, 87, 463–471. [Google Scholar] [CrossRef]
- Baldoli, E.; Maier, J.A.M. Silencing TRPM7 mimics the effects of magnesium deficiency in human microvascular endothelial cells. Angiogenesis 2012, 15, 47–57. [Google Scholar] [CrossRef]
- Baldoli, E.; Castiglioni, S.; Maier, J.A.M. Regulation and Function of TRPM7 in Human Endothelial Cells: TRPM7 as a Potential Novel Regulator of Endothelial Function. PLoS ONE 2013, 8, e59891. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Inoue, K.; Sun, H.; Leng, T.; Feng, X.; Zhu, L.; Xiong, Z.G. TRPM7 regulates vascular endothelial cell adhesion and tube formation. Am. J. Physiol.-Cell Physiol. 2015, 308, C308–C318. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.-Y.; Li, S.-F.; Chen, Y.; Zhao, Y.-J.; Hu, W.; Lu, C.; Zhou, R.-P. Transient receptor potential melastatin 7 and their modulators. Eur. J. Pharmacol. 2022, 931, 175180. [Google Scholar] [CrossRef]
- Krapivinsky, G.; Krapivinsky, L.; Manasian, Y.; Clapham, D.E. The TRPM7 chanzyme is cleaved to release a chromatin-modifying kinase. Cell 2014, 157, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Scrimieri, R.; Cazzaniga, A.; Castiglioni, S.; Maier, J.A.M. Vitamin d prevents high glucose-induced lipid droplets accumulation in cultured endothelial cells: The role of thioredoxin interacting protein. Biomedicines 2021, 9, 1874. [Google Scholar] [CrossRef]
- Paravicini, T.M.; Yogi, A.; Mazur, A.; Touyz, R.M. Dysregulation of vascular TRPM7 and annexin-1 is associated with endothelial dysfunction in inherited hypomagnesemia. Hypertension 2009, 53, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scrimieri, R.; Locatelli, L.; Cazzola, R.; Maier, J.A.M.; Cazzaniga, A. Reactive oxygen species are implicated in altering magnesium homeostasis in endothelial cells exposed to high glucose. Magnes. Res. 2019, 32, 54–62. [Google Scholar] [PubMed]
- Matsuda-Lennikov, M.; Biancalana, M.; Zou, J.; Ravell, J.C.; Zheng, L.; Kanellopoulou, C.; Jiang, P.; Notarangelo, G.; Jing, H.; Masutani, E.; et al. Magnesium transporter 1 (MAGT1) deficiency causes selective defects in N-linked glycosylation and expression of immune-response genes. J. Biol. Chem. 2019, 294, 13638–13656. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, R.; Tabata, S.; Shindo, Y.; Hotta, K.; Suzuki, K.; Soga, T.; Oka, K. Mitochondrial Mg2+ homeostasis decides cellular energy metabolism and vulnerability to stress. Sci. Rep. 2016, 6, 30027. [Google Scholar] [PubMed]
- Cazzaniga, A.; Locatelli, L.; Castiglioni, S.; Maier, J.A.M. The dynamic adaptation of primary human endothelial cells to simulated microgravity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 5957–5966. [Google Scholar]
- Castiglioni, S.; Cazzaniga, A.; Trapani, V.; Cappadone, C.; Farruggia, G.; Merolle, L.; Wolf, F.I.; Iotti, S.; Maier, J.A.M. Magnesium homeostasis in colon carcinoma LoVo cells sensitive or resistant to doxorubicin. Sci. Rep. 2015, 5, 16538. [Google Scholar] [CrossRef] [Green Version]
- Malucelli, E.; Procopio, A.; Fratini, M.; Gianoncelli, A.; Notargiacomo, A.; Merolle, L.; Sargenti, A.; Castiglioni, S.; Cappadone, C.; Farruggia, G.; et al. Single cell versus large population analysis: Cell variability in elemental intracellular concentration and distribution. Anal. Bioanal. Chem. 2018, 410, 337–348. [Google Scholar]
- Sargenti, A.; Farruggia, G.; Zaccheroni, N.; Marraccini, C.; Sgarzi, M.; Cappadone, C.; Malucelli, E.; Procopio, A.; Prodi, L.; Lombardo, M.; et al. Synthesis of a highly Mg2+-selective fluorescent probe and its application to quantifying and imaging total intracellular magnesium. Nat. Protoc. 2017, 12, 461–471. [Google Scholar]
- Cazzaniga, A.; Maier, J.A.M.; Castiglioni, S. Impact of simulated microgravity on human bone stem cells: New hints for space medicine. Biochem. Biophys. Res. Commun. 2016, 473, 181–186. [Google Scholar] [CrossRef]
- Malucelli, E.; Iotti, S.; Gianoncelli, A.; Fratini, M.; Merolle, L.; Notargiacomo, A.; Marraccini, C.; Sargenti, A.; Cappadone, C.; Farruggia, G.; et al. Quantitative chemical imaging of the intracellular spatial distribution of fundamental elements and light metals in single cells. Anal. Chem. 2014, 86, 5108–5115. [Google Scholar] [CrossRef]
- Gianoncelli, A.; Kourousias, G.; Merolle, L.; Altissimo, M.; Bianco, A. Current status of the TwinMic beamline at Elettra: A soft X-ray transmission and emission microscopy station. J. Synchrotron Radiat. 2016, 23, 1526–1537. [Google Scholar] [CrossRef] [PubMed]
- Janssens, K.H.A.; Adams, F.C.; Rindby, A. Microscopic X-ray Fluorescence Analysis; Wiley: Chichester, UK, 2000. [Google Scholar]
- von Stockar, U.; Liu, J. Does microbial life always feed on negative entropy? Thermodynamic analysis of microbial growth. Biochim. Biophys. Acta 1999, 1412, 191–211. [Google Scholar] [CrossRef] [Green Version]
- Solé, V.A.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A multiplatform code for the analysis of energy-dispersive X-ray fluorescence spectra. Spectrochim. Acta Part B At. Spectrosc. 2007, 62, 63–68. [Google Scholar] [CrossRef]
- Malucelli, E.; Iotti, S.; Fratini, M.; Marraccini, C.; Notargiacomo, A.; Gianoncelli, A.; Bukreeva, I.; Cedola, A.; Maier, J.; Farruggia, G.; et al. X-ray fluorescence microscopy of light elements in cells: Self-absorption correction by integration of compositional and morphological measurements. J. Phys. Conf. Ser. 2013, 463, 12022. [Google Scholar] [CrossRef]
- Vakifahmetoglu-Norberg, H.; Ouchida, A.T.; Norberg, E. The role of mitochondria in metabolism and cell death. Biochem. Biophys. Res. Commun. 2017, 482, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Xiong, Z.-G. Silencing TRPM7 promotes growth/proliferation and nitric oxide production of vascular endothelial cells via the ERK pathway. Cardiovasc. Res. 2009, 83, 547–557. [Google Scholar] [CrossRef]
- Franken, G.A.C.; Huynen, M.A.; Martínez-Cruz, L.A.; Bindels, R.J.M.; de Baaij, J.H.F. Structural and functional comparison of magnesium transporters throughout evolution. Cell. Mol. Life Sci. 2022, 79, 418. [Google Scholar] [CrossRef]
- Locatelli, L.; Cazzaniga, A.; Fedele, G.; Zocchi, M.; Scrimieri, R.; Moscheni, C.; Castiglioni, S.; Maier, J.A. A comparison of doxorubicin-resistant colon cancer lovo and leukemia hl60 cells: Common features, different underlying mechanisms. Curr. Issues Mol. Biol. 2021, 43, 163–175. [Google Scholar] [CrossRef]
- Zocchi, M.; Locatelli, L.; Zuccotti, G.V.; Mazur, A.; Béchet, D.; Maier, J.A.; Castiglioni, S. Magnesium Homeostasis in Myogenic Differentiation-A Focus on the Regulation of TRPM7, MagT1 and SLC41A1 Transporters. Int. J. Mol. Sci. 2022, 23, 1658. [Google Scholar] [CrossRef]
- Zhang, P.; Li, W.; Liu, Y.; Gao, Y.; Abumaria, N. Neuroprotective Effects of TRPM7 Deletion in Parvalbumin GABAergic vs. Glutamatergic Neurons following Ischemia. Cells 2022, 11, 1178. [Google Scholar] [CrossRef]
- Park, C.S.; Lee, J.Y.; Choi, H.Y.; Yune, T.Y. Suppression of Transient Receptor Potential Melastatin 7 by Carvacrol Protects against Injured Spinal Cord by Inhibiting Blood-Spinal Cord Barrier Disruption. J. Neurotrauma 2022, 39, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liao, Q.; Xu, W.; Zhang, Z.; Yin, M.; Cao, X. MiR-129-5p Protects H9c2 Cardiac Myoblasts from Hypoxia/Reoxygenation Injury by Targeting TRPM7 and Inhibiting NLRP3 Inflammasome Activation. J. Cardiovasc. Pharmacol. 2021, 77, 586–593. [Google Scholar] [PubMed]
- Bai, Z.; Feng, J.; Franken, G.A.C.; Al’Saadi, N.; Cai, N.; Yu, A.S.; Lou, L.; Komiya, Y.; Hoenderop, J.G.J.; de Baaij, J.H.F.; et al. CNNM proteins selectively bind to the TRPM7 channel to stimulate divalent cation entry into cells. PLoS Biol. 2021, 19, e3001496. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglioni, S.; Locatelli, L.; Fedele, G.; Cazzaniga, A.; Malucelli, E.; Iotti, S.; Maier, J.A. The Interplay between TRPM7 and MagT1 in Maintaining Endothelial Magnesium Homeostasis. Membranes 2023, 13, 286. https://doi.org/10.3390/membranes13030286
Castiglioni S, Locatelli L, Fedele G, Cazzaniga A, Malucelli E, Iotti S, Maier JA. The Interplay between TRPM7 and MagT1 in Maintaining Endothelial Magnesium Homeostasis. Membranes. 2023; 13(3):286. https://doi.org/10.3390/membranes13030286
Chicago/Turabian StyleCastiglioni, Sara, Laura Locatelli, Giorgia Fedele, Alessandra Cazzaniga, Emil Malucelli, Stefano Iotti, and Jeanette A. Maier. 2023. "The Interplay between TRPM7 and MagT1 in Maintaining Endothelial Magnesium Homeostasis" Membranes 13, no. 3: 286. https://doi.org/10.3390/membranes13030286