
Model Evaluation of the Microbial Metabolic Processes in a Hydrogen-Based Membrane Biofilm Reactor for Simultaneous Bromate and Nitrate Reduction


Abstract
:1. Introduction
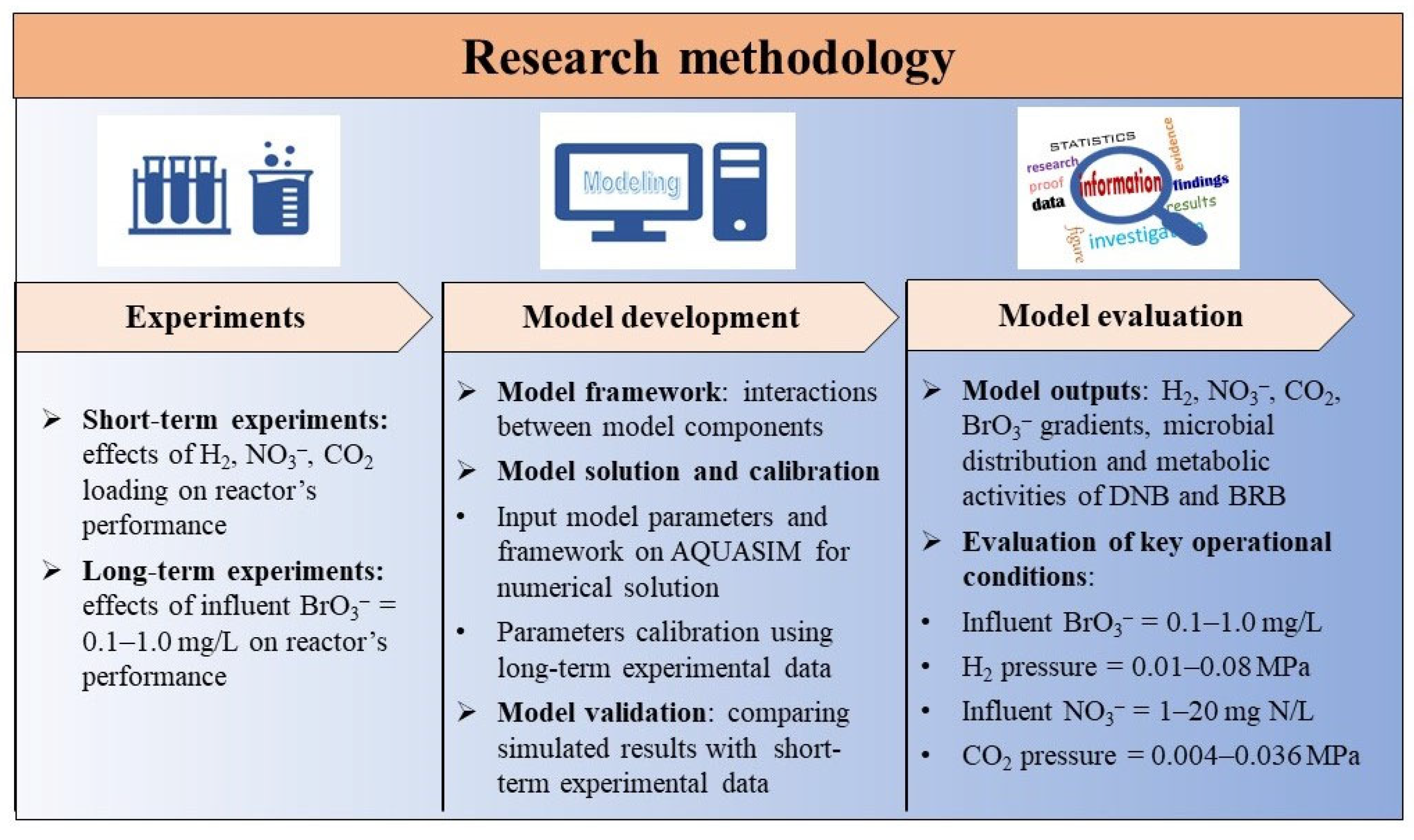
2. Materials and Methods
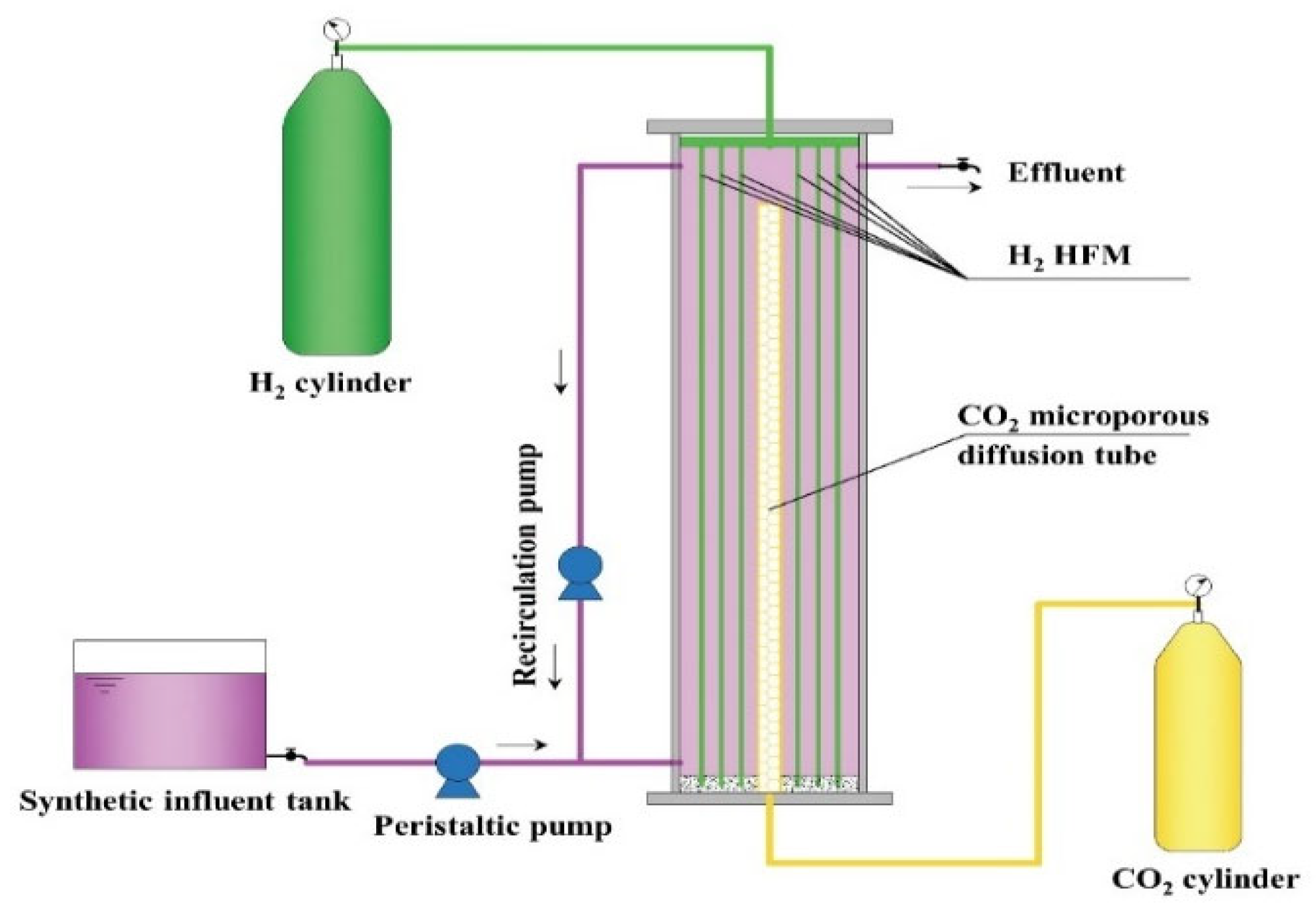
2.1. Reactor Setup and Start-Up
2.2. Short- and Long-Term Experiments and Sample Analysis
2.3. Model Development and Evaluation
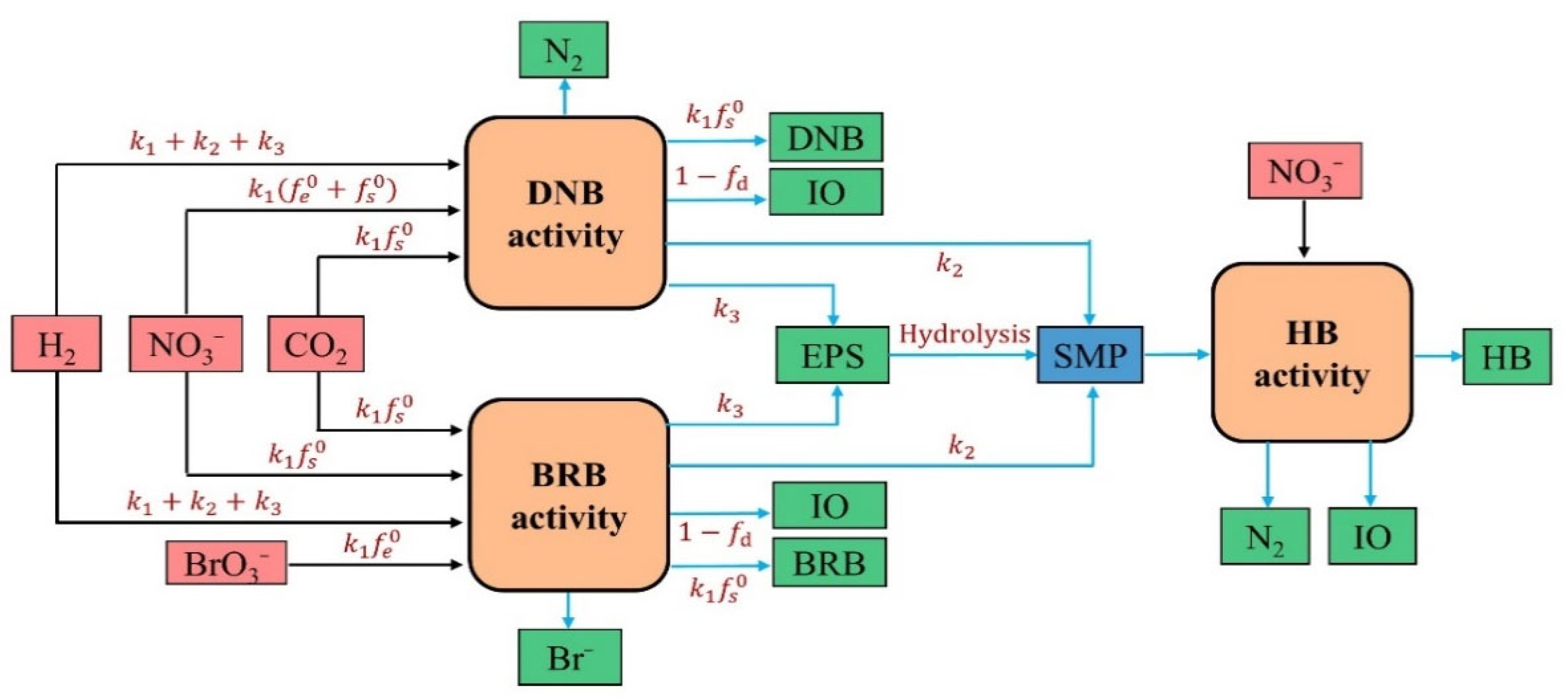
2.3.1. Model Framework Development
2.3.2. Model Solution and Calibration
2.3.3. Model Validation and Evaluation
3. Results and Discussion
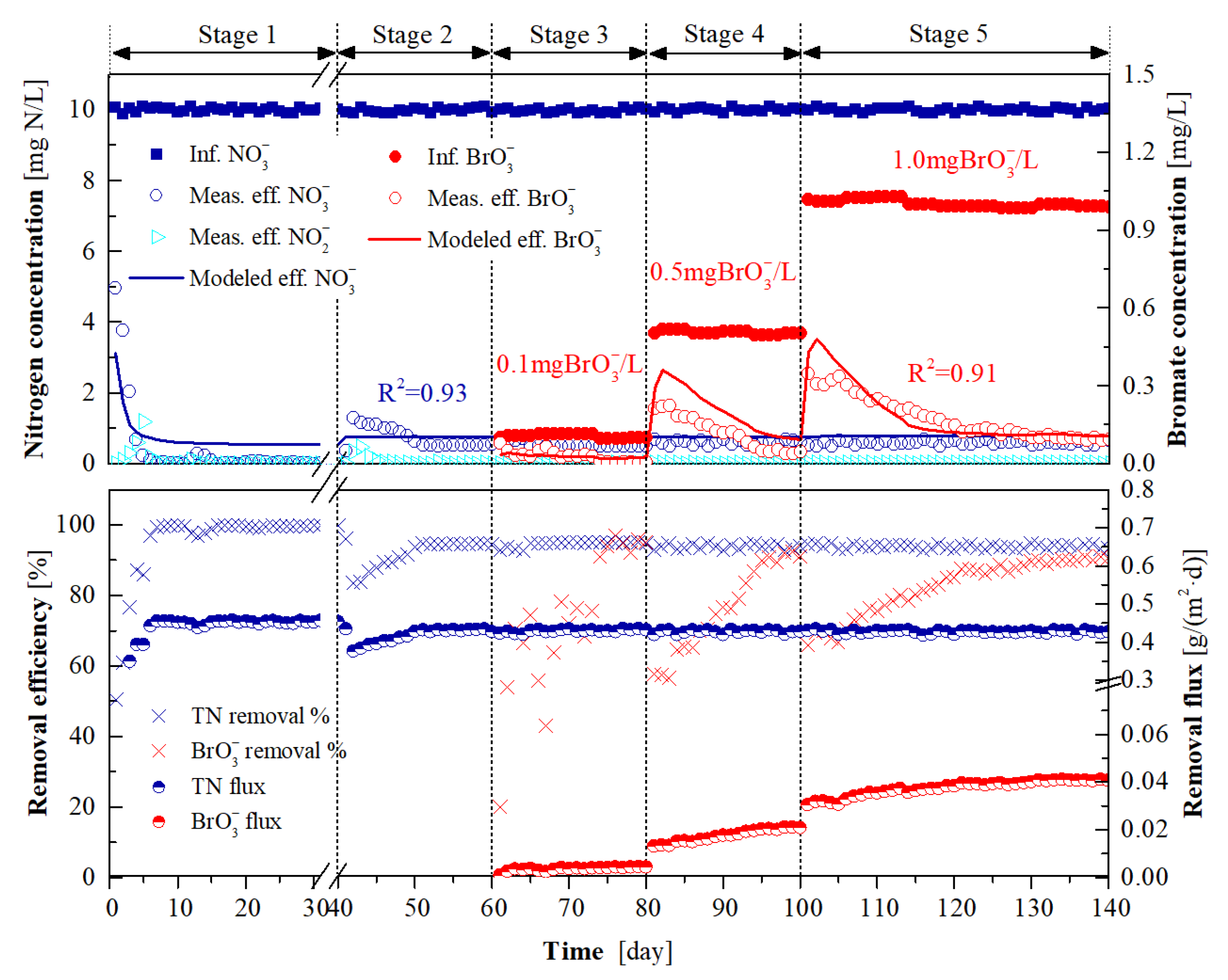
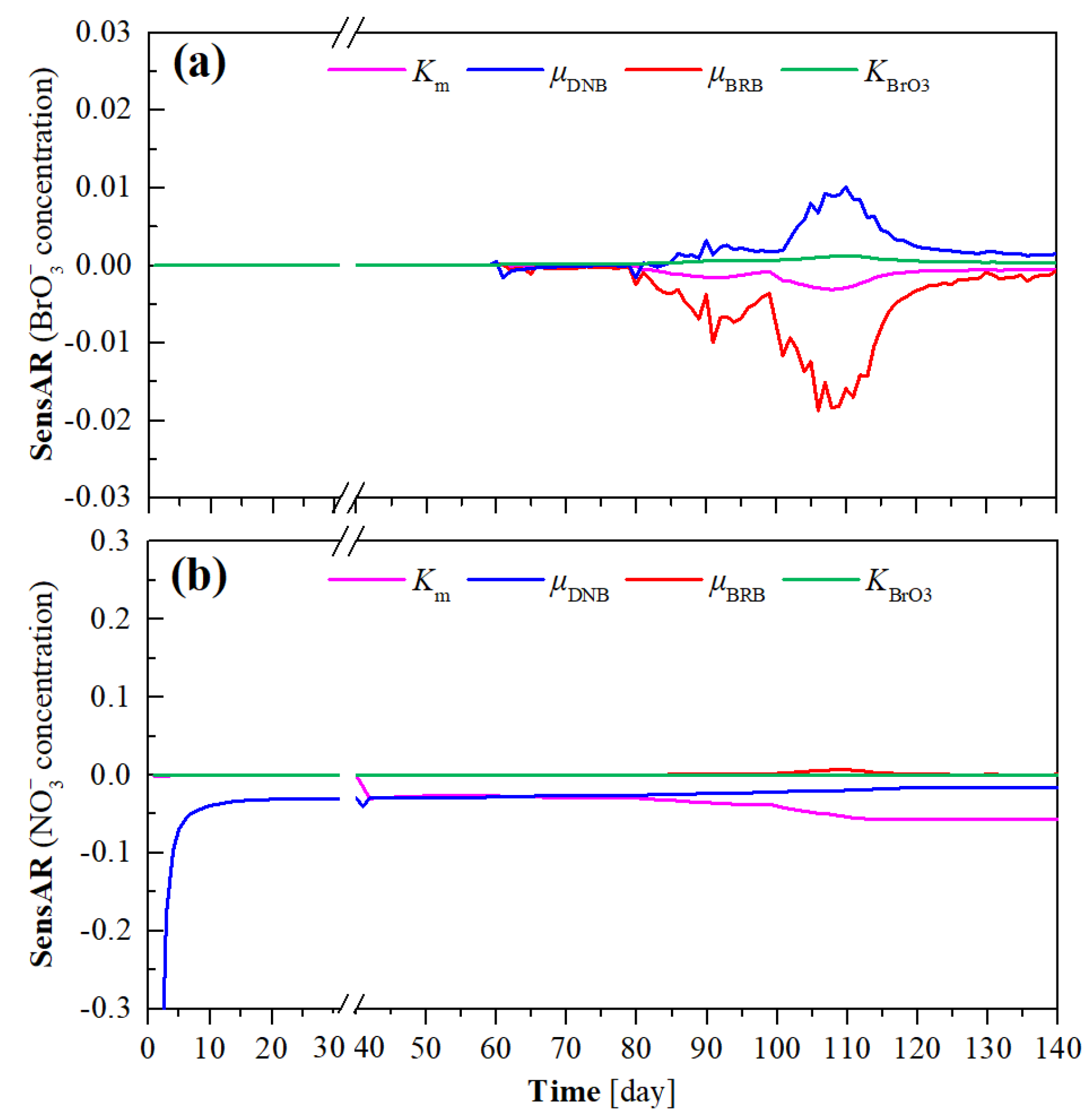
3.1. Long-Term H2-MBfR Performance and Model Calibration
3.2. Short-Term H2-MBfR Performance and Model Validation
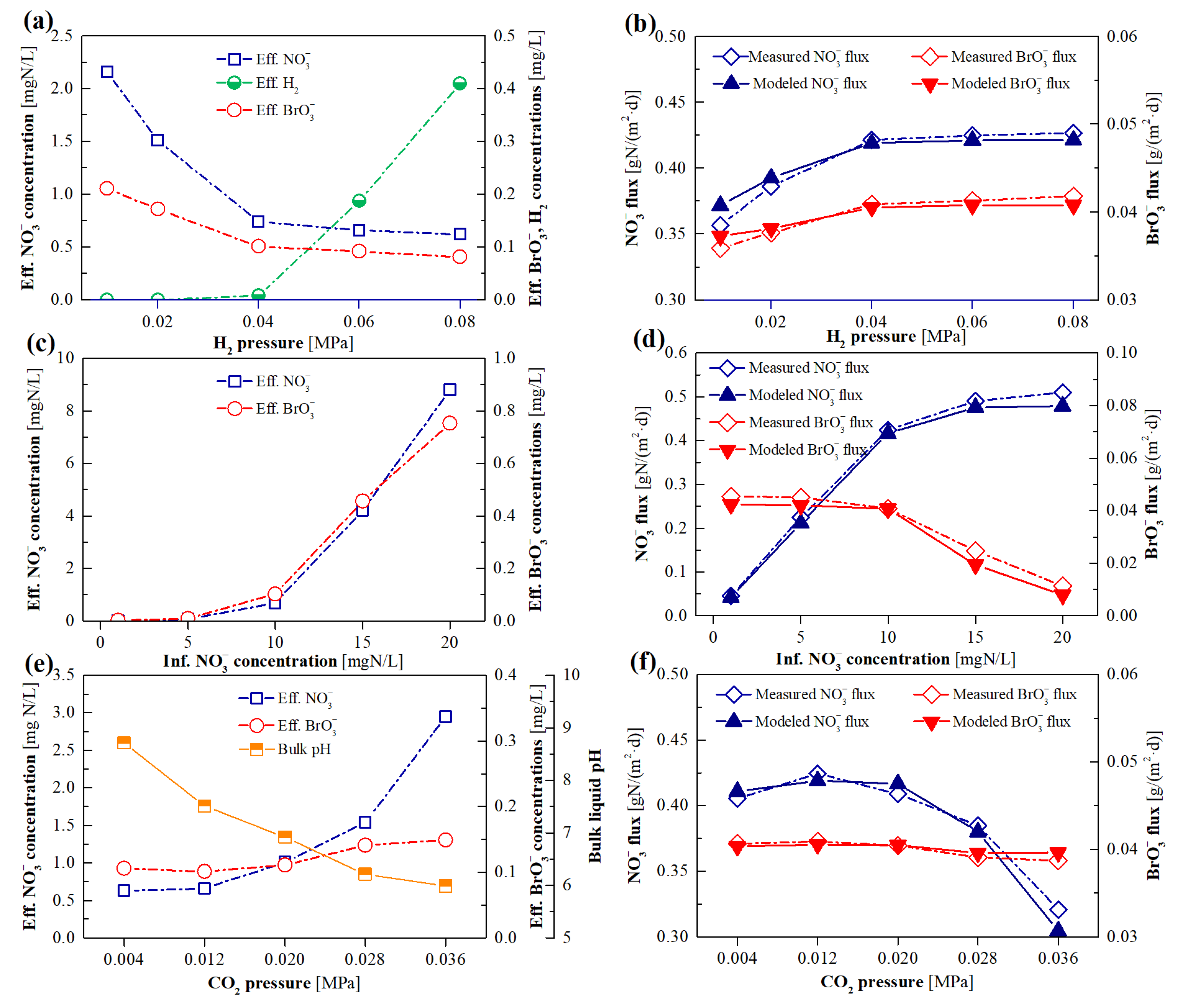
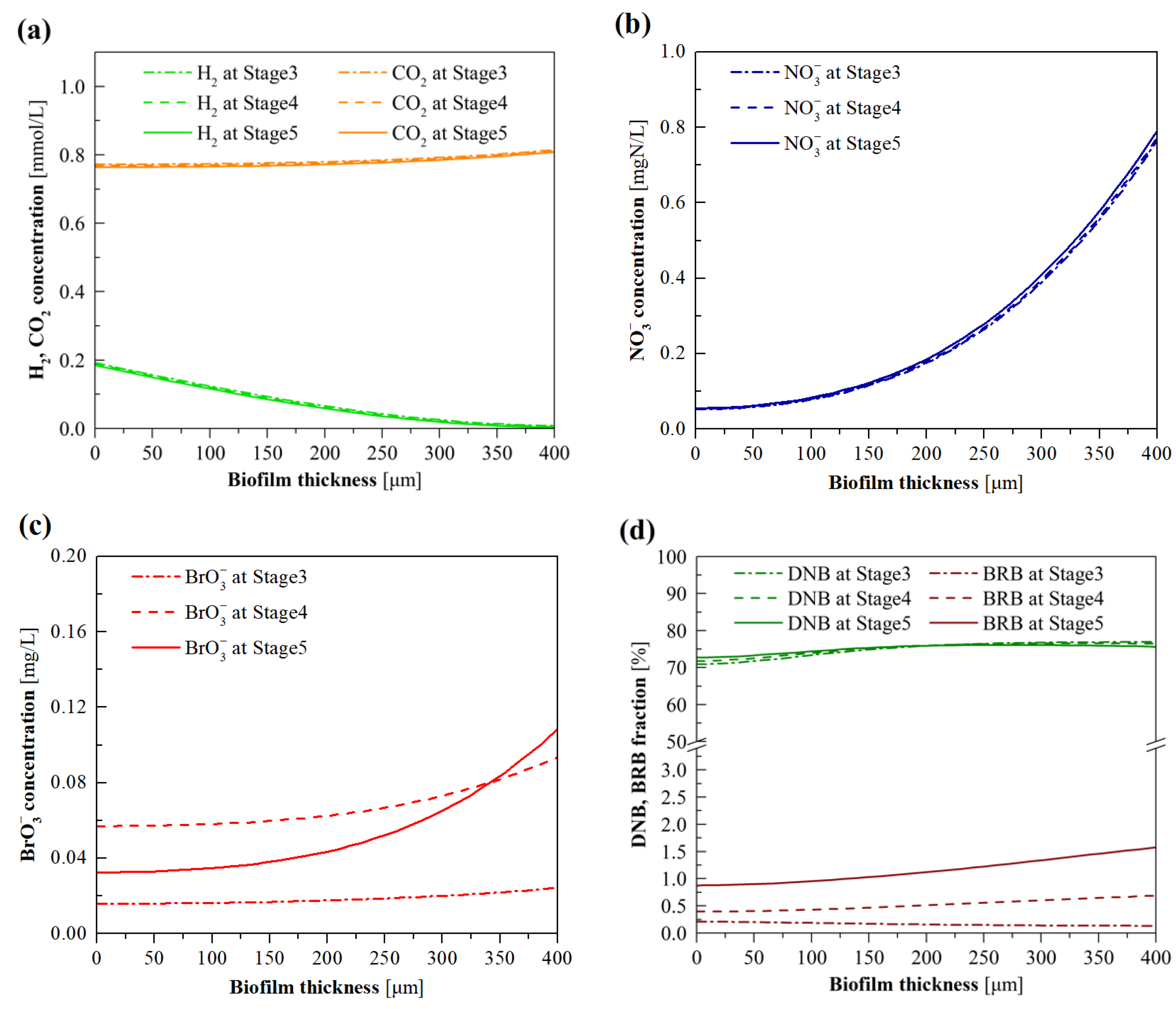
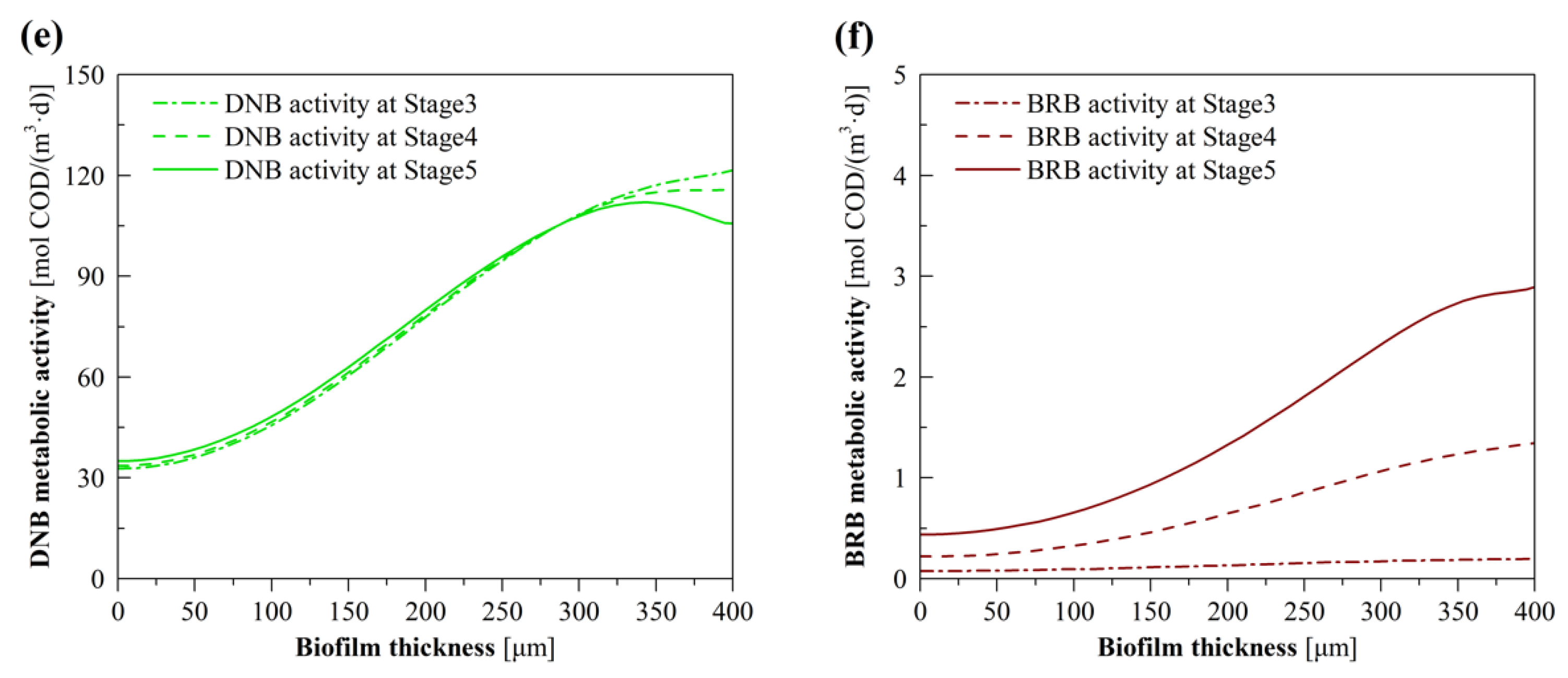
3.3. Model Evaluation of the Effects of BrO3− Loading
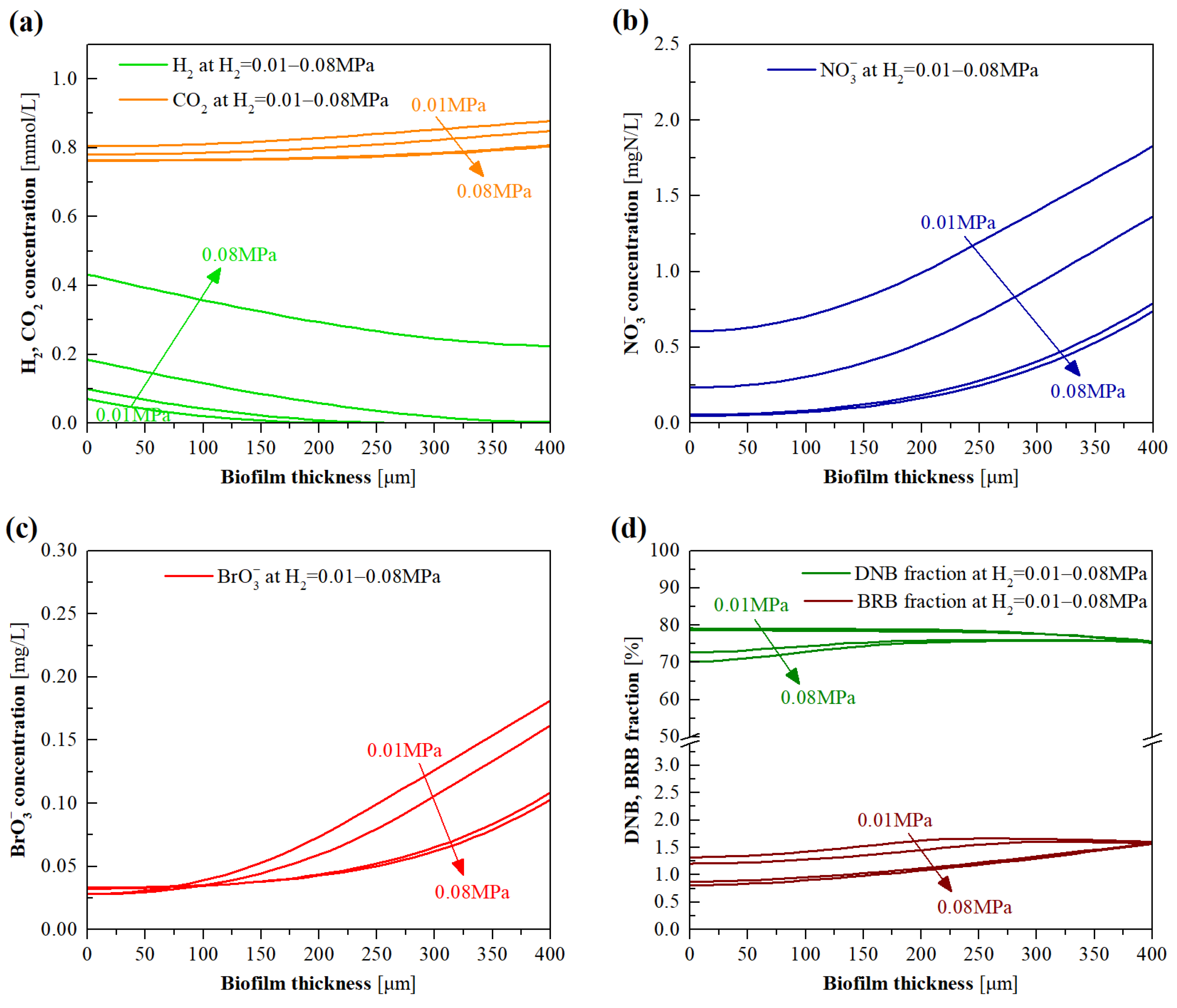
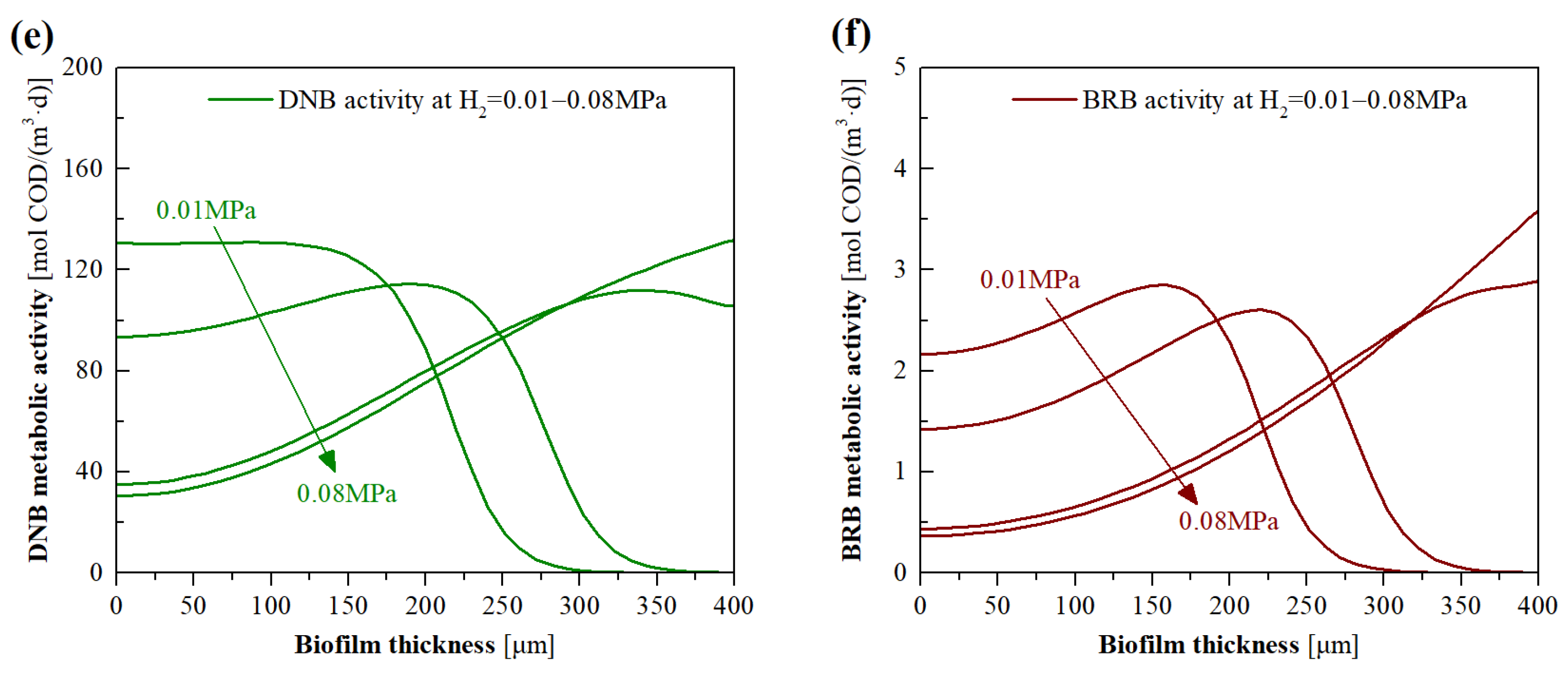
3.4. Model Evaluation of the Effects of H2 Pressure
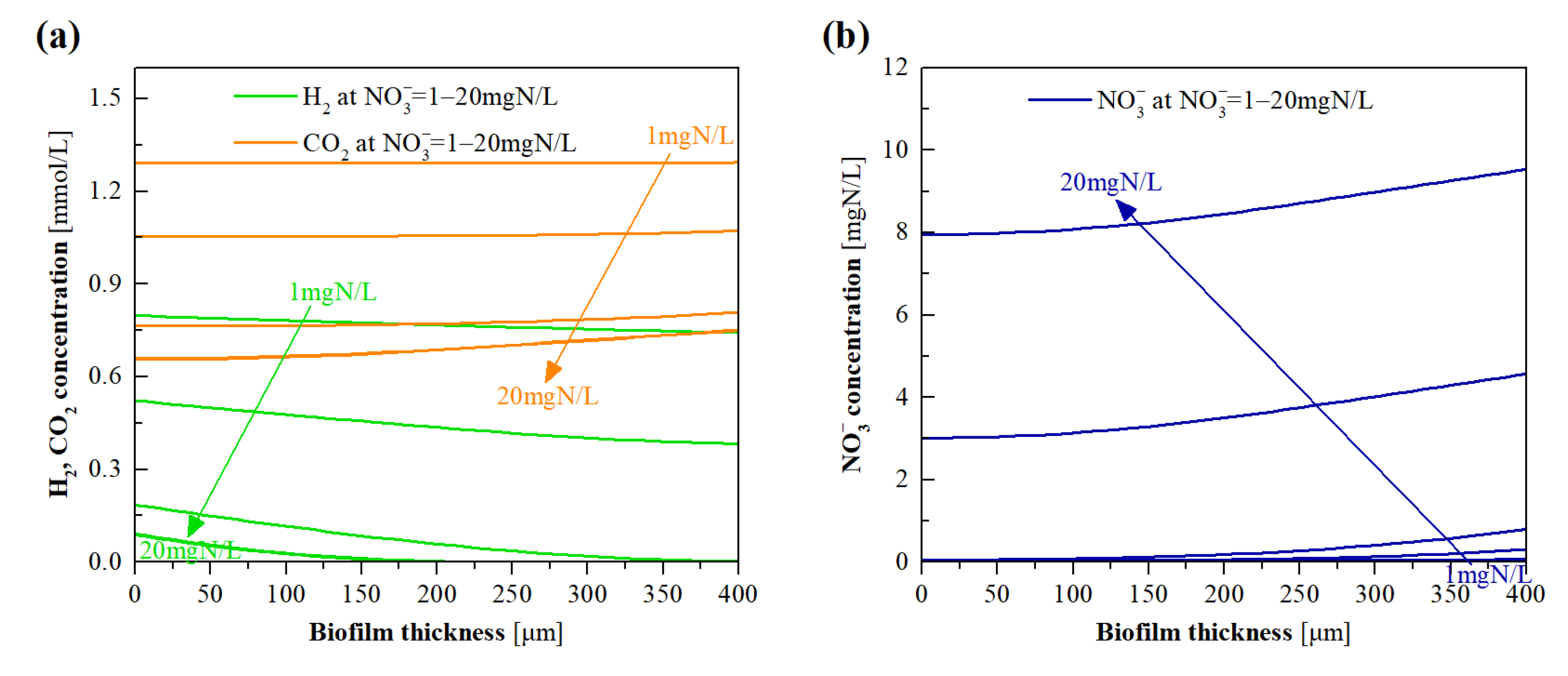
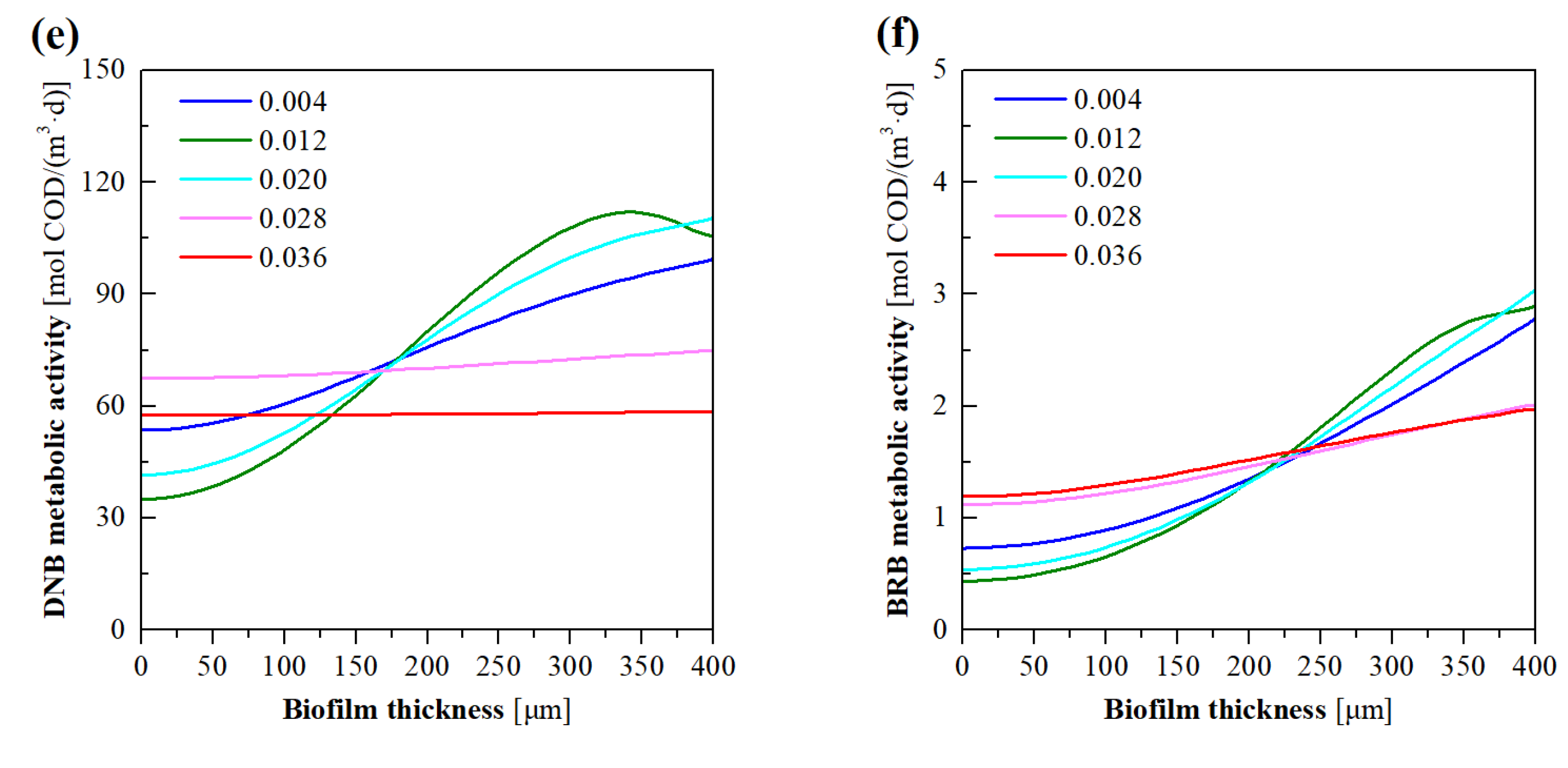
3.5. Model Evaluation of the Effects of NO3− Loading
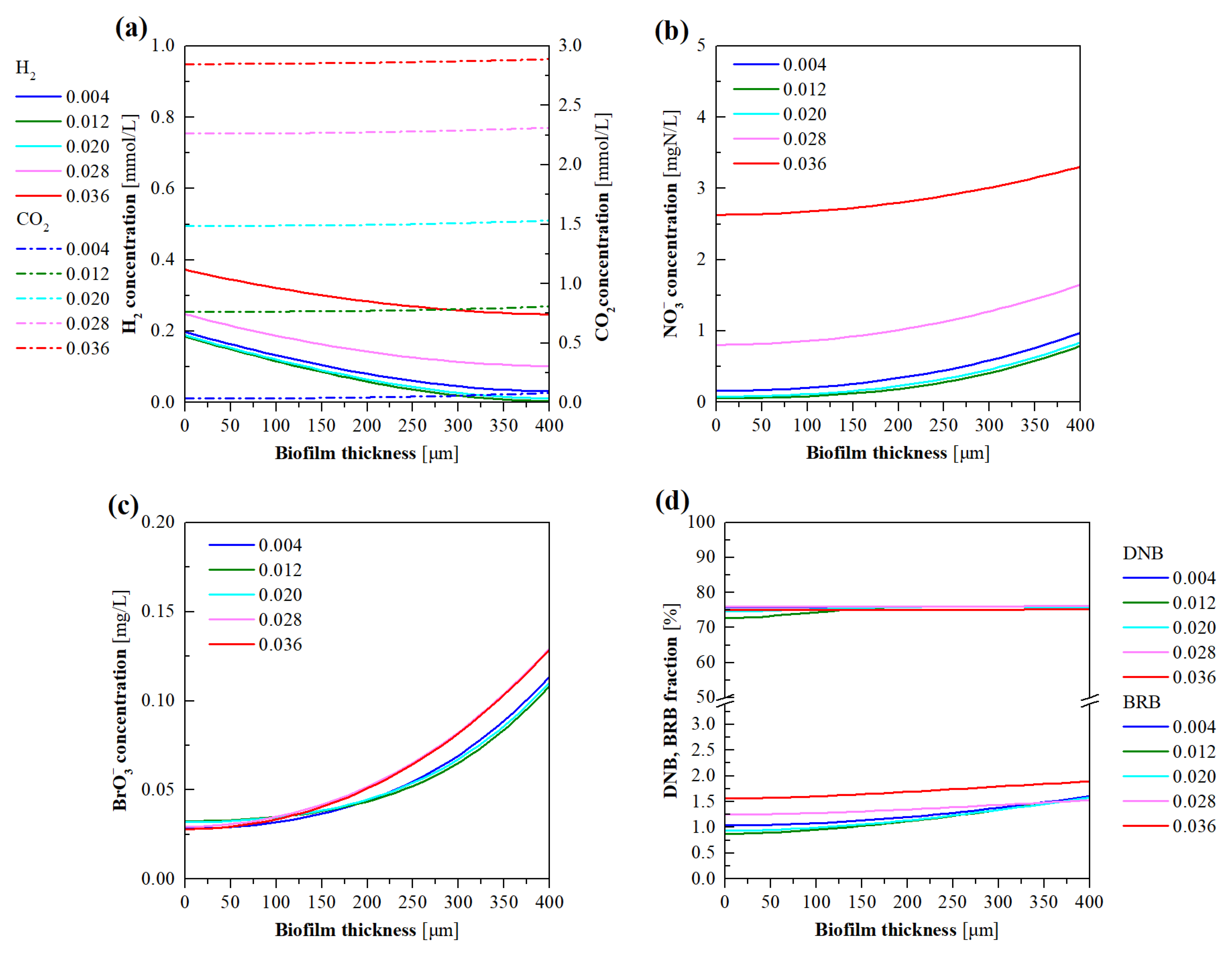
3.6. Model Evaluation of the Effects of CO2 Pressure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Martin, K.J.; Downing, L.S.; Nerenberg, R. Evidence of specialized bromate-reducing bacteria in a hollow fiber membrane biofilm reactor. Water Supply 2009, 8, 473–479. [Google Scholar] [CrossRef]
- Xiao, Q.; Yu, S. Reduction of bromate from drinking water by sulfite/ferric ion systems: Efficacy and mechanisms. J. Hazard. Mater. 2021, 418, 125940. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dong, Z.; Jiang, C.; Wang, C.; Liu, H. An overview of bromate formation in chemical oxidation processes: Occurrence, mechanism, influencing factors, risk assessment, and control strategies. Chemosphere 2019, 237, 124521. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.; Ehrenberg, S.; Godley, A.R.; Lake, R.; Lytton, L.; Cartmell, E. Remediation of bromate-contaminated groundwater in an ex situ fixed-film bioreactor. Sci. Total Environ. 2006, 366, 12–20. [Google Scholar] [CrossRef]
- Jahan, B.N.; Li, L.; Pagilla, K.R. Fate and reduction of bromate formed in advanced water treatment ozonation systems: A critical review. Chemosphere 2021, 266, 128964. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, Q.; Fu, G.; Xu, Y.; Cheng, Y.; Chen, C.; Xiang, R.; Wen, T.; Li, X.; Zeng, G. Denitrifying microbial community with the ability to bromate reduction in a rotating biofilm-electrode reactor. J. Hazard. Mater. 2018, 342, 150–157. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, Y.; Gai, W.-Z.; Deng, Z.-Y. A comparative study on high-efficient reduction of bromate in neutral solution using zero-valent Al treated by different procedures. Sci. Total Environ. 2021, 795, 148786. [Google Scholar] [CrossRef]
- Xia, S.; Liang, J.; Xu, X.; Shen, S. Simultaneous removal of selected oxidized contaminants in groundwater using a continuously stirred hydrogen-based membrane biofilm reactor. J. Environ. Eng. 2013, 25, 96–104. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Peng, L.; Ni, B.-J. Perchlorate, nitrate, and sulfate reduction in hydrogen-based membrane biofilm reactor: Model-based evaluation. Chem. Eng. J. 2017, 316, 82–90. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, Y.; Yuan, Y.; Chen, Y.; Lin, H.; Zheng, J.; Li, H.; Zhang, X. Nitrate removal and dynamics of microbial community of a hydrogen-based membrane biofilm reactor at diverse nitrate loadings and distances from hydrogen supply end. Water 2020, 12, 3196. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Lv, P.-L.; Dong, Q.-Y.; Yeo, S.L.; Rittmann, B.E.; Zhao, H.-P. Bromate and nitrate bioreduction coupled with poly-beta-hydroxybutyrate production in a methane-based membrane biofilm reactor. Environ. Sci. Technol. 2018, 52, 7024–7031. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.J.; Picioreanu, C.; Nerenberg, R. Assessing microbial competition in a hydrogen-based membrane biofilm reactor (MBfR) using multidimensional modeling. Biotechnol. Bioeng. 2015, 112, 1843–1853. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zheng, J.; Perez-Calleja, P.; Picioreanu, C.; Lin, H.; Zhang, X.; Zhang, Y.; Li, H.; Nerenberg, R. New insight into CO2-mediated denitrification process in H2-based membrane biofilm reactor: An experimental and modeling study. Water Res. 2020, 184, 116177. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, H.; Marcus, A.K.; Krajmalnik-Brown, R.; Rittmann, B.E. A steady-state biofilm model for simultaneous reduction of nitrate and perchlorate, part 1: Model development and numerical solution. Environ. Sci. Technol. 2012, 46, 1598–1607. [Google Scholar] [CrossRef]
- Downing, L.S.; Nerenberg, R. Kinetics of microbial bromate reduction in a hydrogen-oxidizing, denitrifying biofilm reactor. Biotechnol. Bioeng. 2007, 98, 543–550. [Google Scholar] [CrossRef]
- Tang, Y.; Zhao, H.; Marcus, A.K.; Krajmalnik-Brown, R.; Rittmann, B.E. A steady-state biofilm model for simultaneous reduction of nitrate and perchlorate, part 2: Parameter optimization and results and discussion. Environ. Sci. Technol. 2012, 46, 1608–1615. [Google Scholar] [CrossRef]
- Li, H.; Zhou, L.; Lin, H.; Zhang, W.; Xia, S. Nitrate Effects on Perchlorate Reduction in a H2/CO2-based Biofilm. Sci. Total Environ. 2019, 694, 133564. [Google Scholar] [CrossRef]
- Lee, K.-C.; Rittmann, B.E. Applying a novel autohydrogenotrophic hollow-fiber membrane biofilm reactor for denitrification of drinking water. Water Res. 2002, 36, 2040–2052. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, C.; Ziv-El, M.; Rittmann, B.E. A pH-control model for heterotrophic and hydrogen-based autotrophic denitrification. Water Res. 2011, 45, 232–240. [Google Scholar] [CrossRef]
- Wu, J.; Yin, Y.; Wang, J. Hydrogen-based membrane biofilm reactors for nitrate removal from water and wastewater. Int. J. Hydrogen Energ. 2018, 43, 1–15. [Google Scholar] [CrossRef]
- Estuardo, C.; Martí, M.C.; Huiliñir, C.; Aspé Lillo, E.; Roeckel von Bennewitz, M. Improvement of nitrate and nitrite reduction rates prediction. Electron. J. Biotechn. 2008, 11, 73–82. [Google Scholar] [CrossRef]
- Vavilin, V.A.; Vasiliev, V.B.; Rytov, S.V.; Ponomarev, A.V. Self-oscillating coexistence of methanogens and sulfate-reducers under hydrogen sulfide inhibition and the pH-regulating effect. Bioresour. Technol. 1994, 49, 105–119. [Google Scholar] [CrossRef]
- Angelidaki, I.; Ellegaard, L.; Ahring, B.K. A mathematical model for dynamic simulation of anaerobic digestion of complex substrates: Focusing on ammonia inhibition. Biotechnol. Bioeng. 1993, 42, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, Y.; Zheng, J.; Li, H.; Ma, J.; Zhang, X.; Wei, Q.; Wang, X.; Zhang, X.; Wang, Z. Mechanistic insights into CO2 pressure regulating microbial competition in a hydrogen-based membrane biofilm reactor for denitrification. Chemosphere 2022, 303, 134875. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Ontiveros-Valencia, A.; Feng, L.; Zhou, C.; Krajmalnik-Brown, R.; Rittmann, B.E. A biofilm model to understand the onset of sulfate reduction in denitrifying membrane biofilm reactors. Biotechnol. Bioeng. 2013, 110, 763–772. [Google Scholar] [CrossRef]
- Lv, X.; Wang, D.; Iqbal, W.; Yang, B.; Mao, Y. Microbial reduction of bromate: Current status and prospects. Biodegradation 2019, 30, 365–374. [Google Scholar] [CrossRef]
- Pang, S.; Rittmann, B.E.; Wu, C.; Yang, L.; Zhou, J.; Xia, S. Synergistic inorganic carbon and denitrification genes contributed to nitrite accumulation in a hydrogen-based membrane biofilm reactor. Bioengineering 2022, 9, 222. [Google Scholar] [CrossRef]
- Ghafari, S.; Hasan, M.; Aroua, M.K. Effect of carbon dioxide and bicarbonate as inorganic carbon sources on growth and adaptation of autohydrogenotrophic denitrifying bacteria. J. Hazard. Mater. 2009, 162, 1507–1513. [Google Scholar] [CrossRef]
- Nerenberg, R. The membrane-biofilm Reactor (MBfR) as a counter-diffusional biofilm process. Curr. Opin. Biotech. 2016, 38, 131–136. [Google Scholar] [CrossRef]
- Wang, F.; van Halem, D.; Ding, L.; Bai, Y.; Lekkerkerker-Teunissen, K.; van der Hoek, J.P. Effective removal of bromate in nitrate-reducing anoxic zones during managed aquifer recharge for drinking water treatment: Laboratory scale simulations. Water Res. 2018, 130, 88–97. [Google Scholar] [CrossRef]
- Xia, S.; Xu, X.; Zhou, C.; Wang, C.; Zhou, L.; Rittmann, B.E. Direct Delivery of CO2 into a Hydrogen-based Membrane Biofilm Reactor and Model Development. Chem. Eng. J. 2016, 290, 154–160. [Google Scholar] [CrossRef]
- Henze, M.; van Loosdrecht, M.C.; Ekama, G.A.; Brdjanovic, D. Biological Wastewater Treatment; IWA Publishing: London, UK, 2008. [Google Scholar]
- Lide, D.R. CRC Handbook of Chemistry and Physics; CRC: Boca Raton, FL, USA, 2012. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component (i) Process (j) | SH2 | SNO3 | SBrO3 | SSMP | SCO2 | XDNB | XBRB | XHB | XIO | XEPS |
|---|---|---|---|---|---|---|---|---|---|---|
| DNB growth | k2 | −k1 | k1 | k3 | ||||||
| BRB growth | k2 | −k1 | k1 | k3 | ||||||
| HB growth | 1 | |||||||||
| DNB decay | −1 | 1−fd | ||||||||
| BRB decay | −1 | 1−fd | ||||||||
| HB Decay | −1 | 1−fd | ||||||||
| Hydrolysis | 1 | −1 | ||||||||
| H2 (mol/m3) | NO3 (mol/m3) | BrO3 (mol/m3) | SMP (mol/m3) | CO2 (mol/m3) | DNB (mol/m3) | BRB (mol/m3) | HB (mol/m3) | IO (mol/m3) | EPS (mol/m3) |
| Process (j) | Kinetic Rate Expressions |
|---|---|
| DNB growth | |
| BRB growth | |
| HB growth | |
| DNB decay | |
| BRB decay | |
| HB Decay | |
| Hydrolysis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Zhang, Y.; Zhang, J.; Dai, X.; Li, H.; Zhang, X.; Wu, Z.; Zheng, J. Model Evaluation of the Microbial Metabolic Processes in a Hydrogen-Based Membrane Biofilm Reactor for Simultaneous Bromate and Nitrate Reduction. Membranes 2022, 12, 774. https://doi.org/10.3390/membranes12080774
Jiang M, Zhang Y, Zhang J, Dai X, Li H, Zhang X, Wu Z, Zheng J. Model Evaluation of the Microbial Metabolic Processes in a Hydrogen-Based Membrane Biofilm Reactor for Simultaneous Bromate and Nitrate Reduction. Membranes. 2022; 12(8):774. https://doi.org/10.3390/membranes12080774
Chicago/Turabian StyleJiang, Minmin, Yuanyuan Zhang, Jie Zhang, Xingru Dai, Haixiang Li, Xuehong Zhang, Zhichao Wu, and Junjian Zheng. 2022. "Model Evaluation of the Microbial Metabolic Processes in a Hydrogen-Based Membrane Biofilm Reactor for Simultaneous Bromate and Nitrate Reduction" Membranes 12, no. 8: 774. https://doi.org/10.3390/membranes12080774