
Effects of Natural Rhamnolipid Mixture on Dioleoylphosphatidylcholine Model Membrane Depending on Method of Preparation and Sterol Content
 , and
, and 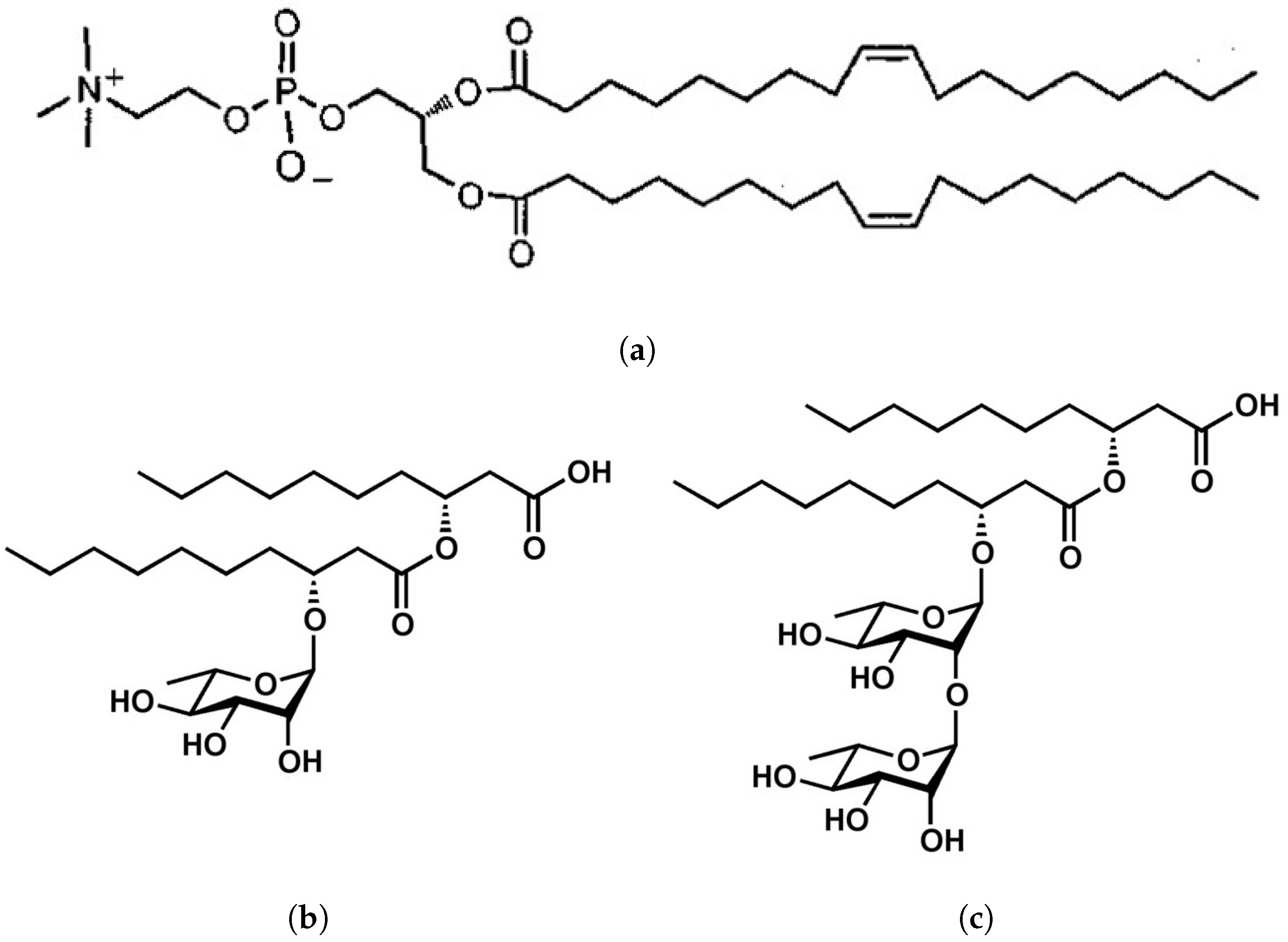{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. NMR Experiments
2.4. ATR-FTIR Experiments
3. Results and Discussion
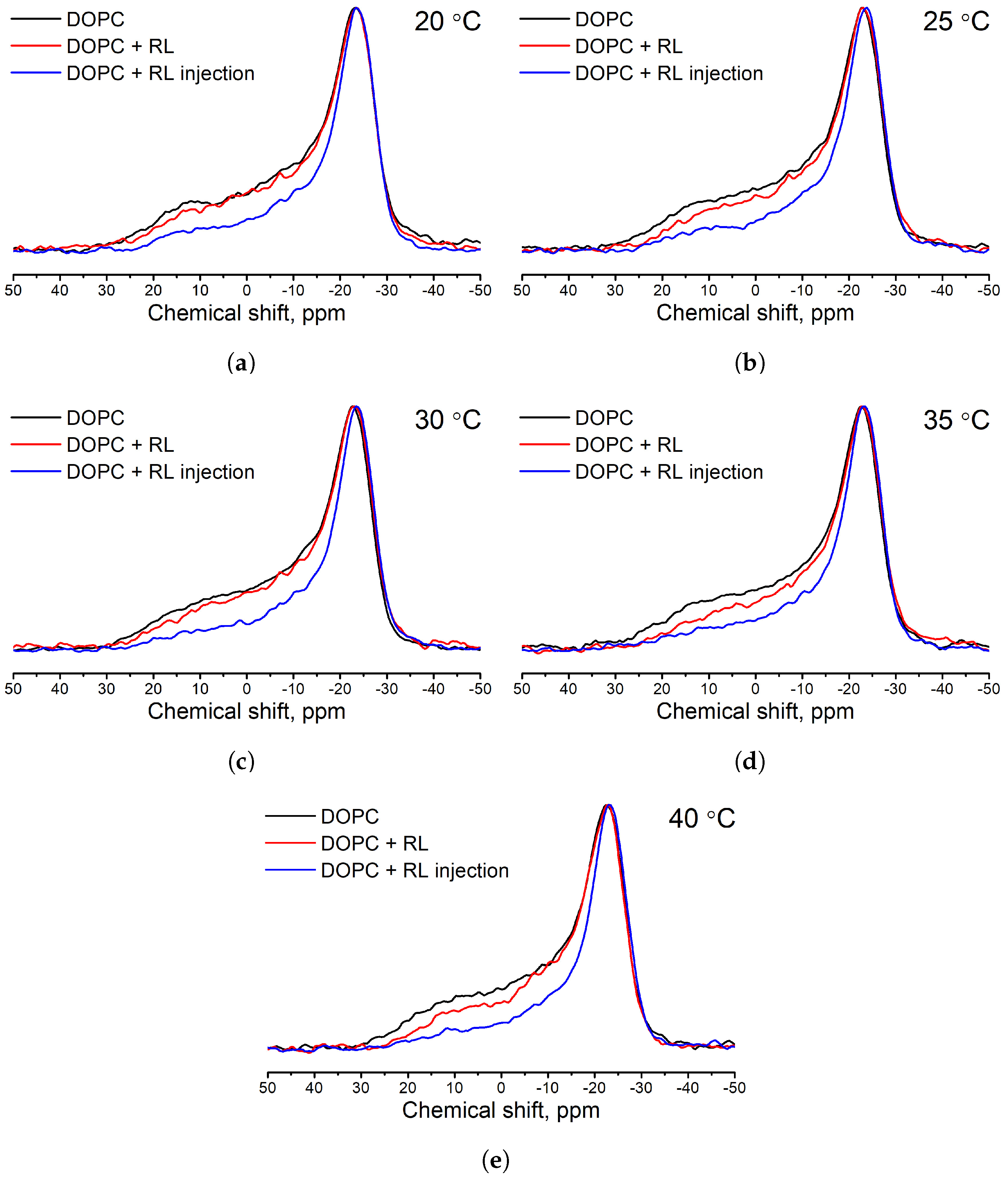
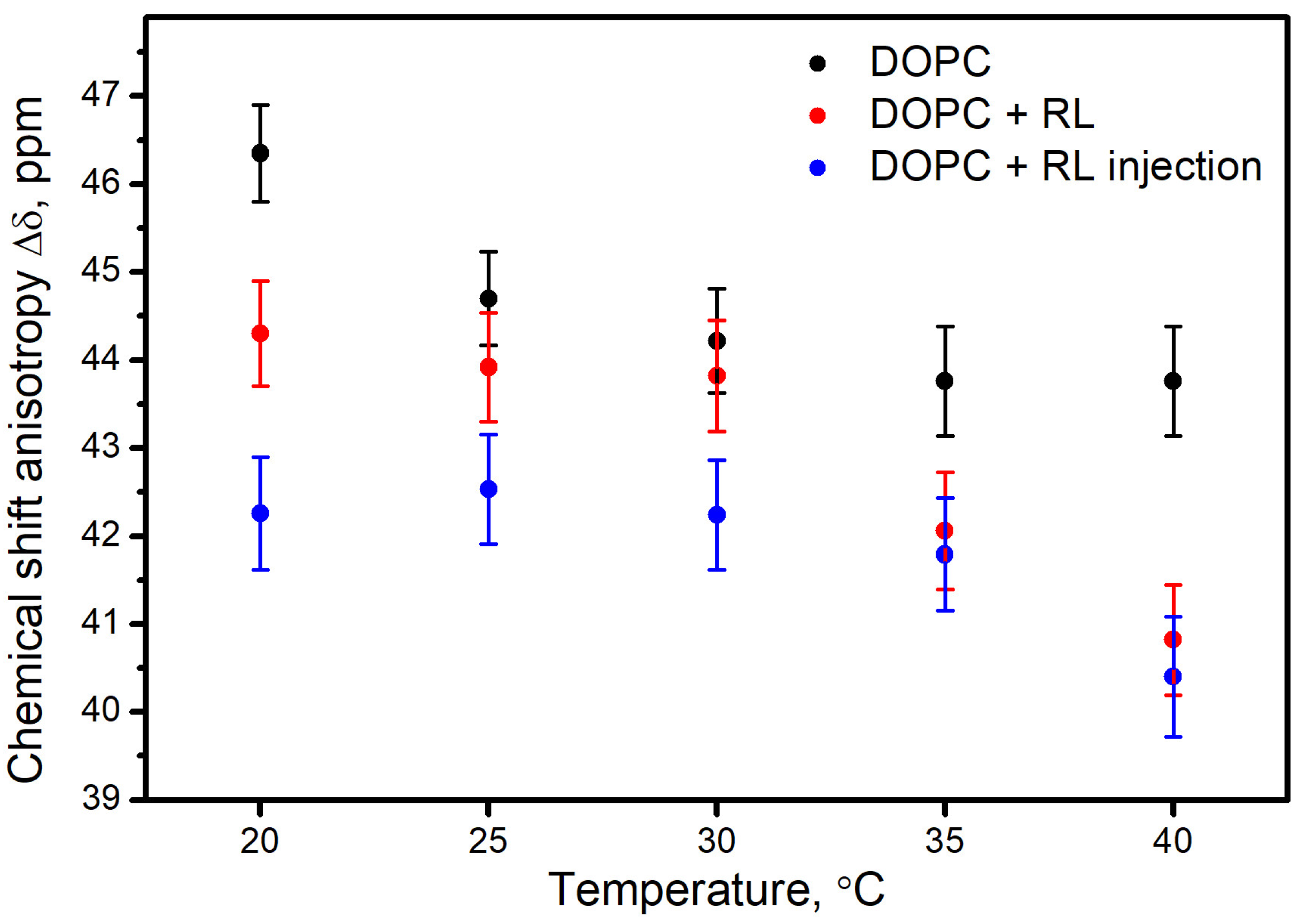
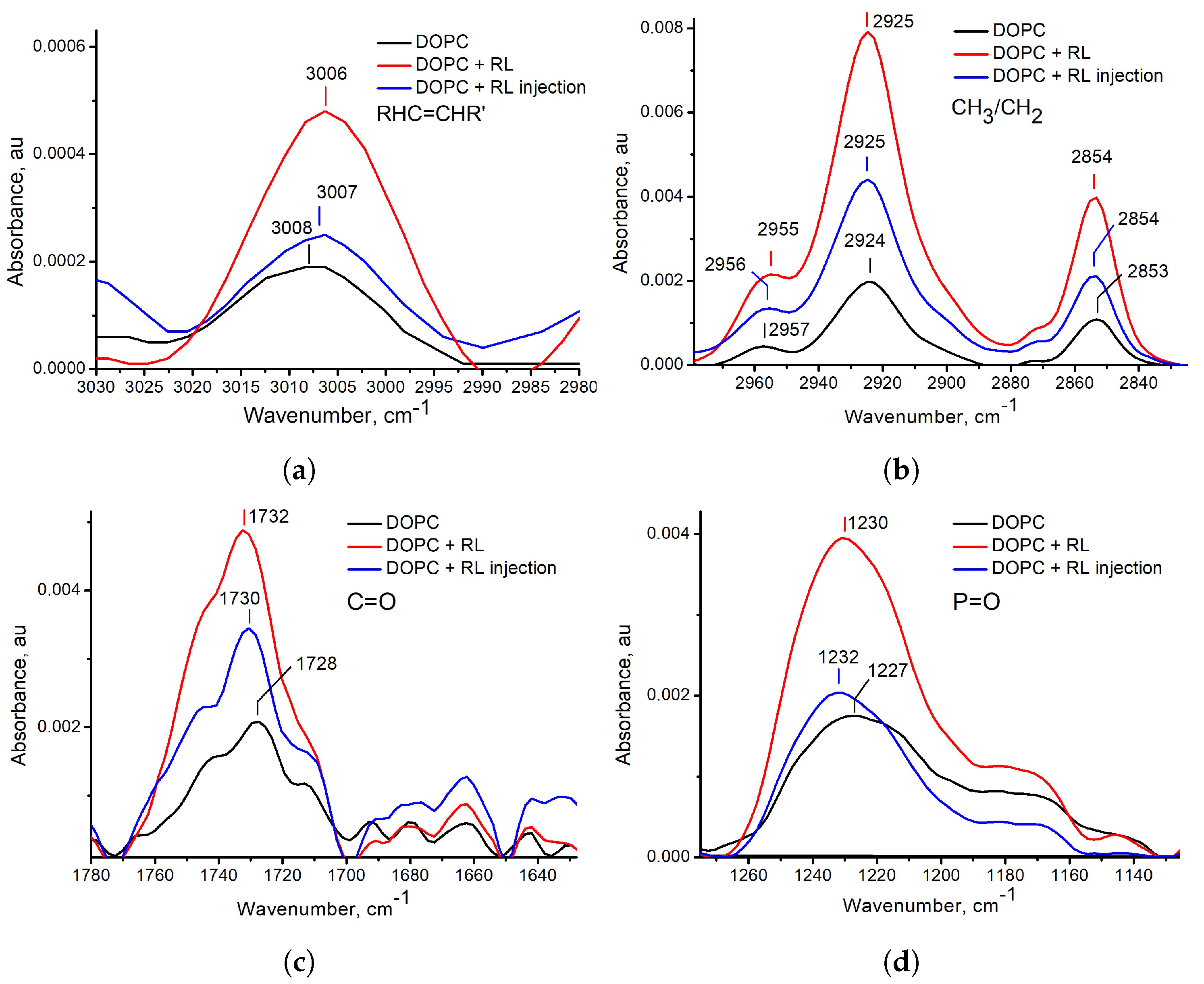
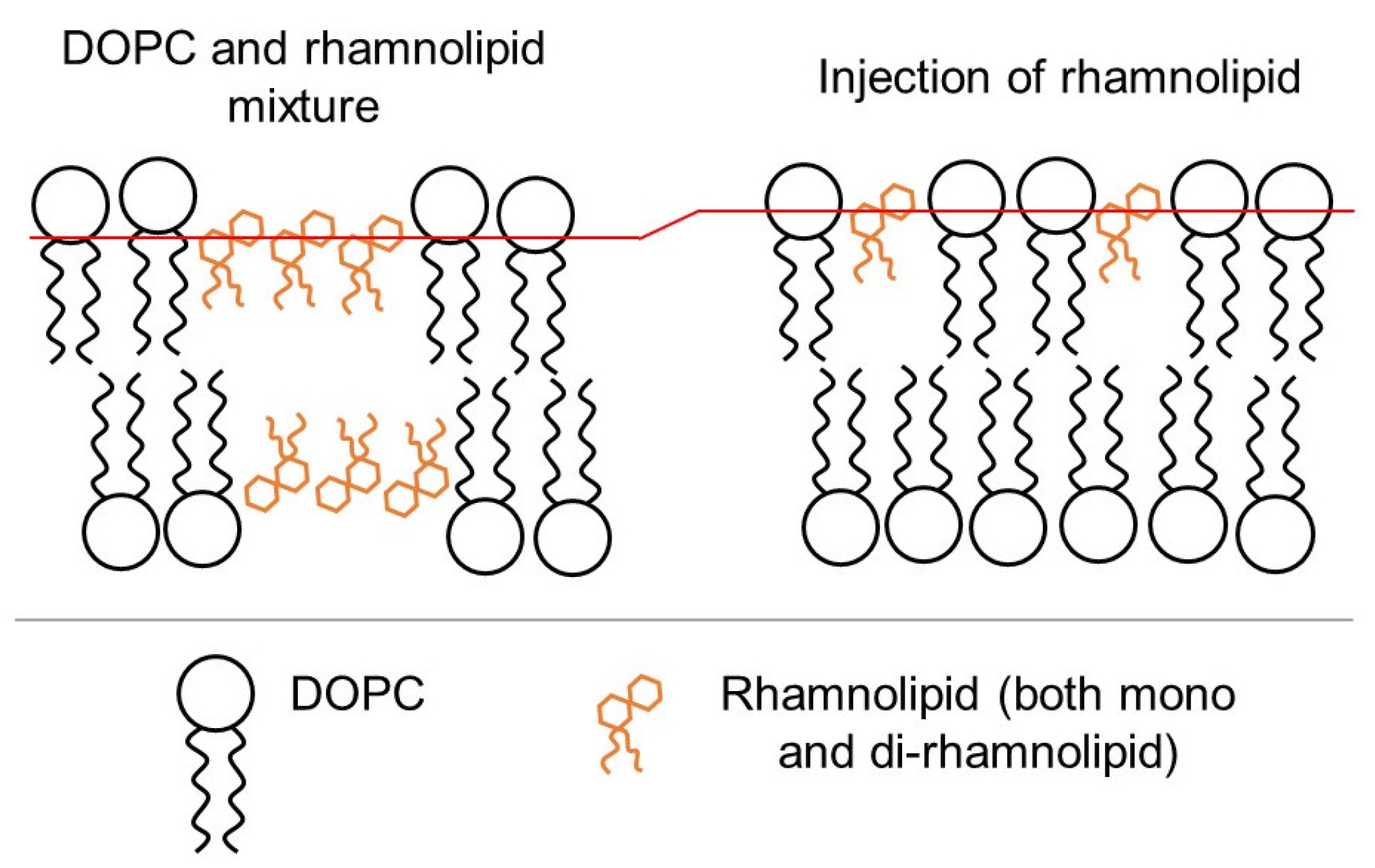
3.1. Influence of the Method of Preparation of Liposomes
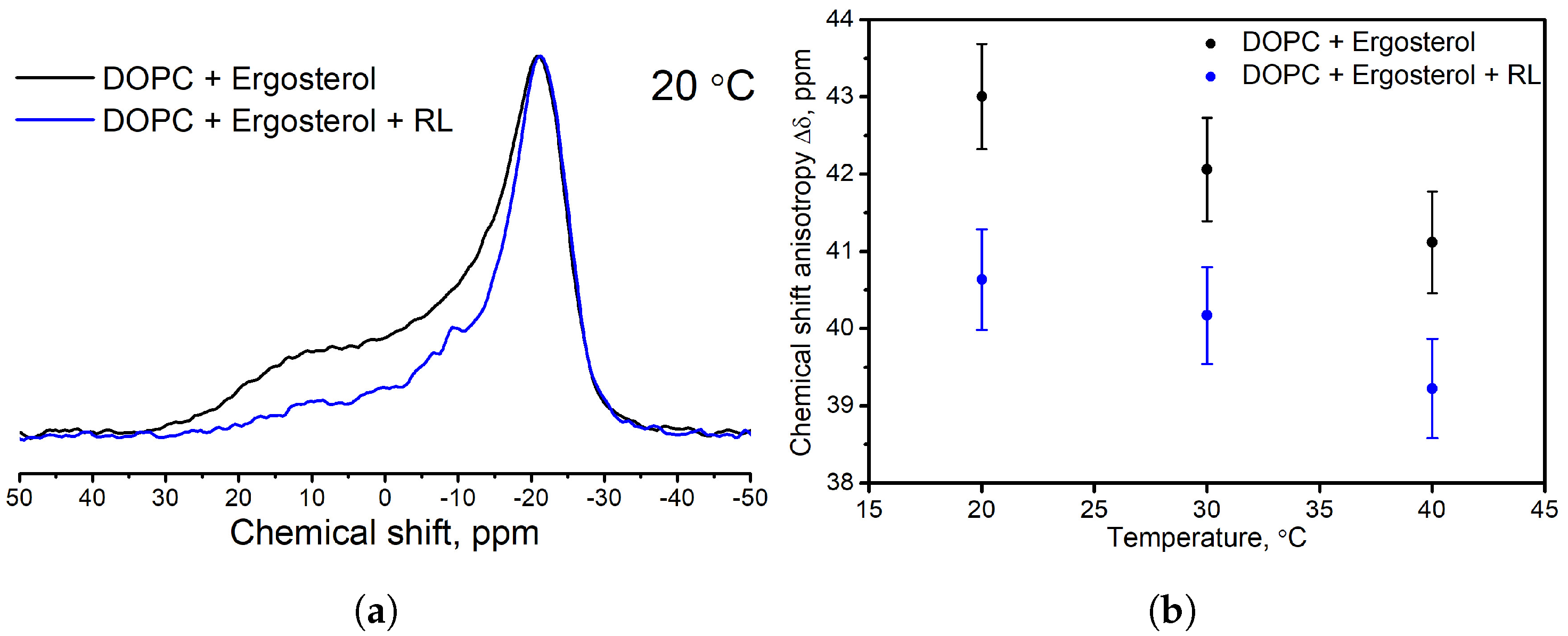
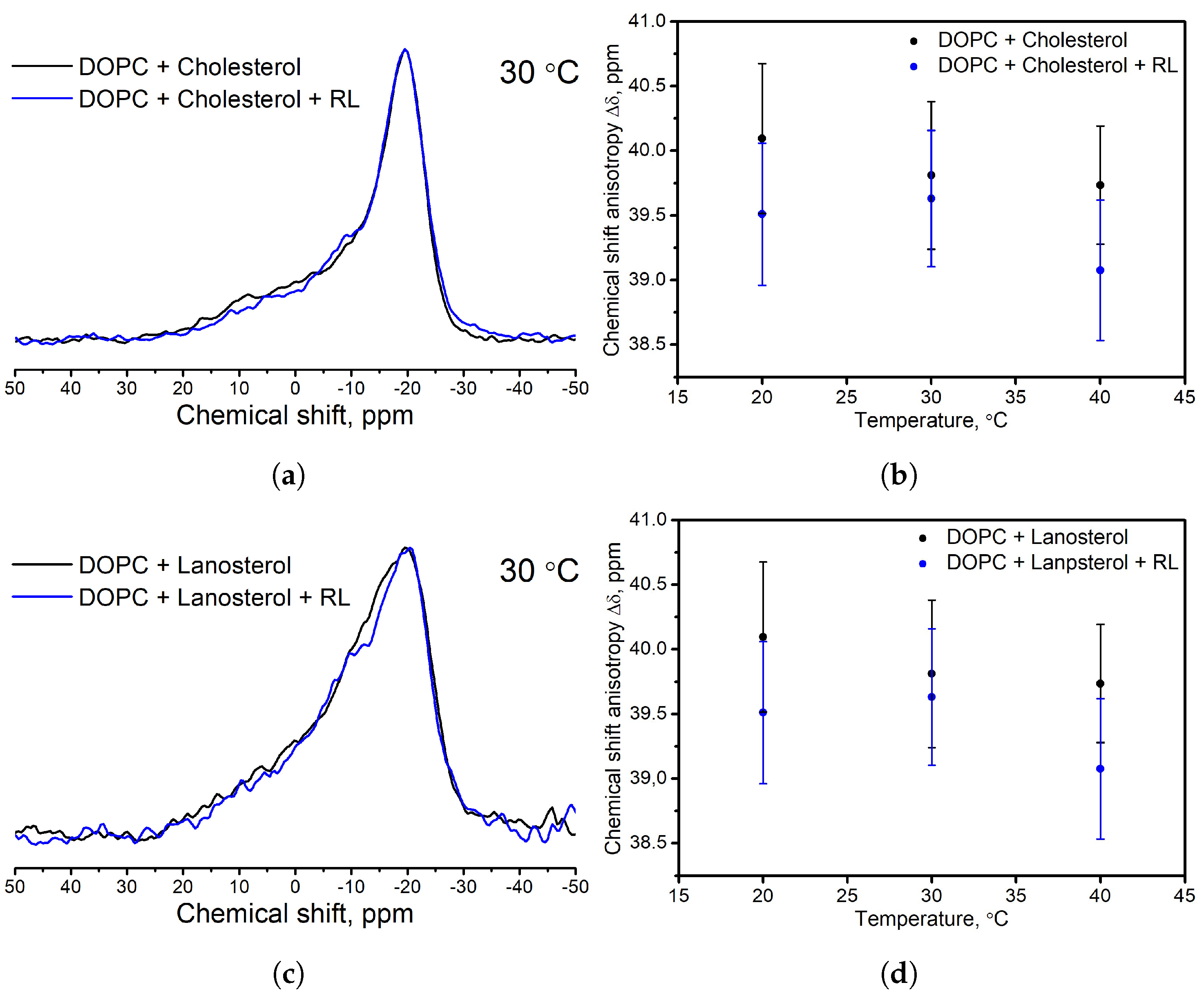
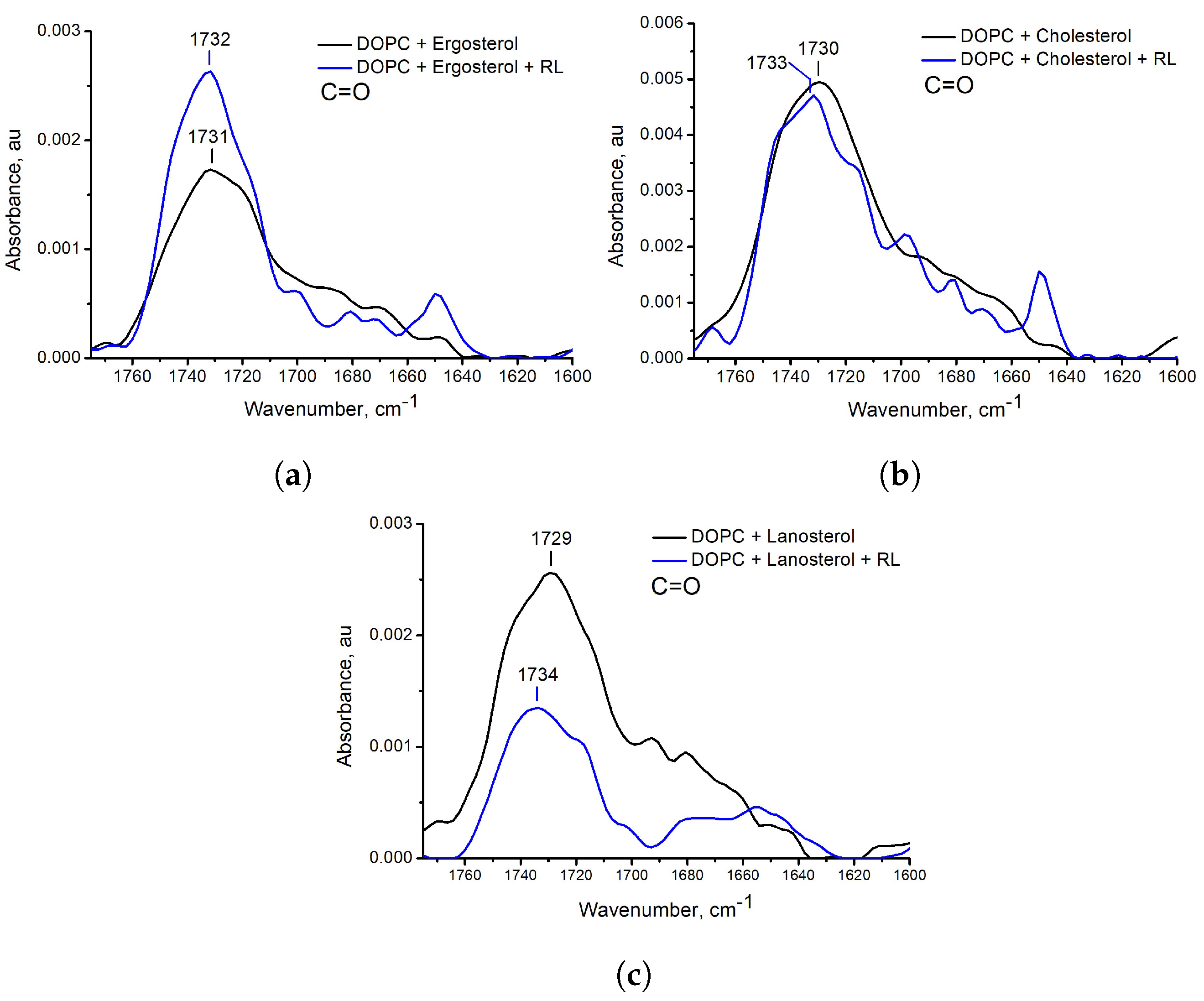
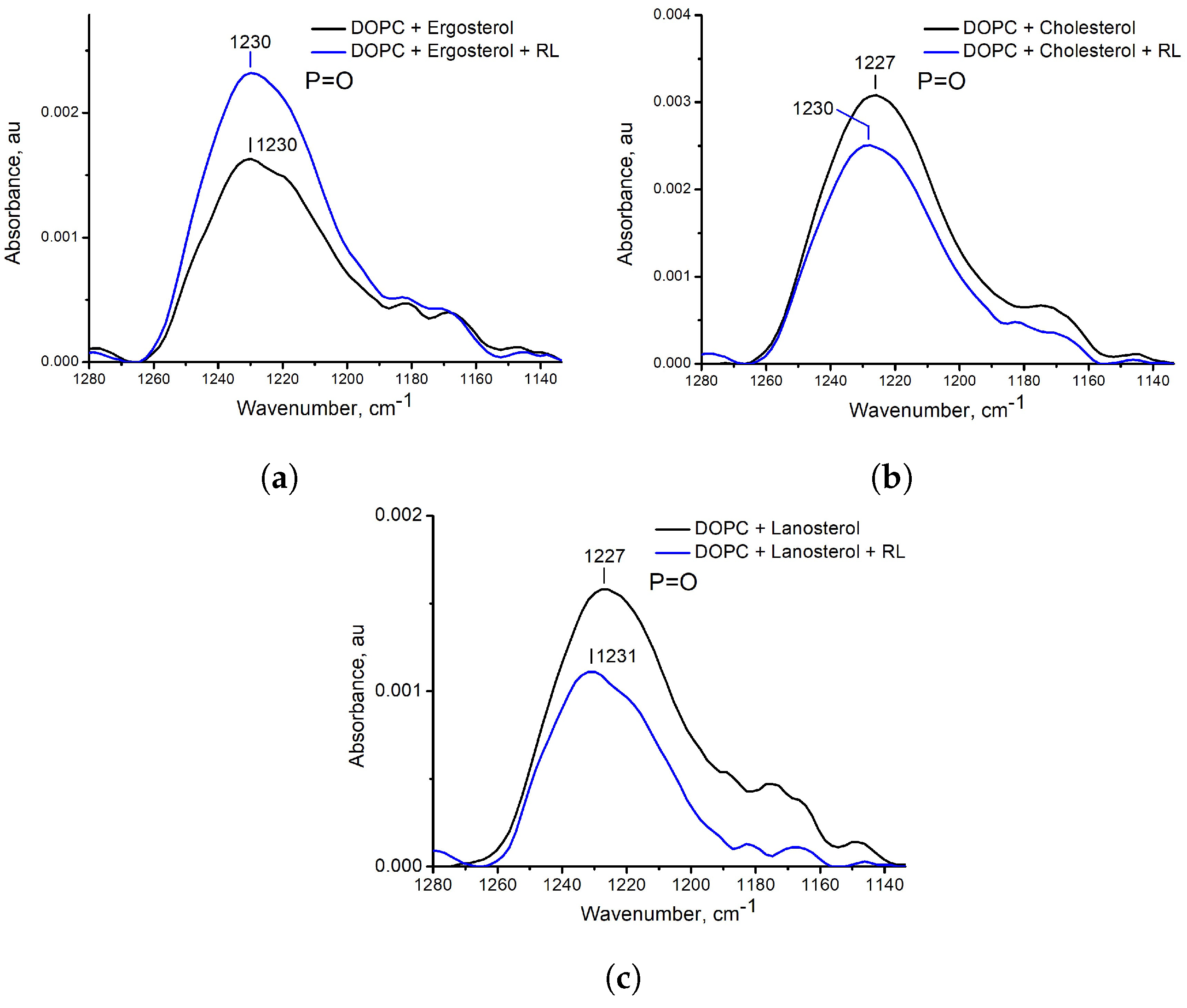
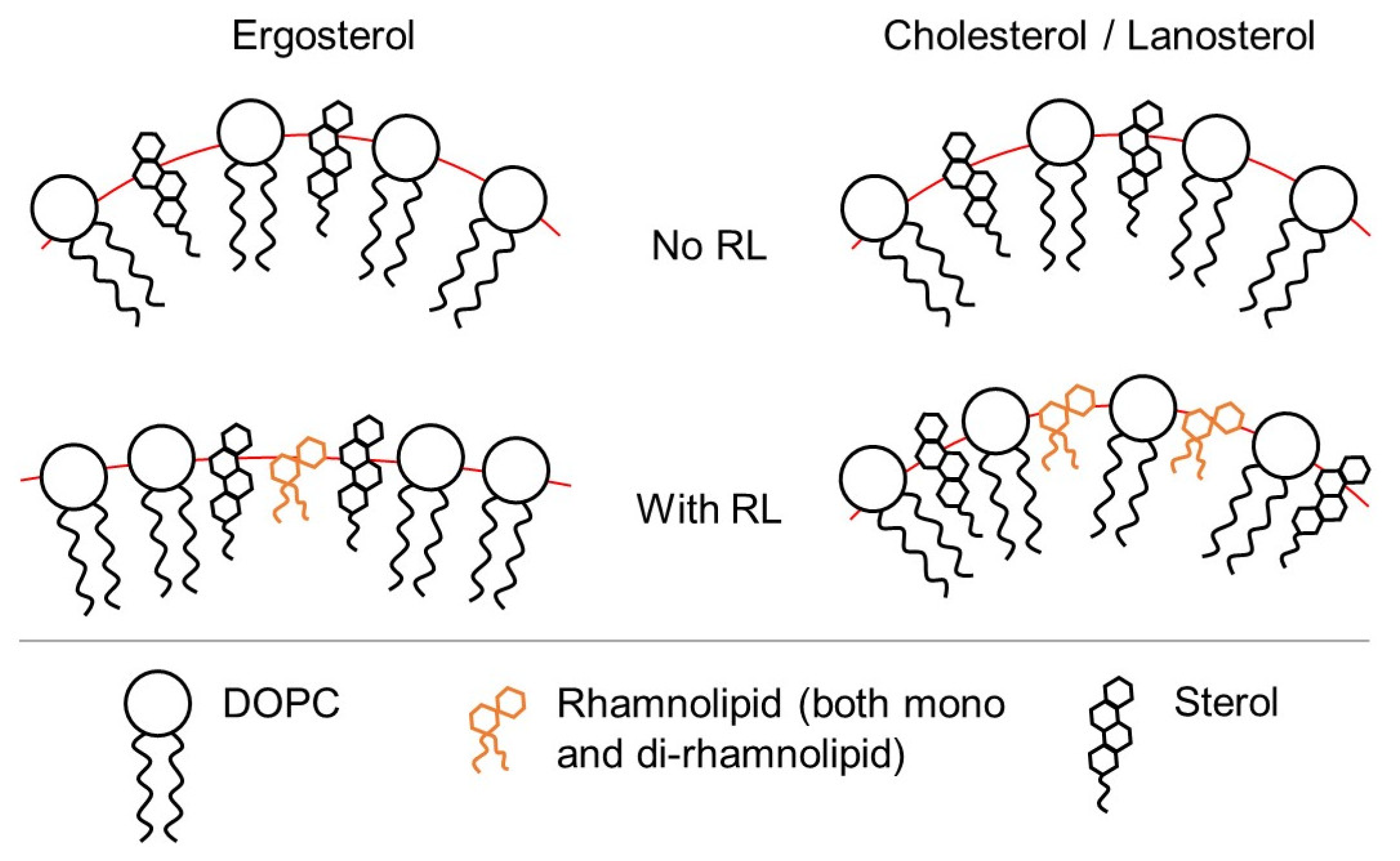
3.2. Influence of the Presence of Sterols
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DOPC | Dioleoylphosphatidylcholine |
| RL | Rhamnolipid |
| NMR | Nuclear magnetic resonance |
| ATR-FTIR | Attenuated total reflection Fourier transform infrared spectroscopy |
References
- Abdel-Mawgoud, A.M.; Lepine, F.; Deziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.C. Biosurfactants: Recent advances. J. Chem. Technol. Biotechnol. Int. Res. Process. Environ. Clean Technol. 1996, 66, 109–120. [Google Scholar]
- Holmberg, K. Natural surfactants. Curr. Opin. Colloid Interface Sci. 2001, 6, 148–159. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [PubMed]
- Balakrishnan, P.; Shanmugam, S.; Lee, W.S.; Lee, W.M.; Kim, J.O.; Oh, D.H.; Kim, D.D.; Kim, J.S.; Yoo, B.K.; Choi, H.G. Formulation and in vitro assessment of minoxidil niosomes for enhanced skin delivery. Int. J. Pharm. 2009, 377, 1–8. [Google Scholar] [CrossRef]
- Gunther, N.W., IV; Nunez, A.; Fett, W.; Solaiman, D.K. Production of rhamnolipids by Pseudomonas chlororaphis, a nonpathogenic bacterium. Appl. Environ. Microbiol. 2005, 71, 2288–2293. [Google Scholar] [CrossRef] [Green Version]
- Gunther, N.W., IV; Nunez, A.; Fortis, L.; Solaiman, D.K. Proteomic based investigation of rhamnolipid production by Pseudomonas chlororaphis strain NRRL B-30761. J. Ind. Microbiol. Biotechnol. 2006, 33, 914–920. [Google Scholar] [CrossRef]
- Onbasli, D.; Aslim, B. Biosurfactant production in sugar beet molasses by some Pseudomonas spp. J. Environ. Biol. 2009, 30, 161–163. [Google Scholar]
- Muller, M.M.; Hausmann, R. Regulatory and metabolic network of rhamnolipid biosynthesis: Traditional and advanced engineering towards biotechnological production. Appl. Microbiol. Biotechnol. 2011, 91, 251–264. [Google Scholar] [CrossRef]
- Soberon-Chavez, G.; Lepine, F.; Deziel, E. Production of rhamnolipids by Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2005, 68, 718–725. [Google Scholar] [CrossRef]
- Shi, J.; Chen, Y.; Liu, X.; Li, D. Rhamnolipid production from waste cooking oil using newly isolated halotolerant Pseudomonas aeruginosa M4. J. Clean. Prod. 2021, 278, 123879. [Google Scholar] [CrossRef]
- Aguirre-Ramirez, M.; Silva-Jimenez, H.; Banat, I.M.; Diaz De Rienzo, M.A. Surfactants: Physicochemical interactions with biological macromolecules. Biotechnol. Lett. 2021, 43, 523–535. [Google Scholar] [CrossRef]
- Itoh, S.; Honda, H.; Tomita, F.; Suzuki, T. Rhamnolipids produced by Pseudomonas aeruginosa grown on n-paraffin (mixture of C12, C13 and C14 fractions). J. Antibiot. 1971, 24, 855–859. [Google Scholar] [CrossRef]
- Arino, S.; Marchal, R.; Vandecasteele, J.P. Involvement of a rhamnolipid-producing strain of Pseudomonas aeruginosa in the degradation of polycyclic aromatic hydrocarbons by a bacterial community. J. Appl. Microbiol. 1998, 84, 769–776. [Google Scholar] [CrossRef]
- Elshikh, M.; Moya-Ramirez, I.; Moens, H.; Roelants, S.; Soetaert, W.; Marchant, R.; Banat, I.M. Rhamnolipids and lactonic sophorolipids: Natural antimicrobial surfactants for oral hygiene. J. Appl. Microbiol. 2017, 123, 1111–1123. [Google Scholar] [CrossRef]
- Vatsa, P.; Sanchez, L.; Clement, C.; Baillieul, F.; Dorey, S. Rhamnolipid biosurfactants as new players in animal and plant defense against microbes. Int. J. Mol. Sci. 2010, 11, 5095–5108. [Google Scholar] [CrossRef] [Green Version]
- Varnier, A.L.; Sanchez, L.; Vatsa, P.; Boudesocque, L.; Garcia-Brugger, A.; Rabenoelina, F.; Sorokin, A.; Renault, J.H.; Kauffmann, S.; Pugin, A. Bacterial rhamnolipids are novel MAMPs conferring resistance to Botrytis cinerea in grapevine. Plant Cell Environ. 2009, 32, 178–193. [Google Scholar] [CrossRef]
- Ranf, S. Sensing of molecular patterns through cell surface immune receptors. Curr. Opin. Plant Biol. 2017, 38, 68–77. [Google Scholar] [CrossRef]
- Chen, J.; Wu, Q.; Hua, Y.; Chen, J.; Zhang, H.; Wang, H. Potential applications of biosurfactant rhamnolipids in agriculture and biomedicine. Appl. Microbiol. Biotechnol. 2017, 101, 8309–8319. [Google Scholar] [CrossRef]
- Yan, F.; Hu, H.; Lu, L.; Zheng, X. Rhamnolipids induce oxidative stress responses in cherry tomato fruit to Alternaria alternata. Pest Manag. Sci. 2016, 72, 1500–1507. [Google Scholar] [CrossRef]
- Stanghellini, M.E.; Miller, R.M. Biosurfactants: Their identity and potential efficacy in the biological control of zoosporic plant pathogens. Plant Dis. 1997, 81, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.S.; Lee, J.Y.; Hwang, B.K. In vivo control and in vitro antifungal activity of rhamnolipid B, a glycolipid antibiotic, against Phytophthora capsici and Colletotrichum orbiculare. Pest Manag. Sci. Former. Pestic. Sci. 2000, 56, 1029–1035. [Google Scholar] [CrossRef]
- Yan, F.; Xu, S.; Chen, Y.; Zheng, X. Effect of rhamnolipids on Rhodotorula glutinis biocontrol of Alternaria alternata infection in cherry tomato fruit. Postharvest Biol. Technol. 2014, 97, 32–35. [Google Scholar] [CrossRef]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. Chemical structure and in vitro antitumor activity of rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol. 2013, 170, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Pacwa-Płociniczak, M.; Płaza, G.; Piotrowska-Seget, Z.; Cameotra, S. Environmental Applications of Biosurfactants: Recent Advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef]
- Hogan, D.E.; Tian, F.; Malm, S.W.; Olivares, C.; Pacheco, R.P.; Simonich, M.T.; Hunjan, A.S.; Tanguay, R.L.; Klimecki, W.T.; Polt, R.; et al. Biodegradability and toxicity of monorhamnolipid biosurfactant diastereomers. J. Hazard. Mater. 2019, 364, 600–607. [Google Scholar] [CrossRef]
- Gidudu, B.; Chirwa, E.M. Evaluation of the toxicity of a rhamnolipid biosurfactant for its application in the optimization of the bio-electrokinetic remediation of petrochemical contaminated soil. Clean. Eng. Technol. 2022, 9, 100521. [Google Scholar] [CrossRef]
- Monnier, N.; Furlan, A.L.; Buchoux, S.; Deleu, M.; Dauchez, M.; Rippa, S.; Sarazin, C. Exploring the dual interaction of natural rhamnolipids with plant and fungal biomimetic plasma membranes through biophysical studies. Int. J. Mol. Sci. 2019, 20, 1009. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; Teruel, J.A.; Espuny, M.J.; Marques, A.; Manresa, A.; Aranda, F.J. Effects of dirhamnolipid on the structural properties of phosphatidylcholine membranes. Int. J. Pharm. 2006, 325, 99–107. [Google Scholar] [CrossRef]
- Sanchez, M.; Teruel, J.A.; Espuny, M.J.; Marques, A.; Aranda, F.J.; Manresa, A.; Ortiz, A. Modulation of the physical properties of dielaidoylphosphatidylethanolamine membranes by a dirhamnolipid biosurfactant produced by Pseudomonas aeruginosa. Chem. Phys. Lipids 2006, 142, 118–127. [Google Scholar] [CrossRef]
- Aranda, F.J.; Teruel, J.A.; Espuny, M.J.; Marques, A.; Manresa, A.; Palacios-Lidón, E.; Ortiz, A. Domain formation by a Rhodococcus sp. biosurfactant trehalose lipid incorporated into phosphatidylcholine membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 2596–2604. [Google Scholar] [CrossRef] [Green Version]
- Haba, E.; Pinazo, A.; Pons, R.; Perez, L.; Manresa, A. Complex rhamnolipid mixture characterization and its influence on DPPC bilayer organization. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 776–783. [Google Scholar] [CrossRef]
- Oliva, A.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. Effect of a dirhamnolipid biosurfactant on the structure and phase behaviour of dimyristoylphosphatidylserine model membranes. Colloids Surf. B Biointerfaces 2020, 185, 110576. [Google Scholar] [CrossRef]
- Sanchez, M.; Aranda, F.J.; Teruel, J.A.; Espuny, M.J.; Marques, A.; Manresa, A.; Ortiz, A. Permeabilization of biological and artificial membranes by a bacterial dirhamnolipid produced by Pseudomonas aeruginosa. J. Colloid Interface Sci. 2010, 341, 240–247. [Google Scholar] [CrossRef]
- Gennis, R.B. Biomembranes: Molecular Structure and Function; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Deleu, M.; Crowet, J.M.; Nasir, M.N.; Lins, L. Complementary biophysical tools to investigate lipid specificity in the interaction between bioactive molecules and the plasma membrane: A review. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 3171–3190. [Google Scholar] [CrossRef] [Green Version]
- Urbina, J.A.; Pekerar, S.; Le, H.B.; Patterson, J.; Montez, B.; Oldfield, E. Molecular order and dynamics of phosphatidylcholine bilayer membranes in the presence of cholesterol, ergosterol and lanosterol: A comparative study using 2H-, 13C-and 31P-NMR spectroscopy. Biochim. Biophys. Acta (BBA)-Biomembr. 1995, 1238, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Uahengo, V.; Likius, D. Mini review on emerging methods of preparation of liposome and its application as Liposome drug delivery systems. Open J. Pharmacol. Pharmacother. 2018, 3, 005–021. [Google Scholar]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252, IN26-IN27. [Google Scholar] [CrossRef]
- Lasic, D.D. Novel applications of liposomes. Trends Biotechnol. 1998, 16, 307–321. [Google Scholar] [CrossRef]
- Traikia, M.; Warschawski, D.E.; Recouvreur, M.; Cartaud, J.; Devaux, P.F. Formation of unilamellar vesicles by repetitive freeze-thaw cycles: Characterization by electron microscopy and 31P-nuclear magnetic resonance. Eur. Biophys. J. 2000, 29, 184–195. [Google Scholar] [CrossRef]
- Filippov, A.V.; Khakimov, A.M.; Munavirov, B.V. 31P NMR Studies of Phospholipids. In Annual Reports on NMR Spectroscopy; Elsevier: Amsterdam, The Netherlands, 2015; Volume 85, pp. 27–92. [Google Scholar]
- Schiller, J.; Muller, M.; Fuchs, B.; Arnold, K.; Huster, D. 31P NMR spectroscopy of phospholipids: From micelles to membranes. Curr. Anal. Chem. 2007, 3, 283–301. [Google Scholar] [CrossRef]
- Holland, G.P.; McIntyre, S.K.; Alam, T.M. Distinguishing individual lipid headgroup mobility and phase transitions in raft-forming lipid mixtures with 31P MAS NMR. Biophys. J. 2006, 90, 4248–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massiot, D.; Fayon, F.; Capron, M.; King, I.; Le Calvé, S.; Alonso, B.; Durand, J.O.; Bujoli, B.; Gan, Z.; Hoatson, G. Modelling one-and two-dimensional solid-state NMR spectra. Magn. Reson. Chem. 2002, 40, 70–76. [Google Scholar] [CrossRef]
- Seelig, J. 31P nuclear magnetic resonance and the head group structure of phospholipids in membranes. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1978, 515, 105–140. [Google Scholar] [CrossRef]
- Burnell, E.E.; Cullis, P.R.; De Kruijff, B. Effects of tumbling and lateral diffusion on phosphatidylcholine model membrane 31P-NMR lineshapes. Biochim. Biophys. Acta (BBA)-Biomembr. 1980, 603, 63–69. [Google Scholar] [CrossRef]
- Herzog, M.; Tiso, T.; Blank, L.M.; Winter, R. Interaction of rhamnolipids with model biomembranes of varying complexity. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183431. [Google Scholar] [CrossRef]
- Furlan, A.L.; Laurin, Y.; Botcazon, C.; Rodríguez-Moraga, N.; Rippa, S.; Deleu, M.; Lins, L.; Sarazin, C.; Buchoux, S. Contributions and limitations of biophysical approaches to study of the interactions between amphiphilic molecules and the plant plasma membrane. Plants 2020, 9, 648. [Google Scholar] [CrossRef]
- Timoszyk, A. Dynamics of Model Membranes by NMR. In Spectroscopic Analyses-Developments and Applications; IntechOpen: London, UK, 2017. [Google Scholar]
- Jezowska, I.; Wolak, A.; Gruszecki, W.I.; Strza\lka, K. Effect of β-carotene on structural and dynamic properties of model phosphatidylcholine membranes. II. A 31P-NMR and 13C-NMR study. Biochim. Biophys. Acta (BBA)-Biomembr. 1994, 1194, 143–148. [Google Scholar] [CrossRef]
- Fa, N.; Ronkart, S.; Schanck, A.; Deleu, M.; Gaigneaux, A.; Goormaghtigh, E.; Mingeot-Leclercq, M.P. Effect of the antibiotic azithromycin on thermotropic behavior of DOPC or DPPC bilayers. Chem. Phys. Lipids 2006, 144, 108–116. [Google Scholar] [CrossRef]
- Wang, F.; Pan, S.; Zhang, P.; Fan, H.; Chen, Y.; Yan, J. Synthesis and application of phosphorus-containing flame retardant plasticizer for polyvinyl chloride. Fibers Polym. 2018, 19, 1057–1063. [Google Scholar] [CrossRef]
- Derenne, A.; Claessens, T.; Conus, C.; Goormaghtigh, E. Infrared spectroscopy of membrane lipids. In Encyclopedia of Biophysics; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1074–1081. [Google Scholar]
- Arrondo, J.L.R.; Goni, F.M. Infrared studies of protein-induced perturbation of lipids in lipoproteins and membranes. Chem. Phys. Lipids 1998, 96, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Fernendez, J.C.; Villalain, J. The use of FT-IR for quantitative studies of the apparent pKa of lipid carboxyl groups and the dehydration degree of the phosphate group of phospholipids. Chem. Phys. Lipids 1998, 96, 41–52. [Google Scholar] [CrossRef]
- Abbasi, H.; Noghabi, K.A.; Ortiz, A. Interaction of a bacterial monorhamnolipid secreted by Pseudomonas aeruginosa MA01 with phosphatidylcholine model membranes. Chem. Phys. Lipids 2012, 165, 745–752. [Google Scholar] [CrossRef]
- Lewis, R.N.; McElhaney, R.N.; Pohle, W.; Mantsch, H.H. Components of the carbonyl stretching band in the infrared spectra of hydrated 1, 2-diacylglycerolipid bilayers: A reevaluation. Biophys. J. 1994, 67, 2367–2375. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potapov, K.; Gordeev, A.; Biktasheva, L.; Rudakova, M.; Alexandrov, A. Effects of Natural Rhamnolipid Mixture on Dioleoylphosphatidylcholine Model Membrane Depending on Method of Preparation and Sterol Content. Membranes 2023, 13, 112. https://doi.org/10.3390/membranes13010112
Potapov K, Gordeev A, Biktasheva L, Rudakova M, Alexandrov A. Effects of Natural Rhamnolipid Mixture on Dioleoylphosphatidylcholine Model Membrane Depending on Method of Preparation and Sterol Content. Membranes. 2023; 13(1):112. https://doi.org/10.3390/membranes13010112
Chicago/Turabian StylePotapov, Konstantin, Alexander Gordeev, Liliya Biktasheva, Maya Rudakova, and Artem Alexandrov. 2023. "Effects of Natural Rhamnolipid Mixture on Dioleoylphosphatidylcholine Model Membrane Depending on Method of Preparation and Sterol Content" Membranes 13, no. 1: 112. https://doi.org/10.3390/membranes13010112