
Inimitable Impacts of Ceramides on Lipid Rafts Formed in Artificial and Natural Cell Membranes


Abstract
:1. Introduction
2. Characteristic Properties of Ceramides and Their Domain Formation in Phospholipid Membranes
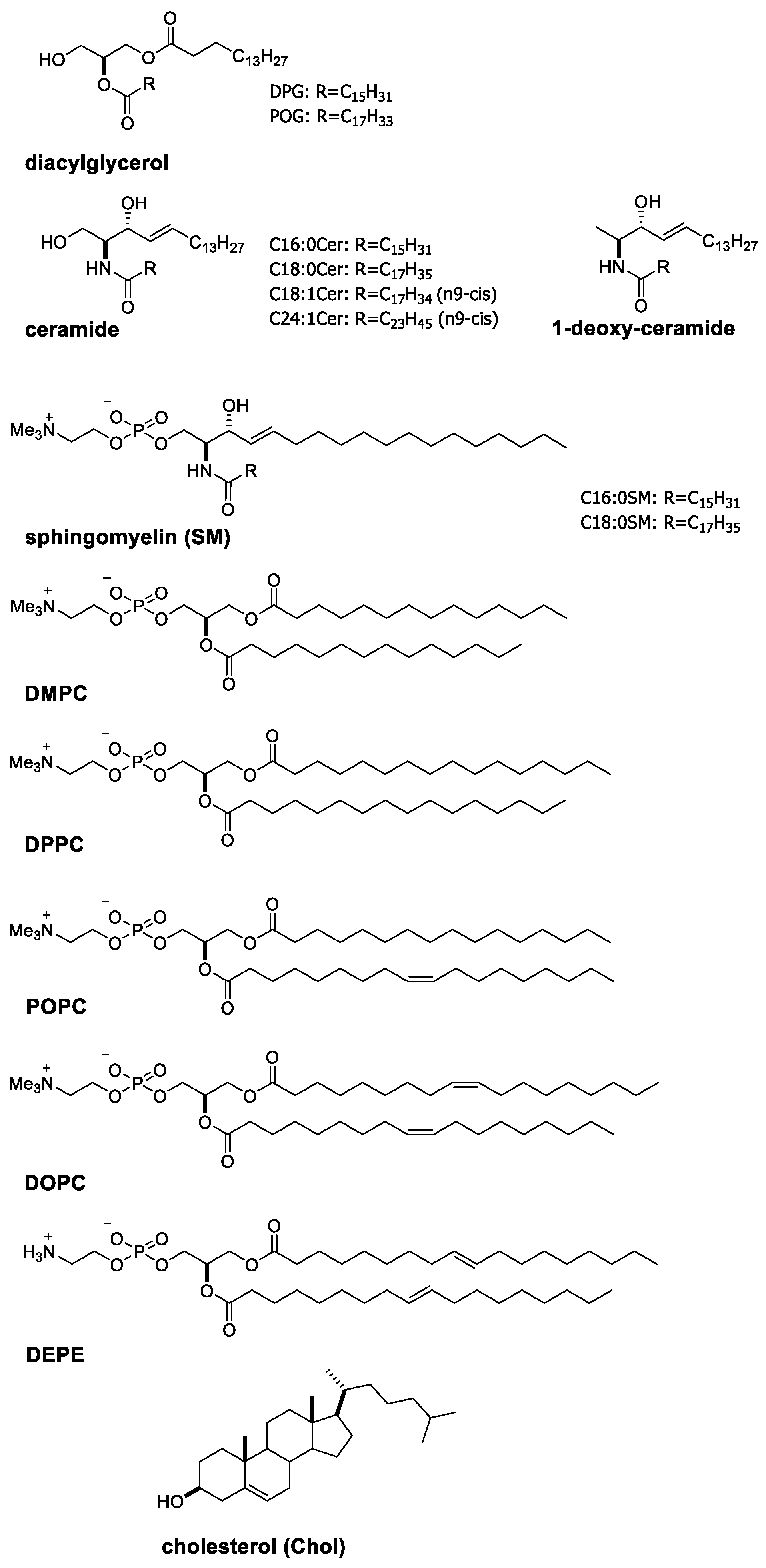
2.1. Characteristic Properties of Ceramide Molecules
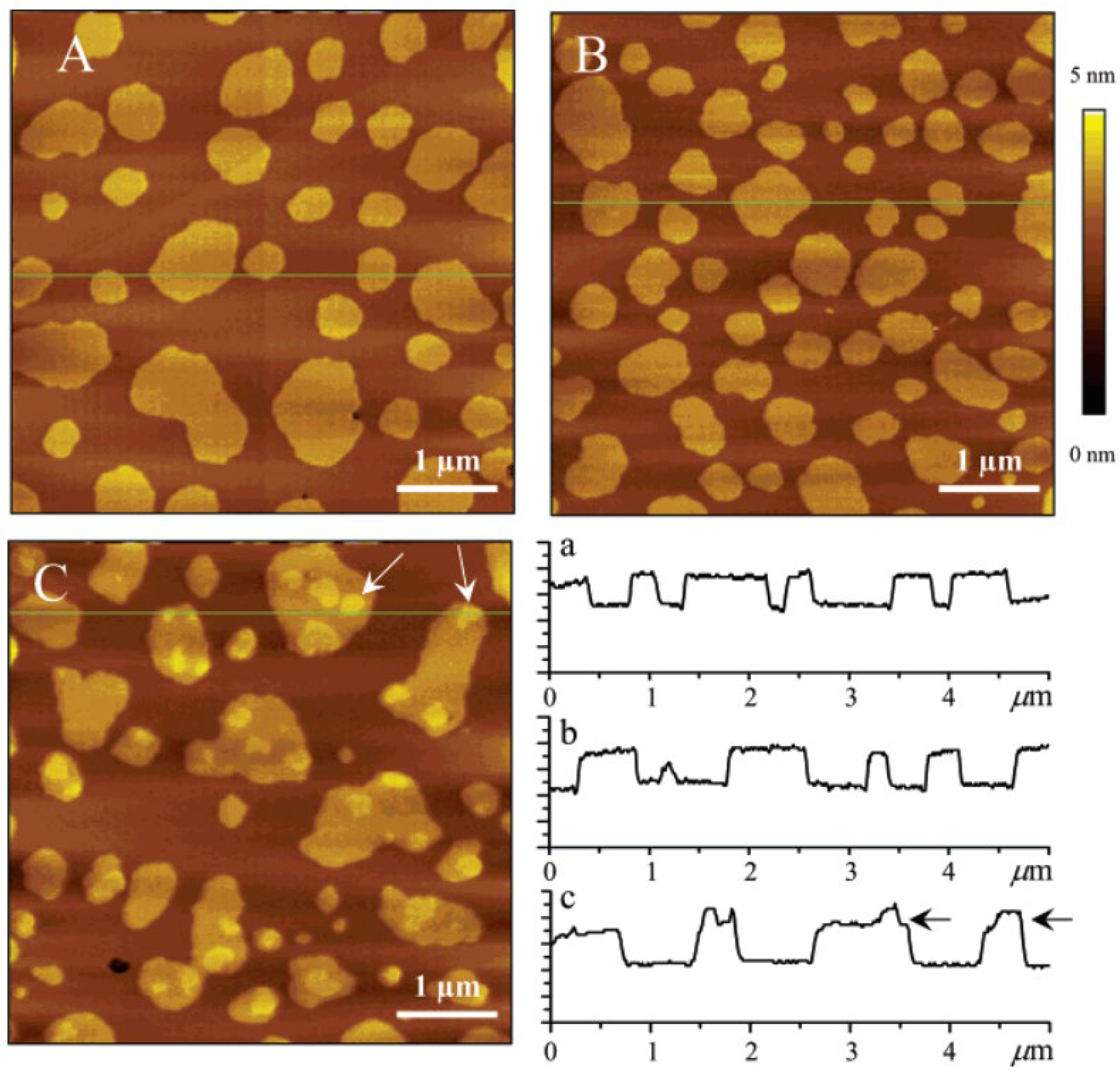
2.2. Formation of Ceramide-Rich Gel Domains in Phospholipid Membranes
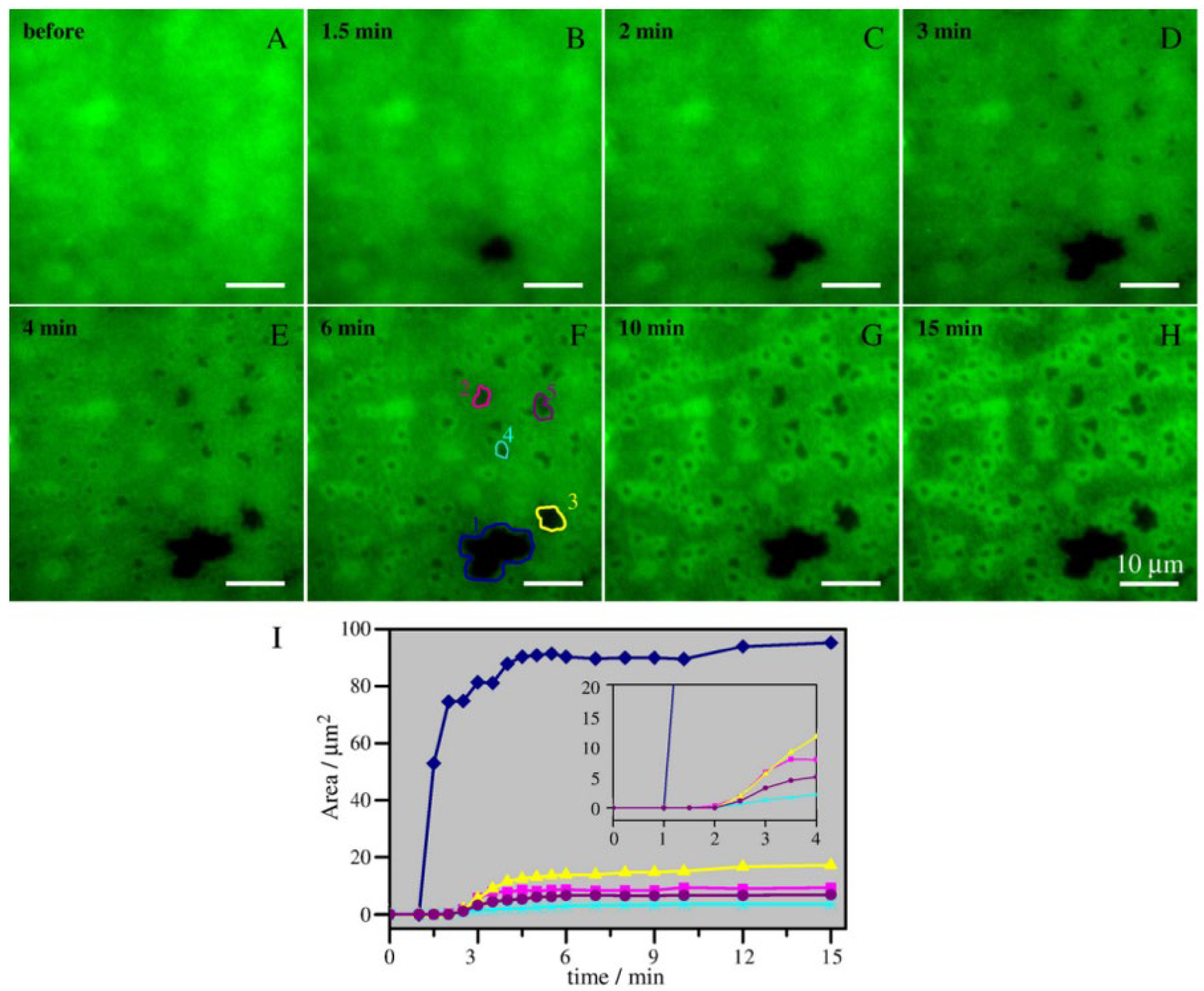
2.3. Ceramide-Induced Fusion of Raft-like Domains
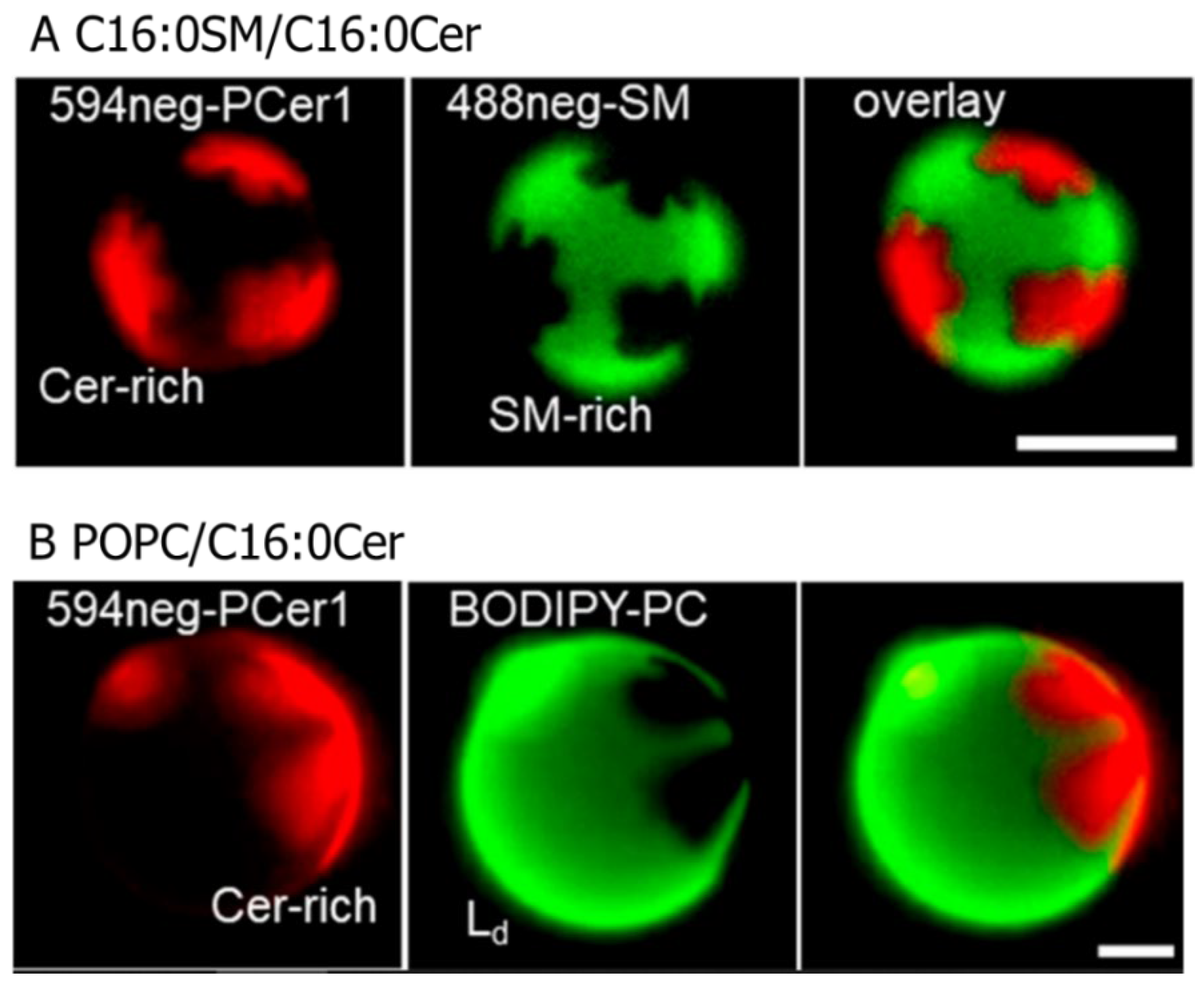
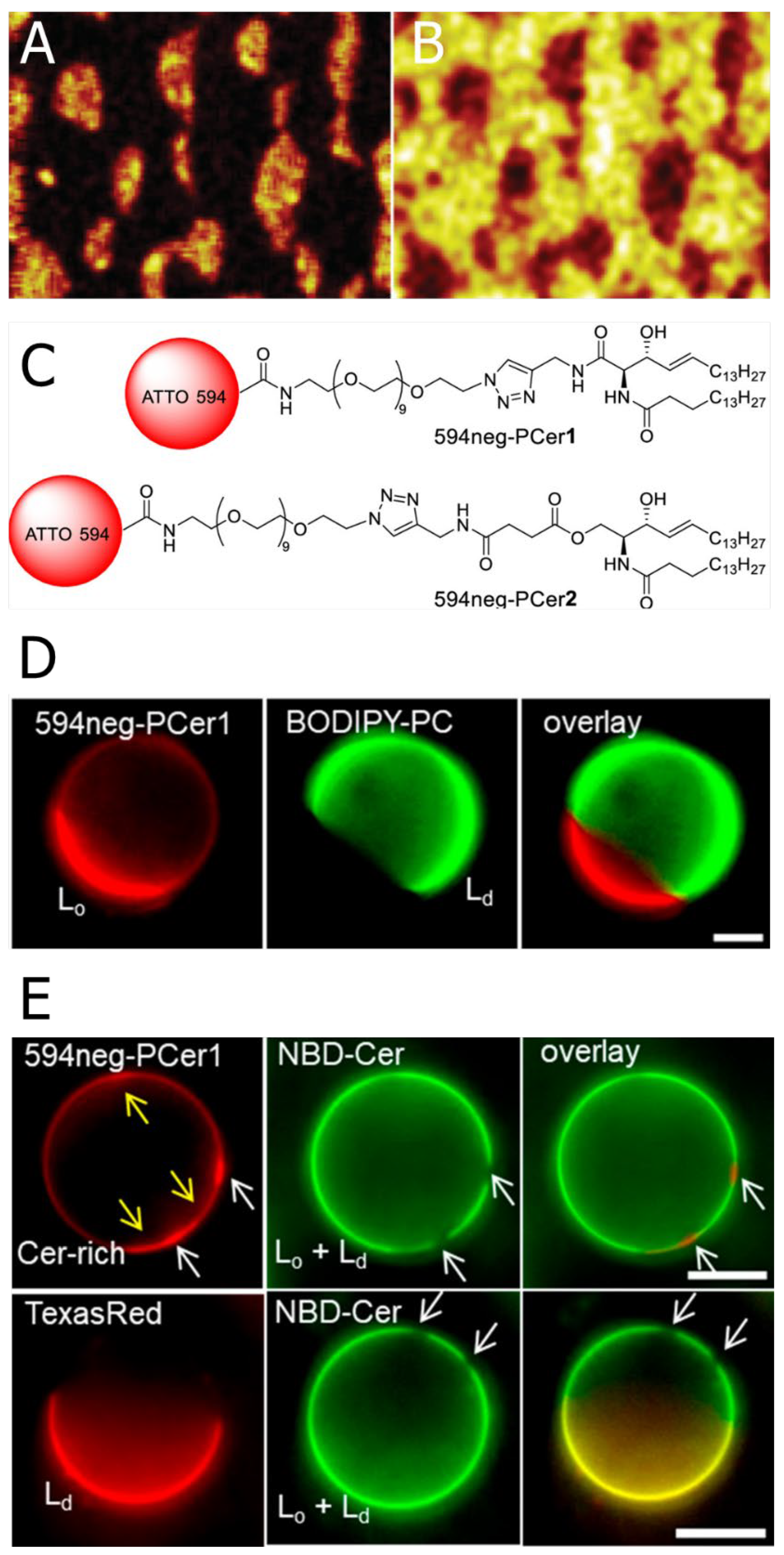
2.4. Devised Methodologies for Visualising Ceramide Distribution
2.5. Ceramide-Induced Compositional Alteration of Raft-like Ordered Domains
3. Formation of Ceramide-Enriched Signal Platforms and Their Biological Functions
3.1. Ceramide-Induced Signal Platform Formation and Transmembrane Signalling
3.2. A Possible Structure of Signal Platforms
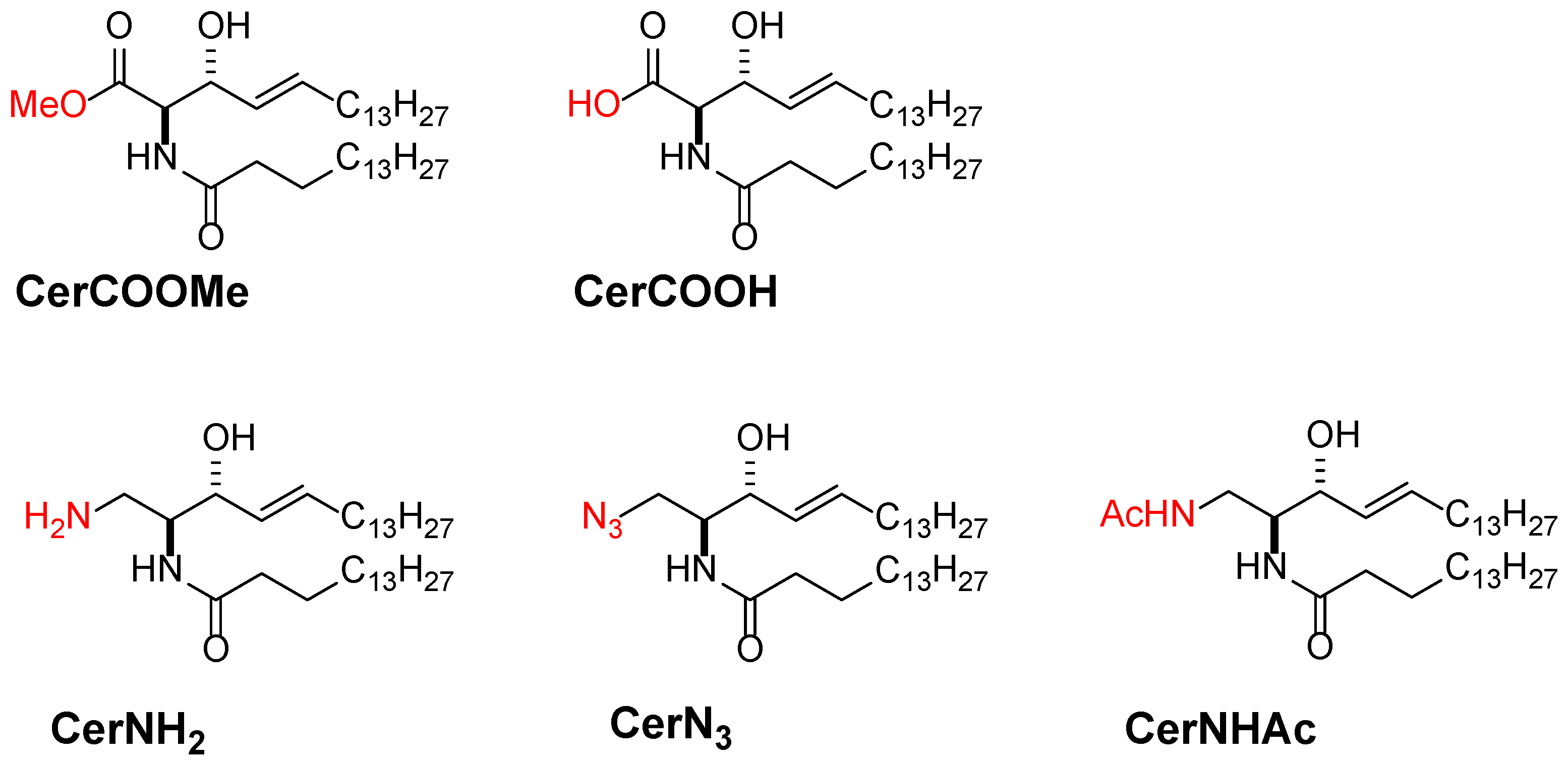
3.3. Promising Ceramide-Analogues for Identifying Intrinsic Ceramide Functions
4. Summary
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shabbir, M.A.; Mehak, F.; Khan, Z.M.; Ahmad, W.; Khan, M.R.; Zia, S.; Rahaman, A.; Aadil, R.M. Interplay between ceramides and phytonutrients: New insights in metabolic syndrome. Trends Food Sci. Technol. 2021, 111, 483–494. [Google Scholar] [CrossRef]
- Skácel, J.; Slusher, B.S.; Tsukamoto, T. Small molecule inhibitors targeting biosynthesis of ceramide, the central hub of the sphingolipid network. J. Med. Chem. 2021, 64, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Pant, D.C.; Aguilera-Albesa, S.; Pujol, A. Ceramide signalling in inherited and multifactorial brain metabolic diseases. Neurobiol. Dis. 2020, 143, 105014. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Okazaki, T. Role of ceramide/sphingomyelin (SM) balance regulated through “SM cycle” in cancer. Cell. Signal. 2021, 87, 110119. [Google Scholar] [CrossRef] [PubMed]
- Stith, J.L.; Velazquez, F.N.; Obeid, L.M. Advances in determining signaling mechanisms of ceramide and role in disease. J. Lipid Res. 2019, 60, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Walchuk, C.; Wang, Y.; Suh, M. The impact of EPA and DHA on ceramide lipotoxicity in the metabolic syndrome. Br. J. Nutr. 2021, 125, 863–875. [Google Scholar] [CrossRef]
- O’Brien, J.S.; Rouser, G. The fatty acid composition of brain sphingolipids: Sphingomyelin, ceramide, cerebroside, and cerebroside sulfate. J. Lipid Res. 1964, 5, 339–3420. [Google Scholar] [CrossRef]
- Akino, T. Sphingosine base and fatty acid compositions of pig brain sphingolipids. Tohoku J. Exp. Med. 1969, 98, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Summers, S.A.; Garza, L.A.; Zhou, H.; Birnbaum, M.J. Regulation of insulin-stimulated glucose transporter GLUT4 translocation and Akt kinase activity by ceramide. Mol. Cell. Biol. 1998, 18, 5457–5464. [Google Scholar] [CrossRef] [Green Version]
- Straczkowski, M.; Kowalska, I.; Baranowski, M.; Nikolajuk, A.; Otziomek, E.; Zabielski, P.; Adamska, A.; Blachnio, A.; Gorski, J.; Gorska, M. Increased skeletal muscle ceramide level in men at risk of developing type 2 diabetes. Diabetologia 2007, 50, 2366–2373. [Google Scholar] [CrossRef] [Green Version]
- Masamune, A.; Igarashi, Y.; Hakomori, S.I. Regulatory role of ceramide in interleukin (IL)-1Β-induced E-selectin expression in human umbilical vein endothelial cells: Ceramide enhances IL-1Β action, but is not sufficient for E-selectin expression. J. Biol. Chem. 1996, 271, 9368–9375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, K.; Dixit, V.M. Ceramide in apoptosis-Does it really matter? Trends Biochem. Sci. 1998, 23, 374–377. [Google Scholar] [CrossRef]
- Jin, Z.H.; Li, W.; Pan, H.Z. Ceramide and apoptosis. Prog. Biochem. Biophys. 1999, 26, 540–541. [Google Scholar]
- Obeid, L.M.; Linardic, C.M.; Karolak, L.A.; Hannun, Y.A. Programmed cell death induced by ceramide. Science 1993, 259, 1769–1771. [Google Scholar] [CrossRef] [PubMed]
- Bielawska, A.; Linardic, C.M.; Hannun, Y.A. Modulation of cell growth and differentiation by ceramide. FEBS Lett. 1992, 307, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Jayadev, S.; Liu, B.; Bielawska, A.E.; Lee, J.Y.; Nazaire, F.; Pushkareva, M.Y.; Obeid, L.M.; Hannun, Y.A. Role for ceramide in cell cycle arrest. J. Biol. Chem. 1995, 270, 2047–2052. [Google Scholar] [CrossRef] [Green Version]
- Venable, M.E.; Lee, J.Y.; Smyth, M.J.; Bielawska, A.; Obeid, L.M. Role of ceramide in cellular senescence. J. Biol. Chem. 1995, 270, 30701–30708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotte, P.J. Molecular properties of various structurally defined sphingomyelins-correlation of structure with function. Prog. Lipid Res. 2013, 52, 206–219. [Google Scholar] [CrossRef]
- Slotte, J.P. The Importance of hydrogen bonding in sphingomyelin’s membrane interactions with co-Lipids. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 304–310. [Google Scholar] [CrossRef]
- Cebecauer, M.; Amaro, M.; Jurkiewicz, P.; Sarmento, M.J.; Šachl, R.; Cwiklik, L.; Hof, M. Membrane lipid nanodomains. Chem. Rev. 2018, 118, 11259–11297. [Google Scholar] [CrossRef]
- Bieberich, E. Sphingolipids and lipid rafts: Novel concepts and methods of analysis. Chem. Phys. Lipids 2018, 216, 114–131. [Google Scholar] [CrossRef] [PubMed]
- Kraft, M.L. Plasma membrane organization and function: Moving past lipid rafts. Mol. Biol. Cell 2013, 24, 2765–2768. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Sampaio, J.L. Membrane organization and lipid rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Codini, M.; Garcia-Gil, M.; Albi, E. Cholesterol and sphingolipid enriched lipid rafts as therapeutic targets in cancer. Int. J. Mol. Sci. 2021, 22, 726. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.J. Sphingolipid symmetry governs membrane lipid raft structure. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 1922–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, J.; Atienza, J.M.; Duclos, R.I.; Rawlings, A.V.; Dong, Z.; Shipley, G.G. Structural and thermotropic properties of synthetic C16:0 (palmitoyl) ceramide: Effect of hydration. J. Lipid Res. 1995, 36, 1936–1944. [Google Scholar] [CrossRef]
- Souza, S.L.; Capitán, M.J.; Álvarez, J.; Funari, S.S.; Lameiro, M.H.; Melo, E. Phase behavior of aqueous dispersions of mixtures of N-palmitoyl ceramide and cholesterol: A lipid system with ceramide-cholesterol crystalline lamellar phases. J. Phys. Chem. B 2009, 113, 1367–1375. [Google Scholar] [CrossRef]
- López-García, F.; Villalaín, J.; Gómez-Fernández, J.C.; Quinn, P.J. The phase behavior of mixed aqueous dispersions of dipalmitoyl derivatives of phosphatidylcholine and diacylglycerol. Biophys. J. 1994, 66, 1991–2004. [Google Scholar] [CrossRef] [Green Version]
- Ekman, P.; Maula, T.; Yamaguchi, S.; Yamamoto, T.; Nyholm, T.K.M.; Katsumura, S.; Slotte, J.P. Formation of an ordered phase by ceramides and diacylglycerols in a fluid phosphatidylcholine bilayer-correlation with structure and hydrogen bonding capacity. Biochim. Biophys. Acta-Biomembr. 2015, 1848, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Monreal, A.M.; Villalaín, J.; Aranda, F.J.; Gómez-Fernández, J.C. The phase behavior of aqueous dispersions of unsaturated mixtures of diacylglycerols and phospholipids. Biochim. Biophys. Acta-Biomembr. 1998, 1373, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Gulbins, E.; Kolesnick, R. Raft Ceramide in molecular medicine. Oncogene 2003, 22, 7070–7077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascher, I. Molecular arrangements in sphingolipids conformation and hydrogen bonding of ceramide and their implication on membrane stability and permeability. Biochim. Biophys. Acta-Biomembr. 1976, 455, 433–451. [Google Scholar] [CrossRef]
- Siskind, L.J.; Davoody, A.; Lewin, N.; Marshall, S.; Colombini, M. Enlargement and contracture of C2-ceramide channels. Biophys. J. 2003, 85, 1560–1575. [Google Scholar] [CrossRef] [Green Version]
- Kinnunen, P.K.J.; Holopainen, J.M. Sphingomyelinase activity of LDLA link between atherosclerosis, ceramide, and apoptosis? Trends Cardiovasc. Med. 2002, 12, 37–42. [Google Scholar] [CrossRef]
- Jiménez-Rojo, N.; Sot, J.; Busto, J.V.; Shaw, W.A.; Duan, J.; Merrill, A.H.; Alonso, A.; Goñi, F.M. Biophysical properties of novel 1-deoxy-(dihydro)ceramides occurring in mammalian cells. Biophys. J. 2014, 107, 2850–2859. [Google Scholar] [CrossRef] [Green Version]
- Garidel, P.; Fölting, B.; Schaller, I.; Kerth, A. The microstructure of the stratum corneum lipid barrier: Mid-infrared spectroscopic studies of hydrated ceramide:palmitic acid:cholesterol model systems. Biophys. Chem. 2010, 150, 144–156. [Google Scholar] [CrossRef]
- Moore, D.J.; Rerek, M.E.; Mendelsohn, R. FTIR spectroscopy studies of the conformational order and phase behavior of ceramides. J. Phys. Chem. B 1997, 101, 8933–8940. [Google Scholar] [CrossRef]
- Notman, R.; Den Otter, W.K.; Noro, M.G.; Briels, W.J.; Anwar, J. The permeability enhancing mechanism of DMSO in ceramide bilayers simulated by molecular dynamics. Biophys. J. 2007, 93, 2056–2068. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Klauda, J.B. Molecular dynamics simulations of ceramide and ceramide-phosphatidylcholine Bilayers. J. Phys. Chem. B 2017, 121, 10091–10104. [Google Scholar] [CrossRef]
- Pandit, S.A.; Scott, H.L. Molecular-dynamics simulation of a ceramide bilayer. J. Chem. Phys. 2006, 124, 014708. [Google Scholar] [CrossRef]
- Matsufuji, T.; Kinoshita, M.; Matsumori, N. Preparation and membrane distribution of fluorescent derivatives of ceramide. Langmuir 2019, 35, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Möuts, A.; Vattulainen, E.; Matsufuji, T.; Kinoshita, M.; Matsumori, N.; Slotte, J.P. On the importance of the C(1)-OH and C(3)-OH functional groups of the long-chain base of ceramide for interlipid interaction and lateral segregation into ceramide-rich domains. Langmuir 2018, 34, 15864–15870. [Google Scholar] [CrossRef] [PubMed]
- Pinto, S.N.; Silva, L.C.; Futerman, A.H.; Prieto, M. Effect of ceramide structure on membrane biophysical properties: The role of acyl chain length and unsaturation. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 2753–2760. [Google Scholar] [CrossRef] [Green Version]
- Holopainen, J.M.; Lehtonen, J.Y.A.; Kinnunen, P.K.J. Lipid microdomains in dimyristoylphosphatidylcholine–ceramide liposomes. Chem. Phys. Lipids 1997, 88, 1–13. [Google Scholar] [CrossRef]
- Veiga, M.P.; Arrondo, J.L.R.; Goñi, F.M.; Alonso, A. Ceramides in phospholipid membranes: Effects on bilayer stability and transition to nonlamellar phases. Biophys. J. 1999, 76, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Sot, J.; Aranda, F.J.; Collado, M.I.; Goñi, F.M.; Alonso, A. Different effects of long- and short-chain ceramides on the gel-fluid and lamellar-hexagonal transitions of phospholipids: A calorimetric, NMR, and x-ray diffraction study. Biophys. J. 2005, 88, 3368–3380. [Google Scholar] [CrossRef] [Green Version]
- López-Montero, I.; Monroy, F.; Vélez, M.; Devaux, P.F. Ceramide: From lateral segregation to mechanical stress. Biochim. Biophys. Acta-Biomembr. 2010, 1798, 1348–1356. [Google Scholar] [CrossRef] [Green Version]
- Catapano, E.R.; Arriaga, L.R.; Espinosa, G.; Monroy, F.; Langevin, D.; López-Montero, I. Solid character of membrane ceramides: A surface rheology study of their mixtures with sphingomyelin. Biophys. J. 2011, 10, 2721–2730. [Google Scholar] [CrossRef] [Green Version]
- López-Montero, I.; Catapano, E.R.; Espinosa, G.; Arriaga, L.R.; Langevin, D.; Monroy, F. Shear and compression rheology of Langmuir monolayers of natural ceramides: Solid character and plasticity. Langmuir 2013, 29, 6634–6644. [Google Scholar] [CrossRef]
- Catapano, E.R.; Natale, P.; Monroy, F.; López-Montero, I. The enzymatic sphingomyelin to ceramide conversion increases the shear membrane viscosity at the air-water interface. Adv. Coll. Interf. Sci. 2017, 247, 555–560. [Google Scholar] [CrossRef]
- Ipsen, J.H.; Karlström, G.; Mourtisen, O.G.; Wennerström, H.; Zuckermann, M.J. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta-Biomembr. 1987, 905, 162–172. [Google Scholar] [CrossRef]
- Quinn, P.J.; Wolf, C. The liquid-ordered phase in membranes. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chachaty, C.; Rainteau, D.; Tessier, C.; Quinn, P.J.; Wolf, C. Building up of the liquid-ordered phase formed by sphingomyelin and cholesterol. Biophys. J. 2005, 88, 4032–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, M.; Tanaka, K.; Matsumori, N. The influence of ceramide and its dihydro analog on the physico-chemical properties of sphingomyelin bilayers. Chem. Phys. Lipids 2020, 226, 104835. [Google Scholar] [CrossRef]
- Maulik, P.R.; Shipley, G.G. Interaction of N-steraroyl sphingomyelin with cholesterol and dipalmitoylphosphatidylcholine in bilayer membranes. Biophys. J. 1996, 760, 2256–2265. [Google Scholar] [CrossRef] [Green Version]
- Siavashi, R.; Phaterpekar, T.; Leung, S.S.W.; Alonso, A.; Goñi, F.M.; Thewalt, J.L. Lamellar phases composed of phospholipid, cholesterol, and ceramide, as studied by 2H NMR. Biophys. J. 2019, 117, 296–306. [Google Scholar] [CrossRef]
- Murthy, A.V.R.; Guyomarc’h, F.; Lopez, C. Palmitoyl ceramide promotes milk sphingomyelin gel phase domains formation and affects the mechanical properties of the fluid phase in milk-SM/DOPC supported membranes. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 635–644. [Google Scholar] [CrossRef]
- Ira; Johnston, L.J. Sphingomyelinase generation of ceramide promotes clustering of nanoscale domains in supported bilayer membranes. Biochim. Biophys. Acta-Biomembr. 2008, 1778, 185–197. [Google Scholar]
- Silva, L.C.; De Almeida, R.F.M.; Castro, B.M.; Fedorov, A.; Prieto, M. Ceramide-domain formation and collapse in lipid rafts: Membrane reorganization by an apoptotic lipid. Biophys. J. 2007, 92, 502–516. [Google Scholar] [CrossRef] [Green Version]
- Sot, J.; Ibarguren, M.; Busto, J.V.; Montes, L.R.; Goñi, F.M.; Alonso, A. Cholesterol displacement by ceramide in sphingomyelin-containing liquid-ordered domains, and generation of gel regions in giant lipidic vesicles. FEBS Lett. 2008, 582, 3230–3236. [Google Scholar] [CrossRef] [Green Version]
- Ira; Linda, J.J. Ceramide promotes restructuring of model raft membranes. Langmuir 2006, 22, 11284–11289. [Google Scholar] [CrossRef] [PubMed]
- Chiantia, S.; Kahya, N.; Ries, J.; Schwille, P. Effects of ceramide on liquid-ordered domains investigated by simultaneous AFM and FCS. Biophys. J. 2006, 90, 4500–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, L.J. Nanoscale imaging of domains in supported lipid membranes. Langmuir 2007, 23, 5886–5895. [Google Scholar] [CrossRef] [PubMed]
- Canals, D.; Salamone, S.; Hannun, Y.A. Visualizing bioactive ceramides. Chem. Phys. Lipids 2018, 216, 142–151. [Google Scholar] [CrossRef]
- Popov, J.; Vobornik, D.; Coban, O.; Keating, E.; Miller, D.; Francis, J.; Petersen, N.O.; Johnston, L.J. Chemical mapping of ceramide distribution in sphingomyelin-rich domains in monolayers. Langmuir 2008, 24, 13502–13508. [Google Scholar] [CrossRef]
- Zheng, L.; McQuaw, C.M.; Ewing, A.G.; Winograd, N. Sphingomyelin/phosphatidylcholine and cholesterol interactions studied by imaging mass spectrometry. J. Am. Chem. Soc. 2007, 129, 15730–15731. [Google Scholar] [CrossRef] [Green Version]
- McQuaw, C.M.; Sostarecz, A.G.; Zheng, L.; Ewing, A.G.; Winograd, N. Lateral heterogeneity of dipalmitoylphosphatidylethanolamine-cholesterol Langmuir-Blodgett films investigated with imaging time-of-flight secondary ion mass spectrometry and atomic force microscopy. Langmuir 2005, 21, 807–813. [Google Scholar] [CrossRef]
- Kinoshita, M.; Suzuki, K.G.N.; Murata, M.; Matsumori, N. Evidence of lipid rafts based on the partition and dynamic behavior of sphingomyelins. Chem. Phys. Lipids 2018, 215, 84–95. [Google Scholar] [CrossRef]
- Passarelli, M.K.; Winograd, N. Lipid imaging with time-of-flight secondary ion mass spectrometry (ToF-SIMS). Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2015, 1811, 976–990. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.S. Latest applications of 3D ToF-SIMS bio-imaging. Biointerphases 2015, 10, 018902. [Google Scholar] [CrossRef]
- Kinoshita, M.; Suzuki, K.G.N.; Matsumori, N.; Takada, M.; Ano, H.; Morigaki, K.; Abe, M.; Makino, A.; Kobayashi, T.; Hirosawa, K.M.; et al. Raft-based sphingomyelin interactions revealed by new fluorescent sphingomyelin analogs. J. Cell Biol. 2017, 216, 1183–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megha; London, E. Ceramide selectively displaces cholesterol from ordered lipid domains (rafts): Implications for lipid raft structure and function. J. Biol. Chem. 2004, 279, 9997–10004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megha; Sawatzki, P.; Kolter, T.; Bittman, R.; London, E. Effect of Ceramide N-acyl chain and polar headgroup structure on the properties of ordered lipid domains (lipid rafts). Biochim. Biophys. Acta-Biomembr. 2007, 1768, 2205–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maula, T.; Urzelai, B.; Peter Slotte, J. The effects of N-acyl chain methylations on ceramide molecular properties in bilayer membranes. Eur. Biophys. J. 2011, 40, 857–863. [Google Scholar] [CrossRef]
- Alanko, S.M.K.; Halling, K.K.; Maunula, S.; Slotte, J.P.; Ramstedt, B. Displacement of sterols from sterol/sphingomyelin domains in fluid bilayer membranes by competing molecules. Biochim. Biophys. Acta-Biomembr. 2005, 1715, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, Y.; Ohba, T.; Miyata, H.; Ohki, K. Rapid phase change of lipid microdomains in giant vesicles induced by conversion of sphingomyelin to ceramide. Biochim. Biophys. Acta-Biomembr. 2006, 1758, 145–153. [Google Scholar] [CrossRef] [Green Version]
- González-Ramírez, E.J.; Artetxe, I.; García-Arribas, A.B.; Goni, F.M.; Alonso, A. Homogeneous and heterogeneous bilayers of ternary lipid compositions containing equimolar ceramide and cholesterol. Langmuir 2019, 35, 5305–5315. [Google Scholar] [CrossRef]
- Busto, J.V.; Sot, J.; Requejo-Isidro, J.; Goni, F.M.; Alonso, A. Cholesterol displaces palmitoylceramide from its tight packing with palmitoylsphingomyelin in the absence of a liquid-disordered phase. Biophys. J. 2010, 99, 1119–1128. [Google Scholar] [CrossRef] [Green Version]
- García-Arribas, A.B.; Axpe, E.; Mujika, J.I.; Mérida, D.; Busto, J.V.; Sot, J.; Alonso, A.; Lopez, X.; García, J.Á.; Ugalde, J.M.; et al. Cholesterol-ceramide interactions in phospholipid and sphingolipid bilayers as observed by positron annihilation lifetime spectroscopy and molecular dynamics simulations. Langmuir 2016, 32, 5434–5444. [Google Scholar] [CrossRef]
- Grassmé, H.; Jendrossek, V.; Riehle, A.; Von Kürthy, G.; Berger, J.; Schwarz, H.; Weller, M.; Kolesnick, R.; Gulbins, E. Host defense against pseudomonas aeruginosa requires ceramide-rich membrane rafts. Nat. Med. 2003, 9, 322–330. [Google Scholar] [CrossRef]
- Cremesti, A.E.; Goni, F.M.; Kolesnick, R. Role of sphingomyelinase and ceramide in modulating rafts: Do biophysical properties determine biologic outcome? FEBS Lett. 2002, 531, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Bollinger, C.R.; Teichgräber, V.; Gulbins, E. Ceramide-enriched membrane domains. Biochim. Biophys. Acta-Mol. Cell Res. 2005, 1746, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusumi, A.; Fujiwara, T.K.; Tsunoyama, T.A.; Kasai, R.S.; Liu, A.A.; Hirosawa, K.M.; Kinoshita, M.; Matsumori, N.; Komura, N.; Ando, H.; et al. Defining raft domains in the plasma membrane. Traffic 2020, 21, 106–137. [Google Scholar] [CrossRef]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid rafts: Controversies resolved, mysteries remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.F. Lipid rafts: Contentious only from simplistic standpoints. Nat. Rev. Mol. Cell Biol. 2006, 7, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.S. Lipid rafts: Now you see them, Now you don’t. Nat. Immunol. 2006, 7, 1139–1142. [Google Scholar] [CrossRef]
- Grassmé, H.; Jekle, A.; Riehle, A.; Schwarz, H.; Berger, J.; Sandhoff, K.; Kolesnick, R.; Gulbins, E. CD95 signaling via ceramide-rich membrane rafts. J. Biol. Chem. 2001, 276, 20589–20596. [Google Scholar] [CrossRef] [Green Version]
- Grassmé, H.; Schwarz, H.; Gulbins, E. Molecular mechanisms of ceramide-mediated CD95 clustering. Biochem. Biophys. Res. Commun. 2001, 284, 1016–1030. [Google Scholar] [CrossRef]
- Stancevic, B.; Kolesnick, R. Ceramide-rich platforms in transmembrane Signaling. FEBS Lett. 2010, 584, 1728–1740. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.G.N.; Kasai, R.S.; Hirosawa, K.M.; Nemoto, Y.L.; Ishibashi, M.; Miwa, Y.; Fujiwara, T.K.; Kusumi, A. Transient GPI-anchored protein homodimers are units for raft organization and function. Nat. Chem. Biol. 2012, 8, 774–783. [Google Scholar] [CrossRef]
- Adam, D.; Heinrich, M.; Kabelitz, D.; Schütze, S. Ceramide: Does it matter for T cells? Trends Immunol. 2002, 23, 1–4. [Google Scholar] [CrossRef]
- Grassmé, H.; Jendrossek, V.; Bock, J.; Riehle, A.; Gulbins, E. Ceramide-rich membrane rafts mediate CD40 clustering. J. Immunol. 2002, 168, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Janes, P.W.; Ley, S.C.; Magee, A.I.; Kabouridis, P.S. The role of lipid rafts in T cell antigen receptor (TCR) signalling. Semin. Immunol. 2000, 12, 23–34. [Google Scholar] [CrossRef]
- Gupta, N.; DeFranco, A.L. Lipid rafts and B cell signaling. Semin. Cell Dev. Biol. 2007, 18, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Bionda, C.; Hadchity, E.; Alphonse, G.; Chapet, O.; Rousson, R.; Rodriguez-Lafrasse, C.; Ardail, D. Radioresistance of human carcinoma cells is correlated to a defect in raft membrane clustering. Free Radic. Biol. Med. 2007, 43, 681–694. [Google Scholar] [CrossRef]
- Yamaji, A.; Sekizawa, Y.; Emoto, K.; Sakuraba, H.; Inoue, K.; Kobayashi, H.; Umeda, M. Lysenin, a novel sphingomyelin-specific binding protein. J. Biol. Chem. 1998, 273, 5300–5306. [Google Scholar] [CrossRef] [Green Version]
- Kiyokawa, E.; Makino, A.; Ishii, K.; Otsuka, N.; Yamaji-Hasegawa, A.; Kobayashi, T. Recognition of sphingomyelin by lysenin and lysenin-related proteins. Biochemistry 2004, 43, 9766–9773. [Google Scholar] [CrossRef]
- Dumitru, C.A.; Carpinteiro, A.; Trarbach, T.; Hengge, U.R.; Gulbins, E. Doxorubicin enhances TRAIL-induced cell death via ceramide-enriched membrane platforms. Apoptosis 2007, 12, 1533–1541. [Google Scholar] [CrossRef]
- Bock, J.; Szabó, I.; Gamper, N.; Adams, C.; Gulbins, E. Ceramide inhibits the potassium channel Kv1.3 by the formation of membrane platforms. Biochem. Biophys. Res. Commun. 2003, 305, 890–897. [Google Scholar] [CrossRef]
- Holmes, T.C.; Fadool, D.A.; Levitan, I.B. Tyrosine phosphorylation of the Kv1.3 potassium channel. J. Neurosci. 1996, 16, 1581–1590. [Google Scholar] [CrossRef]
- Filipp, D.; Zhang, J.; Leung, B.L.; Shaw, A.; Levin, S.D.; Veillette, A.; Julius, M. Regulation of Fyn through translocation of activated Lck into lipid rafts. J. Exp. Med. 2003, 197, 1221–1227. [Google Scholar] [CrossRef]
- Yasuda, H.; Torikai, K.; Kinoshita, M.; Sazzad, M.A.A.; Tsujimura, K.; Slotte, J.P.; Matsumori, N. Preparation of nitrogen analogues of ceramide and studies of their aggregation in sphingomyelin bilayers. Langmuir 2021, 37, 12438–12446. [Google Scholar] [CrossRef]
- Matsufuji, T.; Kinoshita, M.; Möuts, A.; Slotte, J.P.; Matsumori, N. Preparation and membrane properties of oxidized ceramide derivatives. Langmuir 2018, 34, 465–471. [Google Scholar] [CrossRef]
- Moro, K.; Nagahashi, M.; Gabriel, E.; Takabe, K.; Wakai, T. Clinical application of ceramide in cancer treatment. Breast Cancer 2018, 72, 2964–2979. [Google Scholar] [CrossRef]
- Boojar, M.M.A.; Boojar, M.M.A.; Golmohammad, S. Ceramide pathway: A novel approach to cancer chemotherapy. Egypt. J. Basic Appl. Sci. 2018, 5, 237–244. [Google Scholar]
- Brachtendorf, S.; El-Hindi, K.; Grösch, S. Ceramide synthases in cancer therapy and chemoresistance. Prog. Lipid Res. 2019, 74, 160–185. [Google Scholar] [CrossRef]
- Nagahashi, M.; Ramachandran, S.; Kim, E.Y.; Allegood, J.C.; Rashid, O.M.; Yamada, A.; Zhao, R.; Milstien, S.; Zhou, H.; Spiegel, S.; et al. Sphingosine-1-phosphate produced by sphingosine kinase 1 promotes breast cancer progression by stimulating angiogenesis and lymphangiogenesis. Cancer Res. 2012, 72, 726–735. [Google Scholar] [CrossRef] [Green Version]
- Salas, A.; Ponnusamy, S.; Senkal, C.E.; Meyers-Needham, M.; Selvam, S.P.; Saddoughi, S.A.; Apohan, E.R.; Sentelle, D.; Smith, C.; Gault, C.R.; et al. Sphingosine kinase-1 and sphingosine 1-phosphate receptor 2 mediate Bcr-Abl1 stability and drug resistance by modulation of protein phosphatase 2A. Blood 2011, 117, 5941–5952. [Google Scholar] [CrossRef] [Green Version]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid Res. 2013, 54, 5–19. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinoshita, M.; Matsumori, N. Inimitable Impacts of Ceramides on Lipid Rafts Formed in Artificial and Natural Cell Membranes. Membranes 2022, 12, 727. https://doi.org/10.3390/membranes12080727
Kinoshita M, Matsumori N. Inimitable Impacts of Ceramides on Lipid Rafts Formed in Artificial and Natural Cell Membranes. Membranes. 2022; 12(8):727. https://doi.org/10.3390/membranes12080727
Chicago/Turabian StyleKinoshita, Masanao, and Nobuaki Matsumori. 2022. "Inimitable Impacts of Ceramides on Lipid Rafts Formed in Artificial and Natural Cell Membranes" Membranes 12, no. 8: 727. https://doi.org/10.3390/membranes12080727