1. Introduction
Because of the environmental benefits of insect-rearing for food and feed applications [
1], there is an increasing need to develop suitable technologies to produce insect meals, fractionate the main ingredients, and use suitable processing technologies to yield high-quality, standardized, functional, and tailored insect fractions that can be incorporated into complex product formulations [
2]. While the consumption of whole or minimally processed insects in western markets is still a matter of controversy (the “ick” factor), efficient processes for the recovery of functional insect-derived fractions such as protein, fat, or chitin are being developed to promote consumer acceptance, and to explore a wide range of potential food and non-food applications, with opportunities to use them in various forms being searched for [
3].
Amongst the insect species that are currently under consideration, black soldier fly (
Hermetia illucens) larvae are widely accepted for feed applications, while dried mealworm (
Tenebrio molitor) has already received the favorable opinion of the European Food Safety Authority (EFSA) for food applications [
4]. The insect protein content varies depending on the insect species, the development stage, the way they are processed (thermal, mechanical), and the protocol used for protein isolation. Black soldier fly (BSF) larvae contain 42% crude protein and 29% fat on average in dry matter [
5], and although the protein content is lower than for other orthoptera species, the advantages of BSF are their survival rate and efficiency of converting organic materials into their own biomass. As for mealworm, it is considered as a good source of nutrients, with a protein content between 41–66%, fat content between 15–50%, fiber between 9–19% (all in dry bases), and high amounts of potassium, calcium, iron, and magnesium [
6].
There is a growing interest in the potential of insect proteins as a source of functional and bioactive peptides. The existing knowledge of animal and plant proteins has guided research on the functional and bioactive potential of insect proteins and their hydrolysates. A review by Nongonierna et al. [
7] presents the potential of insect protein hydrolysates and lists protein extraction and hydrolysis processes to produce bioactive peptides from several insect species, mainly silkworm (
Bombix mori) but also mealworm, lesser mealworm (
Alphitobius diaperinus), banded cricket (
Gryllodes sigillatus), and migratory locust (
Locusta migratoria). Enzymatic hydrolysis has been used to efficiently enhance bioactive properties such as antioxidant, antihypertensive, antidiabetic, and antimicrobial properties [
8,
9,
10,
11,
12,
13]. As for techno-functional properties of various insect powders and their protein extracts, including solubility, water and oil binding capacity, foaming capacity, surface hydrophobicity, gelling properties, and emulsifying ability, they have already shown promising outcomes [
14,
15,
16,
17,
18,
19]. Gkinali et al. have recently reviewed the potential of mealworm ingredients for the food industry [
6]. They present an up-to-date summary of functional properties of mealworm flours, defatted flours, and protein extracts. Specifically for emulsifying properties, they conclude from the available literature, mealworm protein extracts have a similar ability to stabilize oil/water interfaces as commercial whey protein. As for foaming capacity, the results published so far indicated a good ability of defatted mealworm preparations for foam stabilization.
As far as techno-functional properties of BSF, Bußler et al. (2016) [
17] presented one of the first studies on the effect of defatting on water- and oil-binding capacities, showing no significant changes between original and defatted samples. Recently, Mshayisa et al. [
20] have reported the effect of different protein extraction methods on techno-functional properties of BSF extracts. They have compared alkaline extraction with alkaline extraction coupled with isoelectric point precipitation, finding that water-binding capacity (WBC) was the highest for BSF protein extracts obtained with alkaline extraction and isoelectric point precipitation, while the oil-binding capacity (OBC) was higher for the original flours (defatted and non-defatted) than for the protein extracts. Regarding emulsifying and foaming properties, they were enhanced in the protein extracts, regardless of the extraction method applied. Moreover, BSF protein concentrate has successfully been used to stabilize sunflower and lemon oil emulsions, produced using a high-throughput membrane emulsification system [
5]. The authors showed that the distribution of droplet sizes for the emulsions and fluxes of emulsions prepared with BSF protein concentrates were comparable to the ones obtained with whey protein isolate, showcasing the potential of BSF to totally or partially replace whey protein in food formulations.
Regardless of the method used to obtain the protein fraction, there is no literature on fractionation of insect proteins with membrane technologies; therefore, the vast existing knowledge on membrane fractionation for dairy, fish, and plant proteins is highly relevant and the mandatory starting point for their design. Usually, whey proteins fractionation and purification by membrane filtration starts with a microfiltration (MF) or centrifugation for defatting and/or clarification. The permeate from MF can be subsequently fractionated using ultrafiltration (UF) to obtain concentrated fractions with a molecular weight range that will depend on the molecular weight cut-off (MWCO) of the membranes employed. A critical issue for insect protein fractionation by UF is the knowledge about their molecular weight distribution. Protein extracts from defatted BSF flours obtained after alkaline solubilization and precipitation at pH 4 presented two major bands at 14.3 and 80.5 kDa [
17], while protein extracts from defatted mealworm flours isolated through the same process showed a different distribution of the protein fractions, with a major fraction between 13–40 kDa and another fraction between 67–250 kDa [
17].
This work assesses for the first time the use of UF to fractionate mealworm and BSF protein concentrates to enhance interfacial properties such as foaming capacity, foam stability, and emulsifying activity. Based on previous knowledge of mealworm and BSF protein molecular size distribution, fractionation with a commercial regenerated cellulose acetate membrane of 30 kDa is suggested to obtain two protein fractions for each insect protein concentrate. The ability of both fractions to stabilize oil–water and air–water interfaces will be tested using standardized methods. The influence of the initial protein concentration in the flux and protein at the outlet of the filtration cell will be studied based on data from or after UF. Since there is no knowledge available, special attention will be paid to determining if the standard cleaning procedures provided by the manufacturer allow for the restoration of membrane permeability facilitating membrane re-use. Foaming and emulsifying properties of the UF fractions will be compared with those of original insect protein concentrates and those obtained using a classical chemical fractionation method, such as acidic solubilization.
2. Materials and Methods
2.1. Materials
Partially defatted black soldier fly powder (BSF) was kindly provided by Hexafly (Ireland). Mealworm (Tenebrio molitor) powder (TM) was purchased from Kreca (The Netherlands). Sodium hydroxide pellets (NaOH, Chem-Lab NV, Zedelgem, Belgium) and hydrochloric acid (37–38% HCL, J. T. Baker, Schwerte, Germany) were used for both BSF and TM protein extraction. For defatting of TM powder, 2-methyltetrahydorfuran (EMPLURA, Darmstadt, Germany) was used. Phosphate buffer (pH 7) was prepared using sodium phosphate dibasic heptahydrate (HNa2O4P7H2O, ACROS, Barcelona, Spain) and sodium phosphate monobasic monohydrate (H2NaO4PH2O, ACROS, Barcelona, Spain). Acetic buffer (pH 5) was prepared with sodium acetate (Sigma-Aldrich, St. Louis, MO, USA) and acetic acid (96%, Panreac, Barcelona, Spain).
2.2. Defatting of Mealworm (TM) Powder
The defatting of TM powder was carried out as per the method provided by Wang et al. [
21] with a slight modification. In brief, TM powder (50 g) was mixed with the 2-methyltetrahydrofuran at a ratio of 1:5 (
w/
v) and the mixture was stirred at 600 rpm on a magnetic stirrer (AGIMATIC-S 7000242, J.P. SELECTA, Barcelona, Spain) for 1 h. Then, the mixture was allowed to stand and settle until complete phase separation, and the solvent layer was decanted. The remaining precipitated TM powder was mixed again with 2-methyltetrahydrofuran at a ratio of 1:5 (
w/
v) and the separation–extraction process was carried out twice more. At the end of the defatting process, the remaining solvent in the precipitate, after the solvent layer was decanted, was evaporated overnight by air drying in a fume hood. Dried, defatted TM powder was then used for protein extraction.
2.3. Black Soldier Fly (BSF) and Mealworm (TM) Protein Concentrates
The protein extraction process for BSF and mealworm to obtain the protein concentrates was based on that of Wang et al. [
5] with some modifications. The defatted TM or partially defatted BSF powder (30 g) was mixed with 0.25 M NaOH solution at a ratio 1:5 (
w/
v) separately and the mixture was heated to 40 °C for one hour with constant agitation at 400 rpm on a magnetic stirrer (RCT ST, IKA, Königswinter, Germany). The mixture was centrifuged (Meditronic 7000599, J.P. SELECTA, Barcelona, Spain) at 3200×
g for 15 min and the supernatant was separated to continue the protein extraction. In BSF supernatant samples the lipid fraction on the top layer was carefully separated. The pH (Accumet AE150, Fisher Scientific, Singapore) of the supernatant was adjusted to reach a value between 4.0–4.2 by adding 37% hydrochloric acid followed by centrifugation at 2233×
g for 15 min. After centrifugation, the pellets were collected in aluminum petri dishes and were kept at −60 °C. The entire process was repeated twice with the pellets that remained after the first centrifugation step. The pellets from the three centrifugation processes were combined and freeze-dried (LYOQUEST-85 PLUS, Telstar, Barcelona, Spain) for 24 h at 0.2 mbar with the plates heated to 20 °C. Freeze-dried protein concentrates were stored in the fridge until further use.
2.4. Membranes
Ultrafiltration experiments were performed using regenerated cellulose membranes from Millipore (PLTK04310 and lot no. C2EB73191) with a molecular weight cut-off of 30 kDa, filter diameter of 44.5 mm and filtration area of 13.4 cm2. All membranes were conditioned as per the guidelines provided by manufacturer before use. Briefly, the membrane disc was carefully removed from the protective package and placed glossy-side-down in a beaker of Milli-Q water for 20 min. This process was repeated three times in total. The clean water flux was determined and recorded at 5 × 104 Pa and 500 rpm. Then, the conditioned membrane disc was stored in wet conditions at 4 °C until further use.
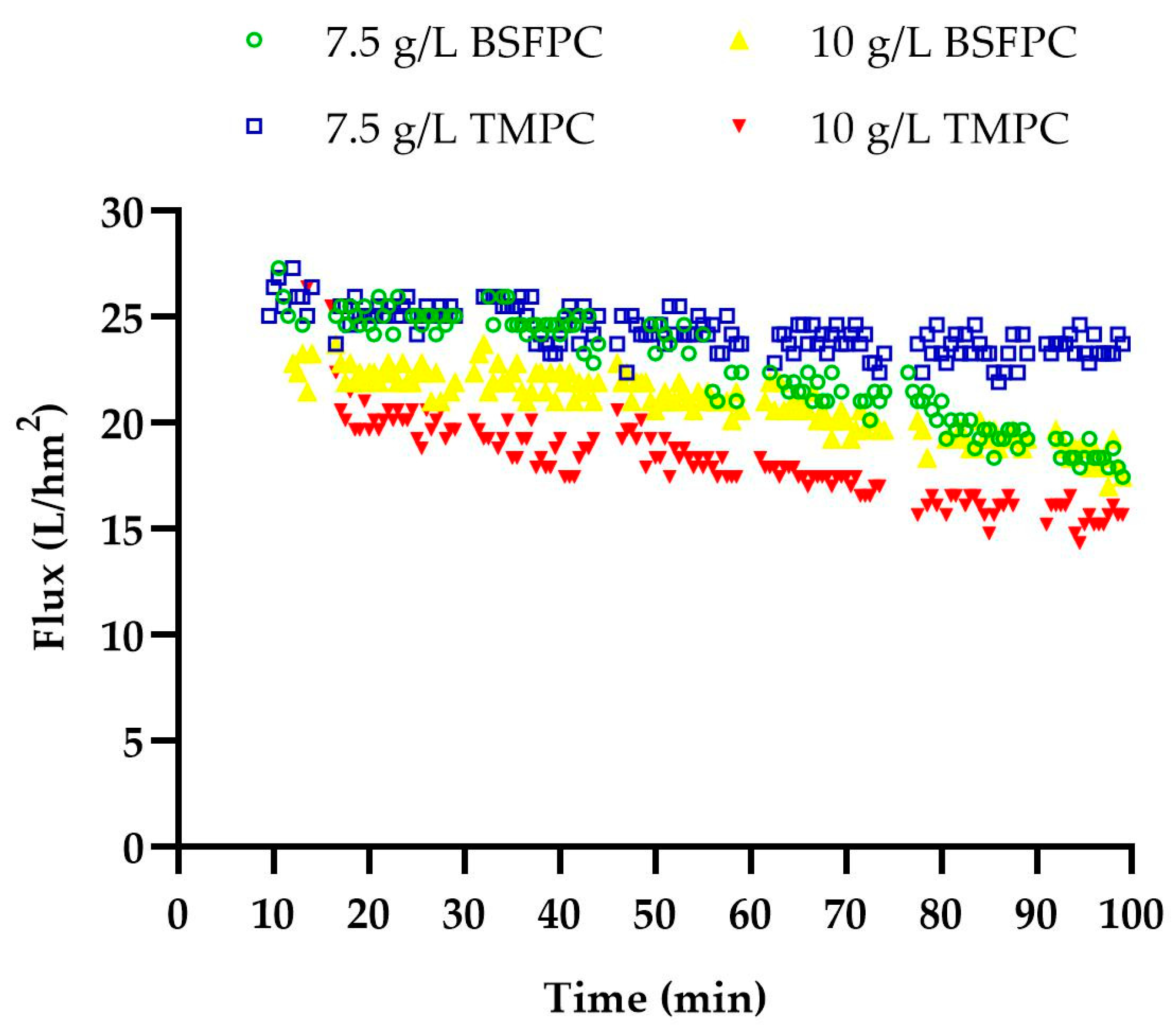
2.5. Ultrafiltration of Black Soldier Fly (BSF) and Mealworm (TM) Protein Concentrates
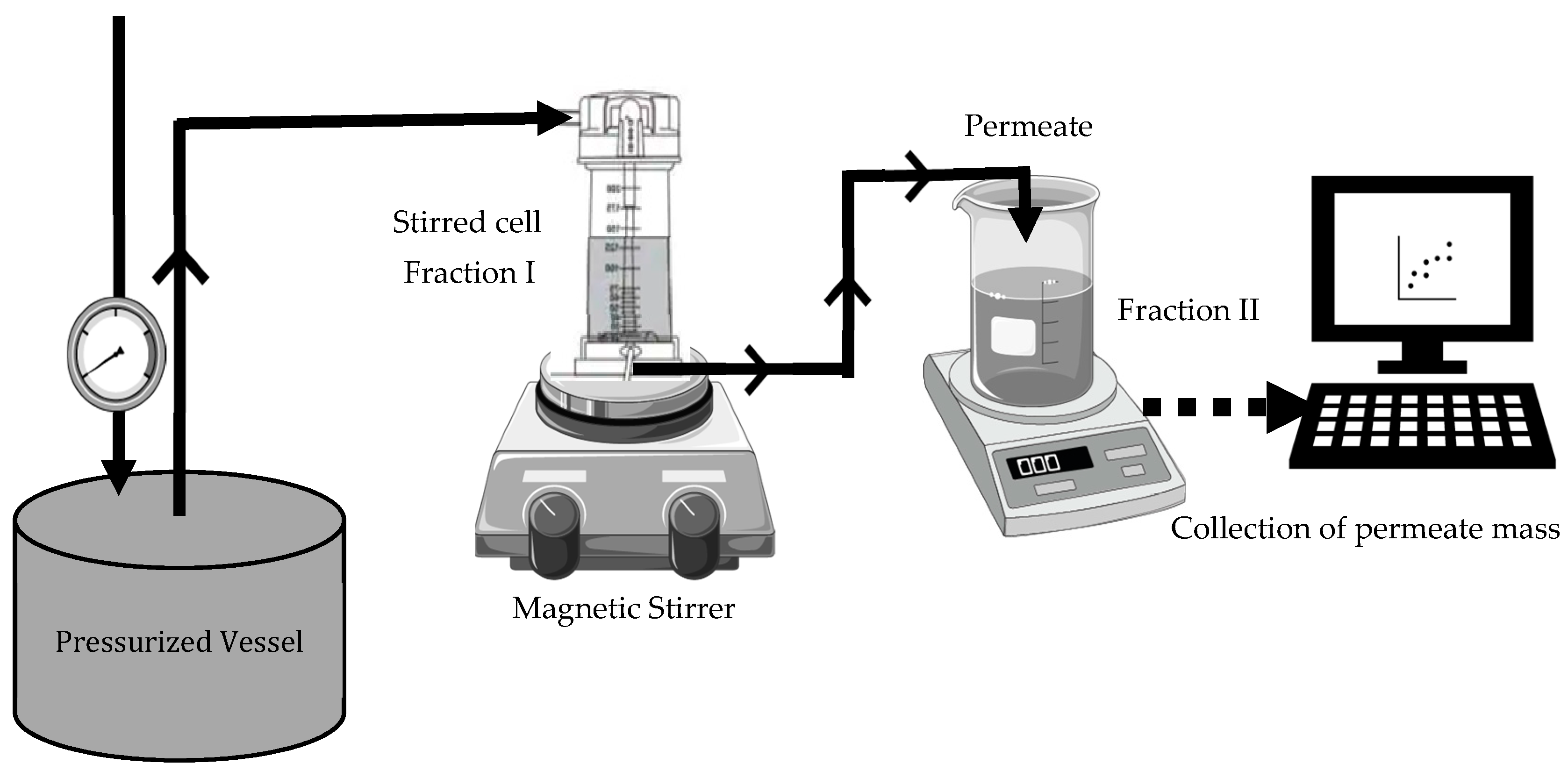
Fractionation of black soldier fly protein concentrate, BSFPC, and mealworm protein concentrate, TMPC, was performed using an experimental apparatus that consisted of a stirred cell (UFSC05001, Amicon 8050, Bedford, MA, USA) connected to a stainless-steel pressurized vessel (Aisi 316L, Advantec MFS, Dublin, CA, USA) containing a protein solution (
Figure 1). All the experiments were carried out at room temperature (20–22 °C), with a stirring speed of 500 rpm and constant transmembrane pressure of 5 × 10
4 Pa.
Freshly prepared 10 or 7.5 g/L protein concentrate solution (BSFPC or TMPC) was loaded into the apparatus. In brief, a protein solution was prepared with 20 g of freeze-dried protein concentrate powder in 400 mL of phosphate buffer (pH 7) and stirred for 2 h while adjusting the pH to 7 with 4 M sodium hydroxide. The solution was then stored at 4 °C overnight. Subsequently, the solution was centrifuged (Meditronic 7000599, J.P. SELECTA, Barcelona, Spain) twice at 2860× g for 15 min. The supernatant was used to prepare 10 or 7.5 g/L protein concentrate solution for the UF run. A new or cleaned 30 kDa membrane was used for each experiment. The permeate was collected in a reservoir placed on an electronic balance (AND FX-3000i, A&D company Ltd, Tokyo, Japan) interfaced to a computer to collect and record time and mass data every 30 s throughout ultrafiltration, which lasted for 90–120 min. Moreover, 500 µL of permeate was collected at 15 min intervals during UF. Each run was performed in duplicate. The fraction which was concentrated inside the stirred cell was collected (referred as Fraction I hereinafter) and the permeate was also collected (referred as Fraction II hereinafter) separately and stored at 4 °C until further use. Further, after each run the membrane was unmounted and cleaned with 0.1 M sodium hydroxide solution for 30 min followed by 100 ppm sodium hypochlorite solution for 30 min. Then, the cleaned membranes were measured for the recovery of clean water flux, and for membranes which were not above the 85% of recovery for clean water flux the cleaning procedure was repeated until the accepted level of flux recovery was reached. The apparatus, without the membrane, was cleaned with 1% sodium hydroxide solution followed by a rinse with distilled water.
Permeate Flux
Permeate flux (L·m
−2·h
−1) was calculated for each ultrafiltration run according to Equation (1).
where Δ
W is the change in mass at the
ith point, Δ
t is the change in time at the
ith point, and A is the filtration area of the membrane. Water density was taken to convert from mass to volume.
2.6. Black Soldier Fly Protein Concentrate and Mealworm Protein Concentrate Isolation by Acidic Solubilization
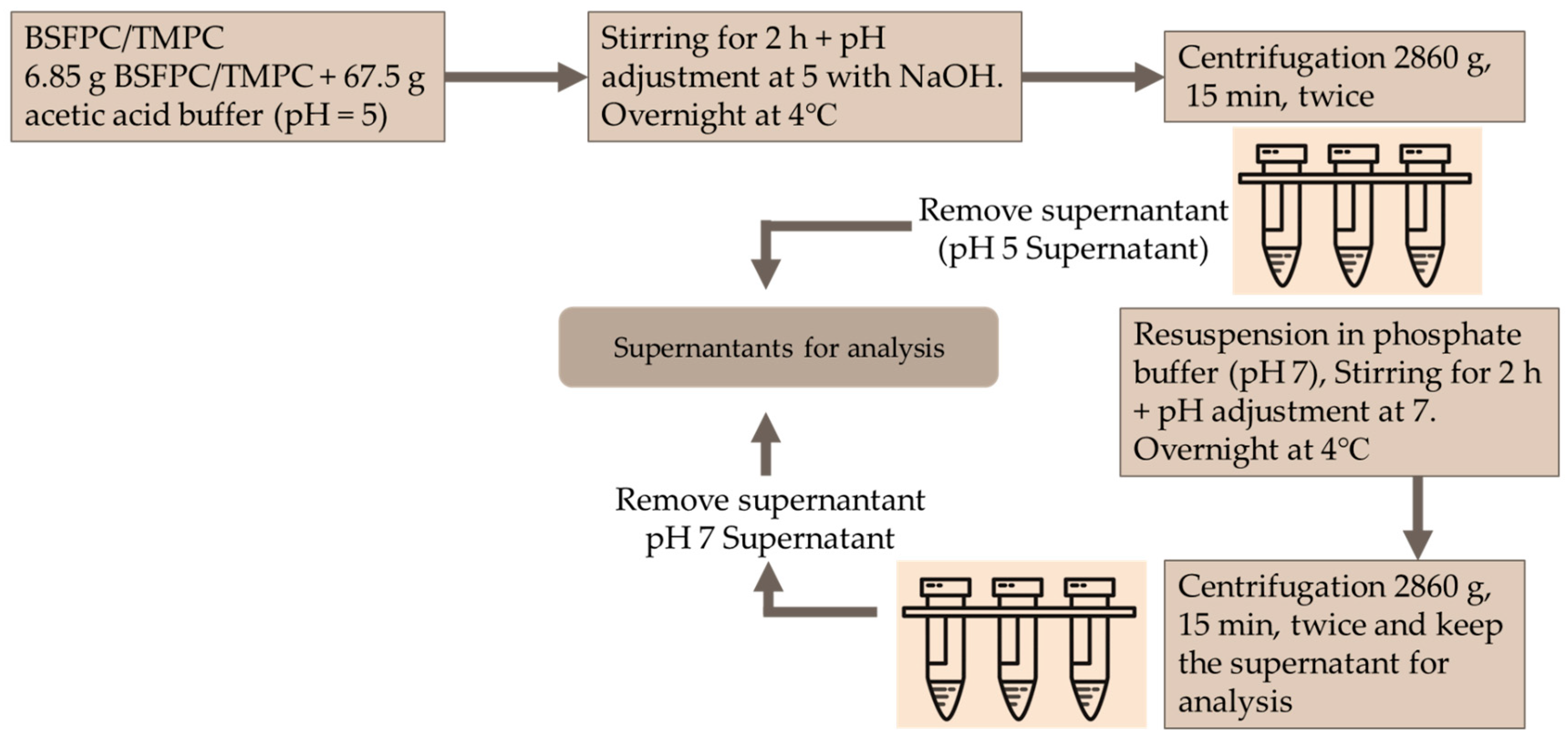
BSFPC and TMPC solutions were fractioned using the pH shifting method (
Figure 2). Briefly, a 10 g/L protein solution was prepared by mixing 6.75 g of BSFPC or TMPC protein powder with 67.5 g of acetic acid buffer (pH 5) and stirred for 2 h while adjusting the pH to 5 with 4 M sodium hydroxide. The mixture was stored at 4 °C for overnight. Subsequently, the mixture was centrifuged (Meditronic 7000599, J.P. SELECTA, Barcelona, Spain) twice at 2860×
g for 15 min. The supernatant was collected and stored at 4 °C for further analysis. The pellet was re-suspended with the same amount of phosphate buffer (pH 7) for 2 h while adjusting pH to 7 with 4 M sodium hydroxide. The resuspended mixture was stored at 4 °C overnight. The supernatant was collected after centrifugation (Meditronic 7000599, J.P. SELECTA, Barcelona, Spain) two times at 2860×
g for 15 min. The collected supernatant was stored at 4 °C for further analysis. As controls, both BSFPC and TMPC solutions were prepared in phosphate buffer (pH 7) as mentioned in
Section 2.4.
2.7. Characterization of the Black Soldier Fly and Mealworm Protein Fractions
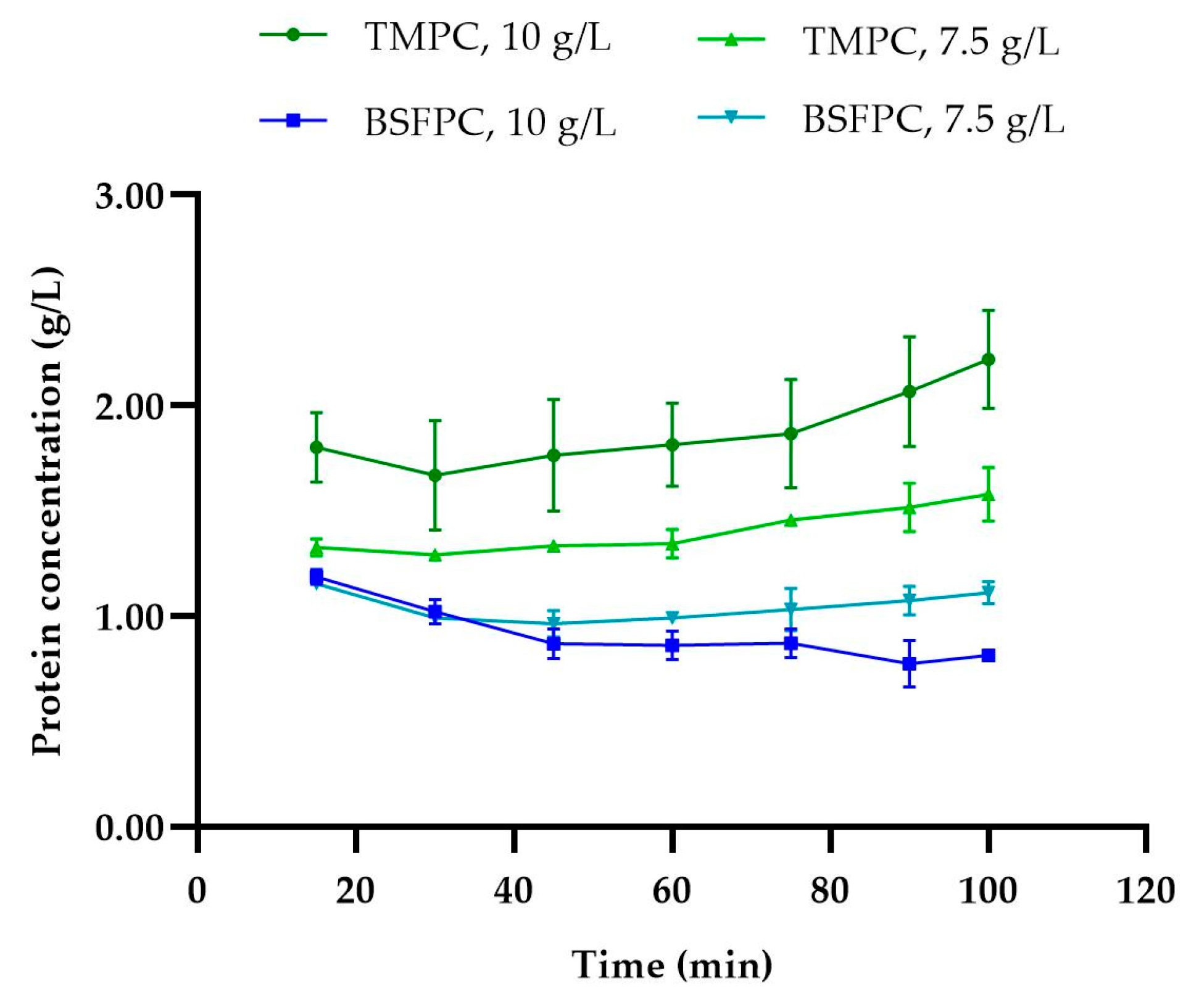
2.7.1. Protein Content
Protein fractions from both ultrafiltration and acidic solubilization processes were analyzed to determine the total soluble protein content using a colorimetric method with a Pierce™ bicinchoninic acid (BCA) protein assay kit (Thermo Scientific, Rockford, IL, USA). BCA assay protein quantification uses bovine serum albumin (BSA) as a standard, and the results are expressed in BSA-equivalent value. To be noted is that black soldier fly and mealworm concentrations that are BSA eq g/L are provided hereinafter as g/L for simplicity.
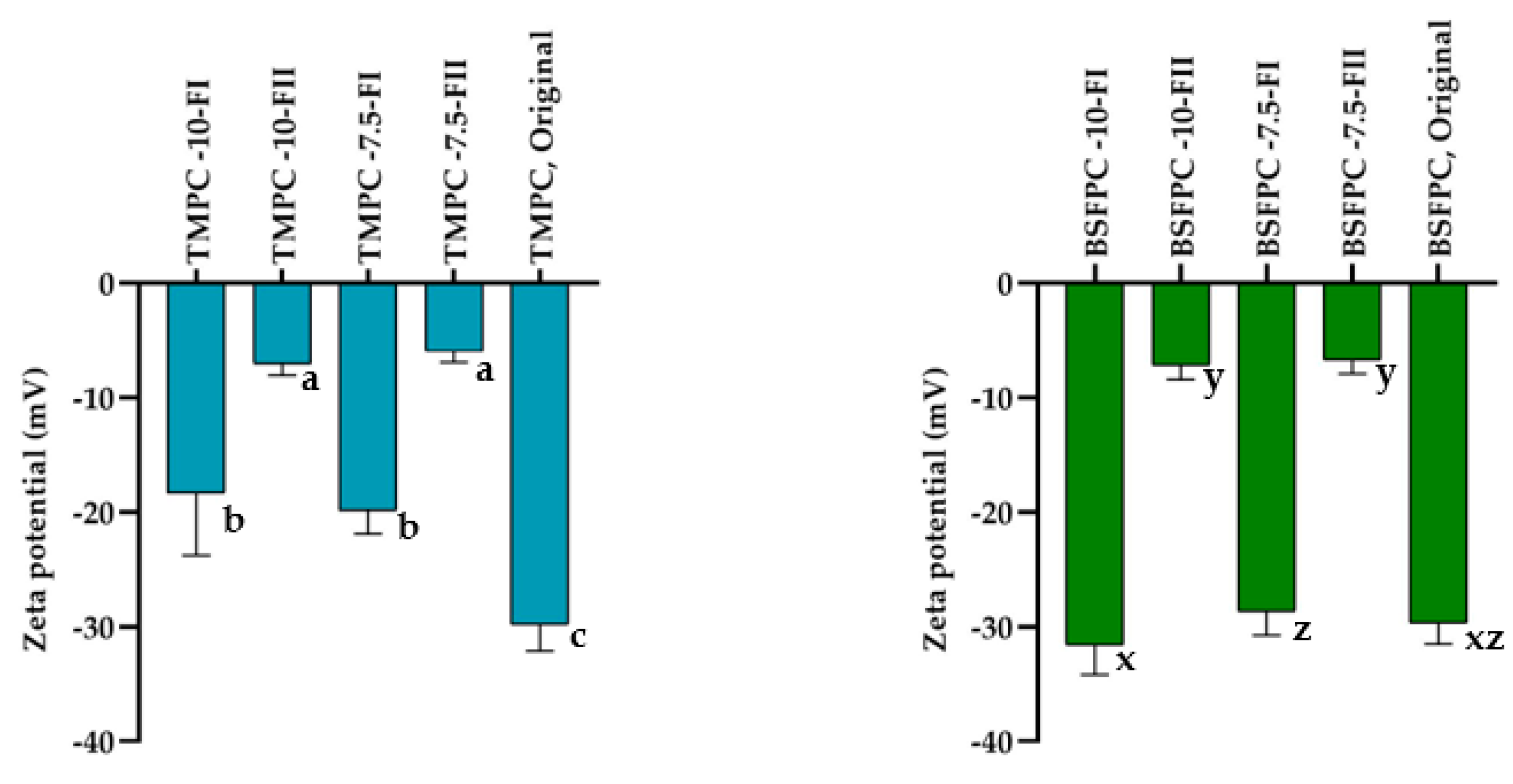
2.7.2. Surface Charge (ζ Potential)
Zeta potential of protein fractions from ultrafiltration was measured using Zetasizer Nano-ZS (Malvern Instruments, Worcestershire, UK). Samples were diluted 10 times by phosphate buffer (pH 7).
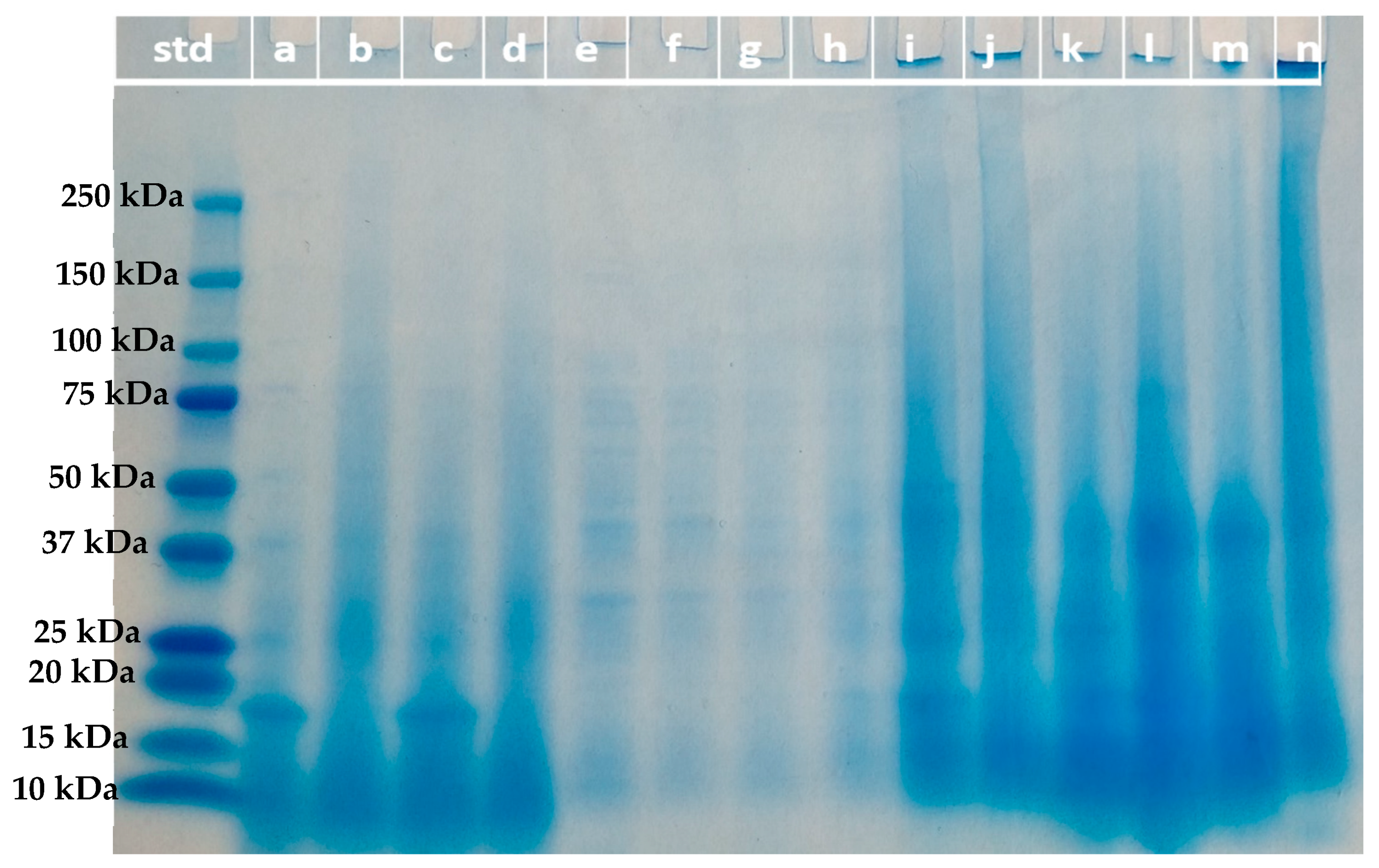
2.7.3. Molecular Weight Distribution of Protein Fractions (SDS-PAGE)
The molecular weight distribution of the protein fractions from ultrafiltration and acidic solubilization was determined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) under reducing conditions [
22]. Samples (10 µL) at a concentration of 2 mg/mL and/or 12 mg/mL were loaded to a 4–20% Mini-PROTEAN
® TGX™ precast gel (Bio-Rad, Hercules, CA, USA). Electrophoresis was run at 80 V for 100 min. Gel was stained with Coomassie blue G250 (Bio-Rad, Hercules, CA, USA) solution for 45 min followed by destaining for 2–3 h. The molecular weight marker (Precision plus protein dual color standard, Bio-Rad, Hercules, CA, USA) with bands varied from 250 kDa to 10 kDa was used as the standard (Bio-Rad, Hercules, CA, USA).
2.8. Techno-Functional Properties of the Black Soldier Fly and Mealworm Protein Fractions
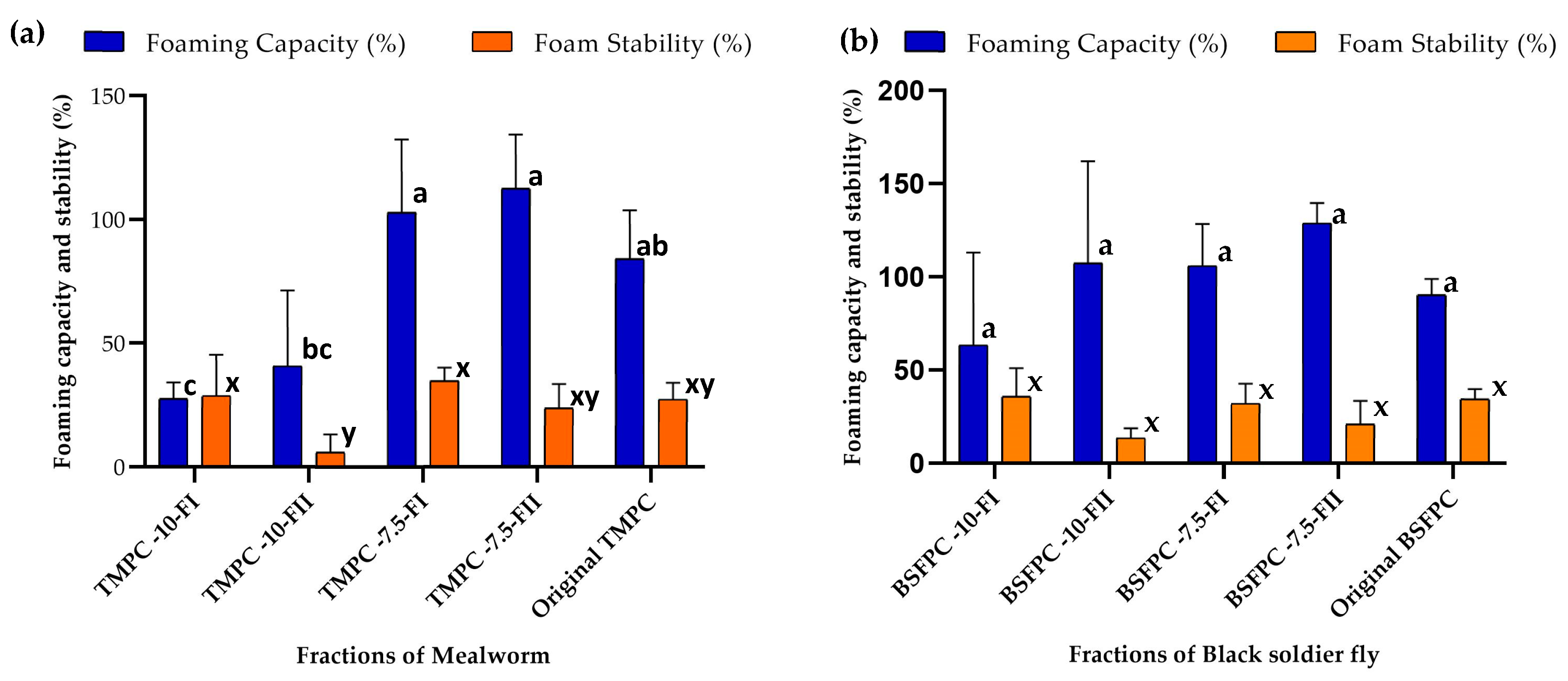
2.8.1. Foaming Capacity and Foam Stability
The foaming properties were analyzed according to the literature [
5,
21] with minor modifications. A sample of 10 mL of 1 g/L protein solution prepared with BSF or TM protein fractions and appropriate buffer (5 mM, pH 7 or pH 5) were placed in a 50 mL plastic tube and subjected to vigorous rotor-stator homogenization (Ultra Turrax T18 digital, IKA, Königswinter, Germany) at 12,000 rpm for 2 min. The height of the foam layer after 10 s and 120 min was recorded. Foaming capacity (FC) and foam stability (FS) (from experiments run in duplicate) were calculated using Equations (2) and (3), respectively [
5]:
where
is the initial height of protein solution in the tube,
is the height of generated foam after agitation,
is the initial foaming capacity (after 10 s) and
is the one obtained after 120 min.
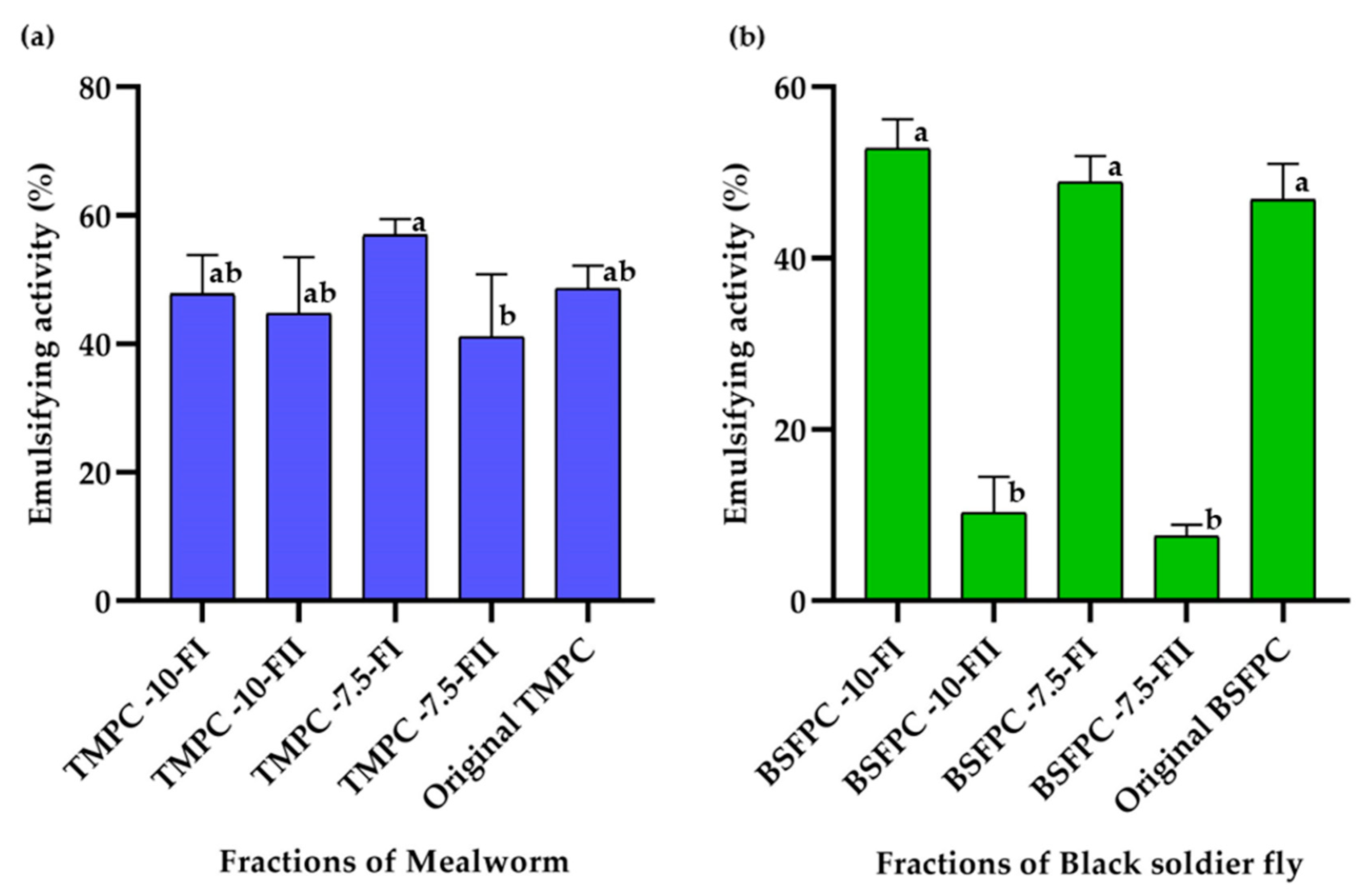
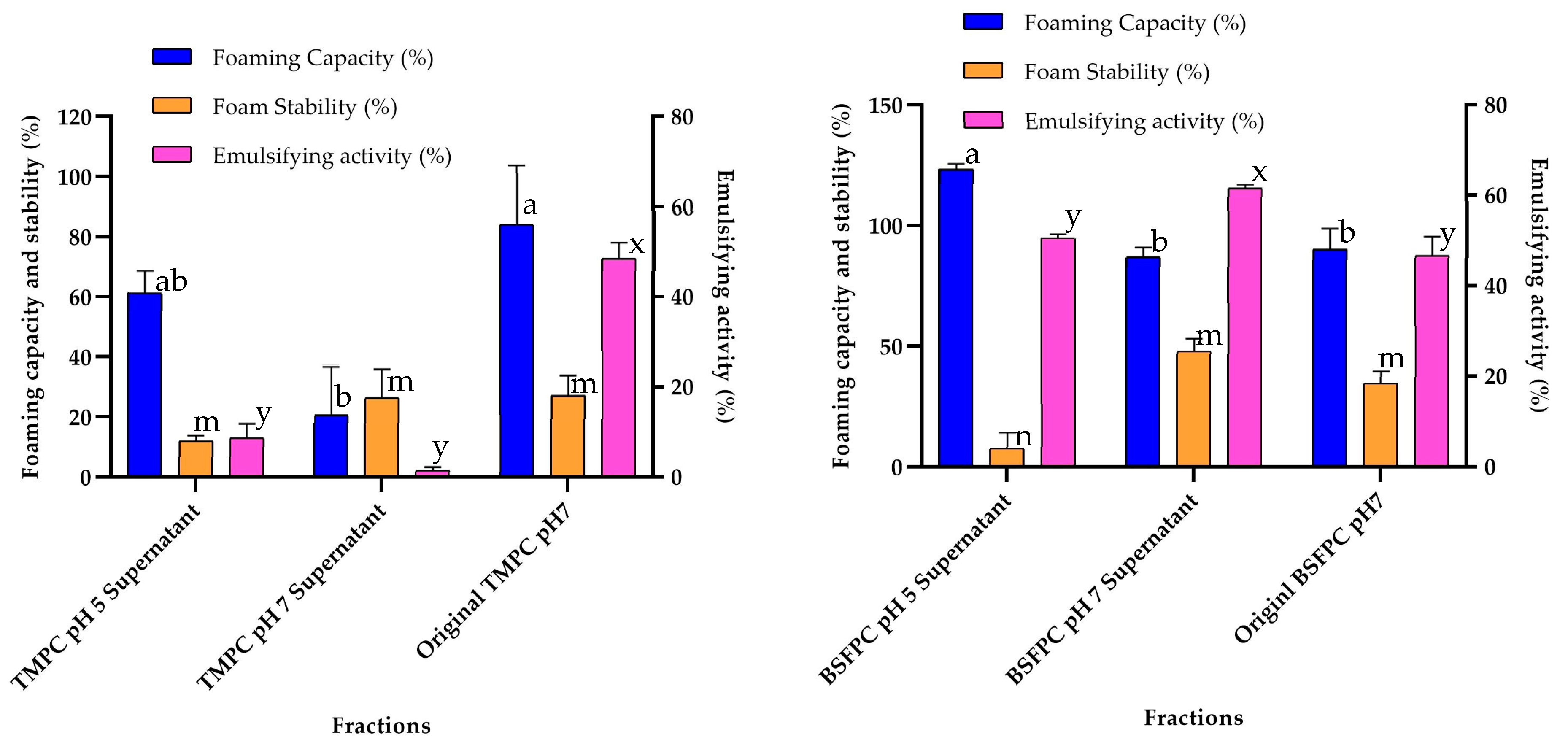
2.8.2. Emulsifying Activity (EA)
EA was always evaluated at 1 g/L protein concentration using the method described by Purschke et al. [
2]. Briefly, 5 mL of protein solution and 5 mL of sunflower oil were homogenized in a beaker using Ultra Turrax T18 digital disperser at 11,000 rpm for 30 s. An aliquot of 9 mL of the emulsion was transferred into a 15 mL scaled tube and centrifuged at 3250×
g for 20 min at room temperature. Duplicates were performed for each sample. The height of the emulsified layer was noted, and the emulsifying activity was calculated using Equation (4) [
5]:
where
is the height of emulsified layer and
is the total height of solution in the tube.
2.9. UF Membrane Characterization
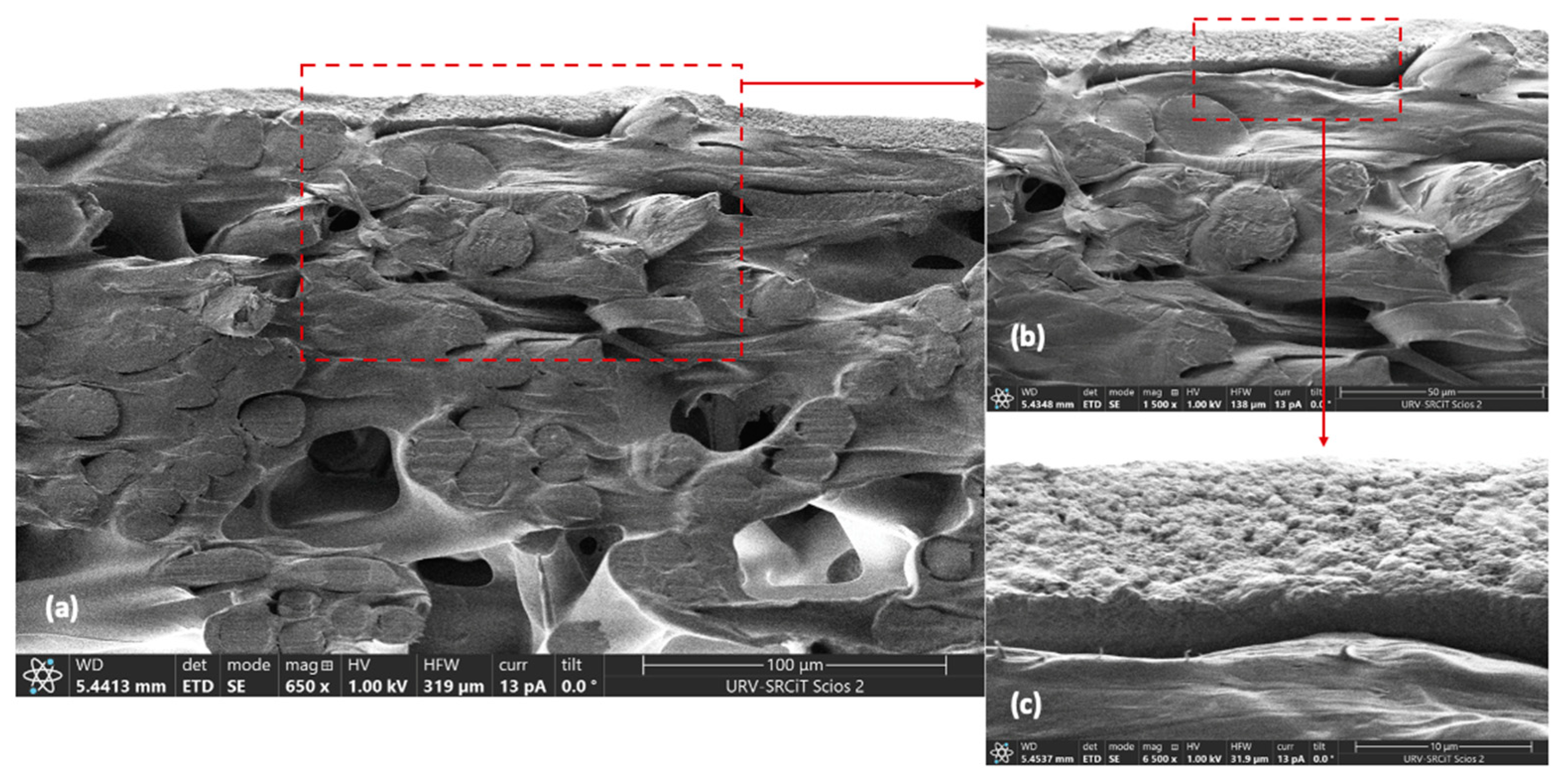
Morphology of cross-sectional area of a new-conditioned membrane was assessed using a field emission scanning electron microscope (FESEM, Scios™ 2, FEI Company Ltd., Hillsboro, OR, USA). Membrane samples were cut using cryogenic scissors and then were fixed on carbon supports to examine under FESEM.
2.10. Statistical Analysis
Results are expressed as mean ± standard deviation. Data from techno-functional properties and zeta potential were analyzed using the general linear model (GLM) at a 95% confidence interval using Minitab 19™ (Minitab Inc., Chicago, IL, USA).


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}