

The PH Domain and C-Terminal polyD Motif of Phafin2 Exhibit a Unique Concurrence in Animals

Abstract
:1. Introduction
2. Material and Methods
2.1. Prediction of the Three-Dimensional Structure of Phafin2
2.2. Sequence Retrieval and Acquisition
2.3. Functional Protein Domain and Motif Search
2.4. Multiple Sequence Alignment and Phylogenetic Analysis
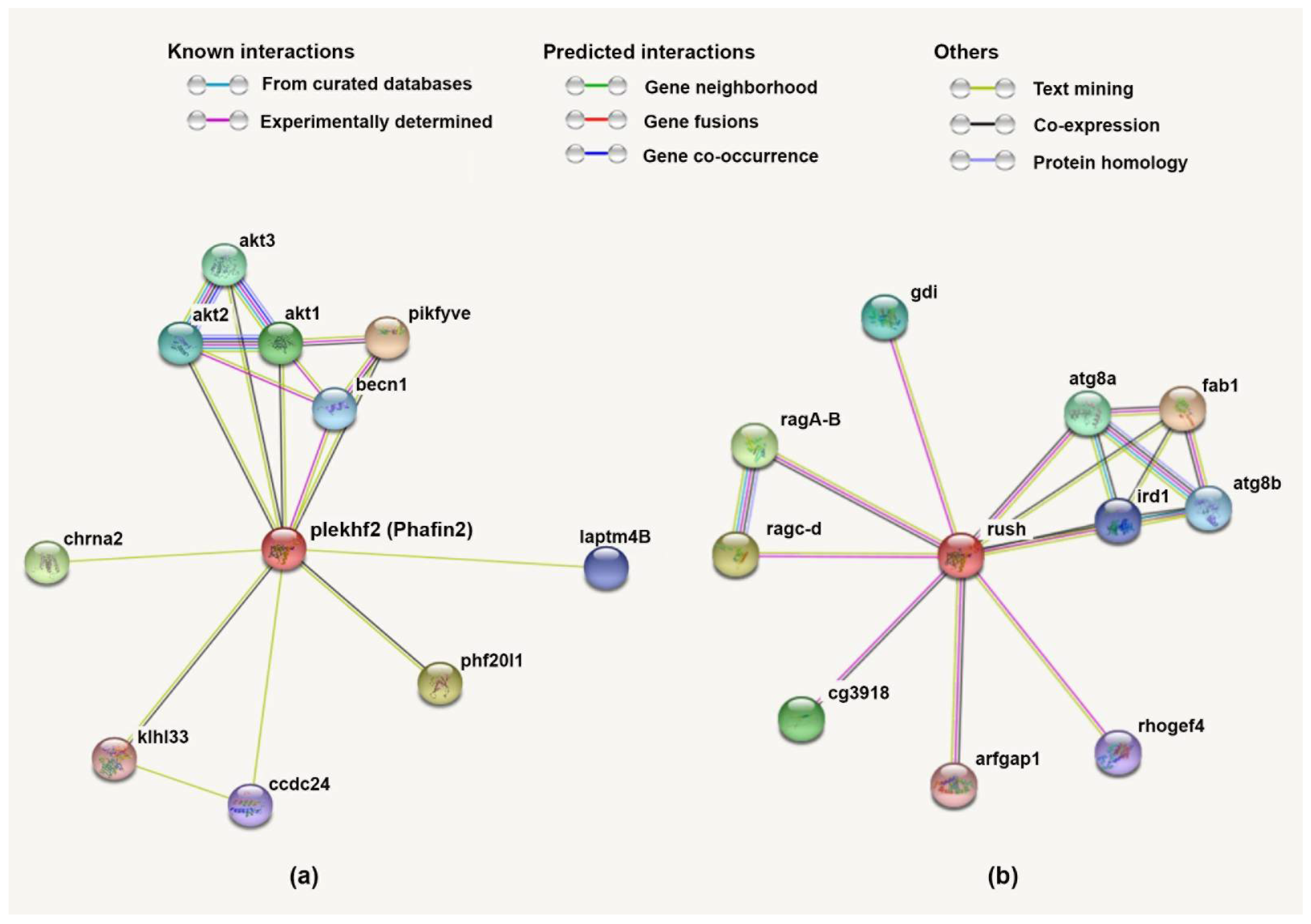
2.5. Protein–Protein Interactions Analyses
3. Results and Discussion
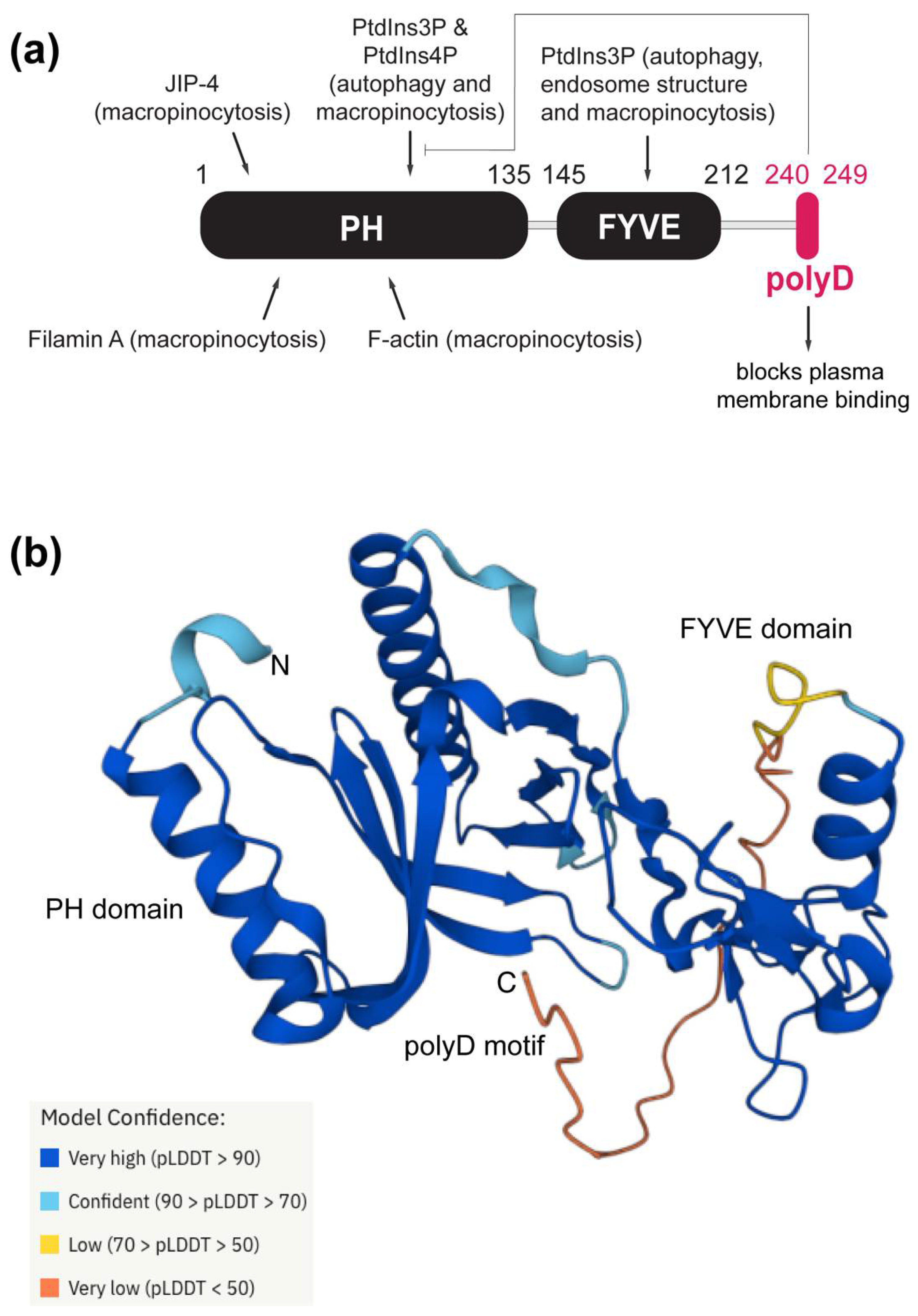
3.1. Genomic Features of the polyD Motif in Phafin2′s Human Homologs
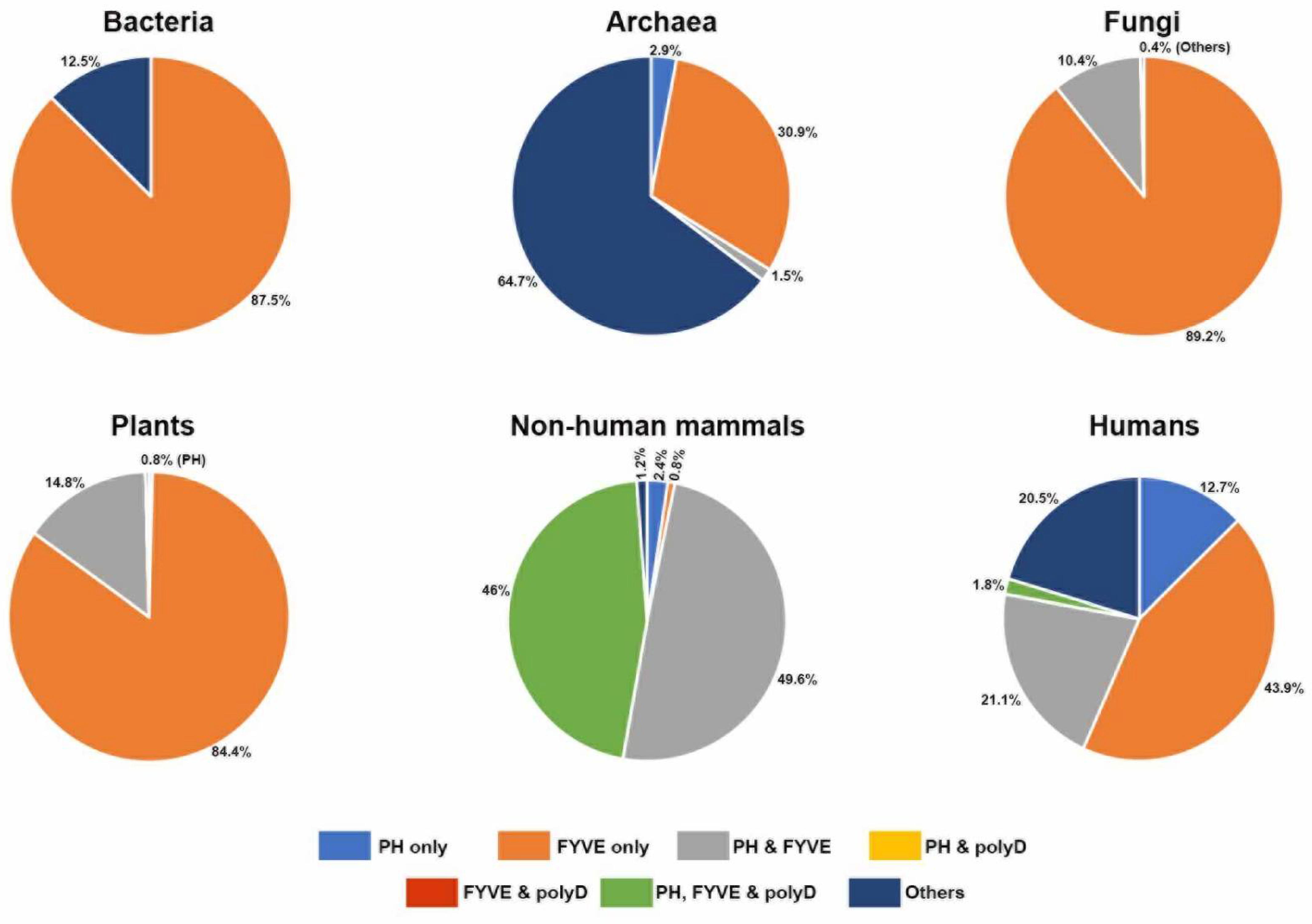
3.2. Concurrence of the polyD Motif with the PH and FYVE Domains in Phafin2 and Other Related PH-, FYVE-, or polyD-Containing Proteins
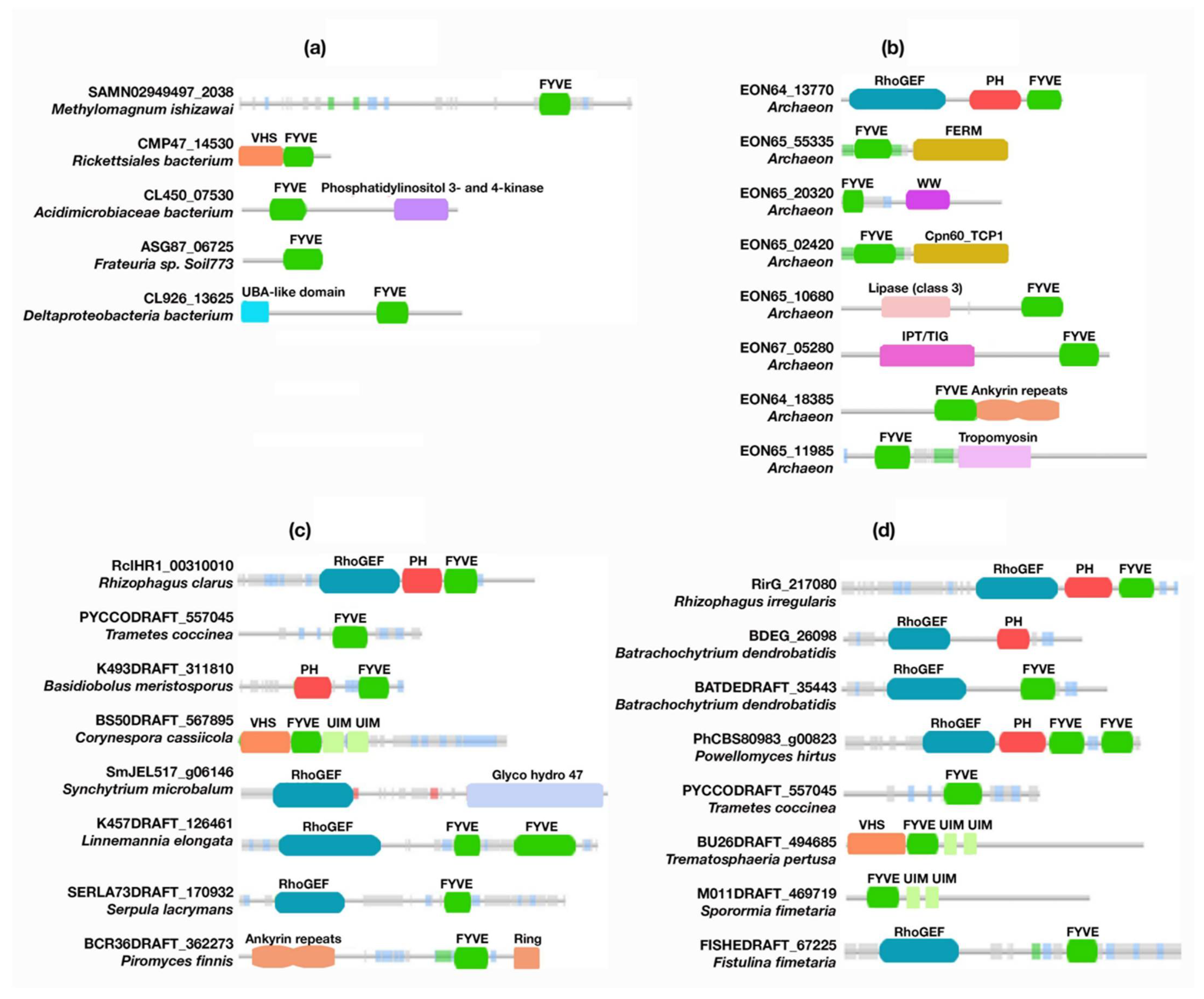
3.2.1. Bacteria
3.2.2. Archaea
3.2.3. Protozoans
3.2.4. Fungi
3.2.5. Plants
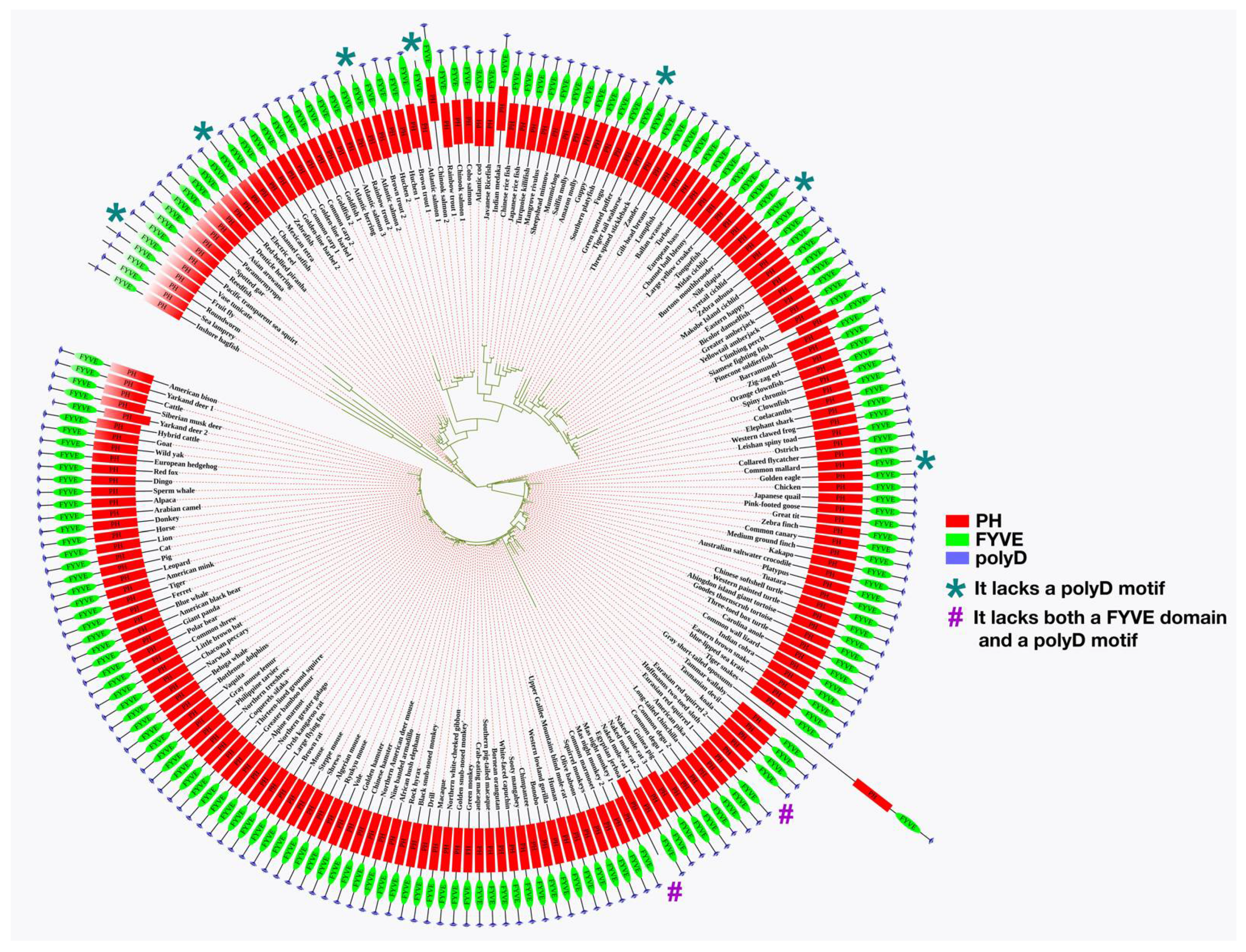
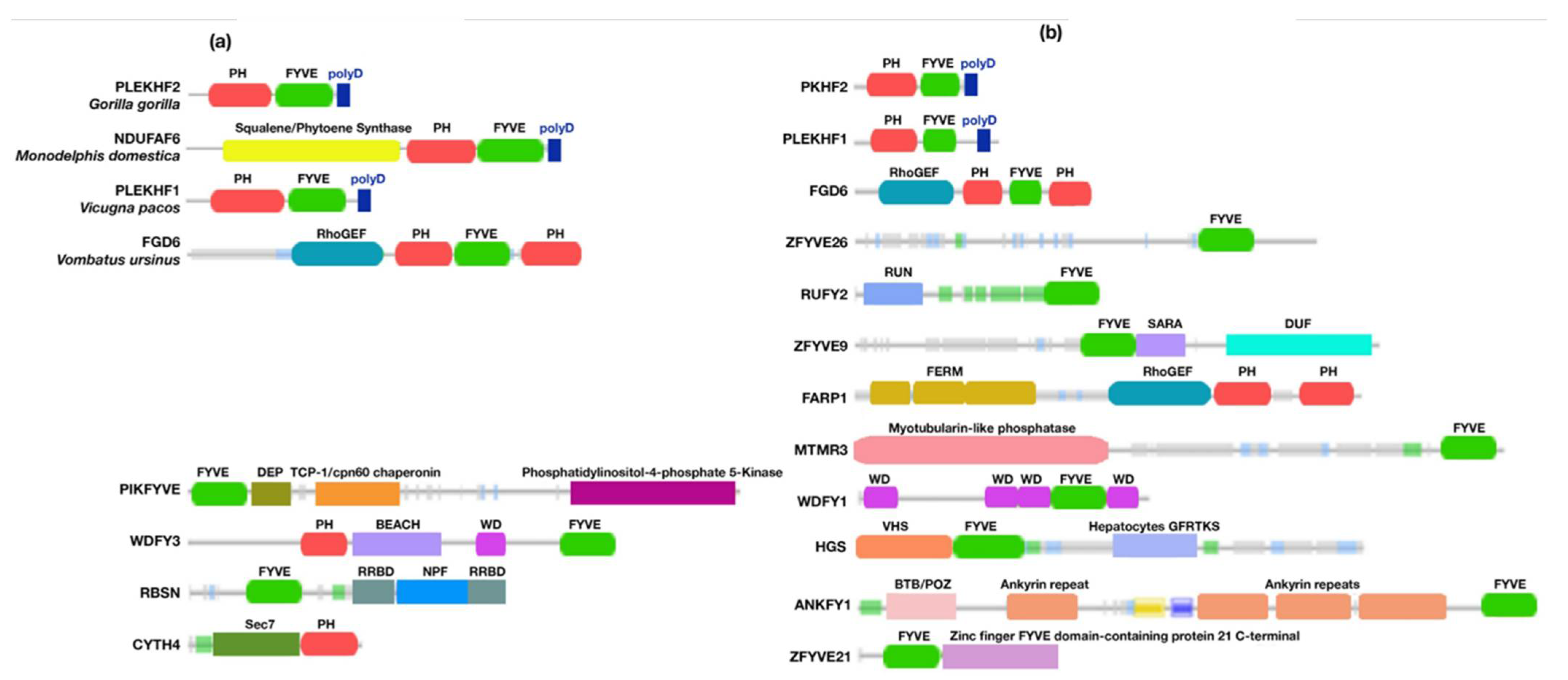
3.2.6. Animals
3.3. The Presence of Phafin2 or Related Proteins in Model Organisms and Their Structural Features
3.4. Protein–Protein Network and Functional Lineage of Human Phafin2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, T.-X.; Finkielstein, C.V.; Capelluto, D.G. The C-terminal acidic motif of Phafin2 inhibits PH domain binding to phosphatidylinositol 3-phosphate. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183230. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.-X.; Jo, A.; Deng, J.; Ellena, J.F.; Lazar, I.M.; Davis, R.; Capelluto, D.G.S. Structural, thermodynamic, and phosphatidylinositol 3-phosphate binding properties of Phafin2. Protein Sci. 2017, 26, 814–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsara, E.; Shamsara, J. Bioinformatics analysis of the genes involved in the extension of prostate cancer to adjacent lymph nodes by supervised and unsupervised machine learning methods: The role of SPAG1 and PLEKHF2. Genomics 2020, 112, 3871–3882. [Google Scholar] [CrossRef]
- Lin, W.-J.; Yang, C.-Y.; Lin, Y.-C.; Tsai, M.-C.; Yang, C.-W.; Tung, C.-Y.; Ho, P.-Y.; Kao, F.-J.; Lin, C.-H. Phafin2 modulates the structure and function of endosomes by a Rab5-dependent mechanism. Biochem. Biophys. Res. Commun. 2010, 391, 1043–1048. [Google Scholar] [CrossRef]
- Li, C.; Liu, Q.; Li, N.; Chen, W.; Wang, L.; Wang, Y.; Yu, Y.; Cao, X. EAPF/Phafin-2, a novel endoplasmic reticulum-associated protein, facilitates TNF-alpha-triggered cellular apoptosis through endoplasmic reticulum-mitochondrial apoptotic pathway. J. Mol. Med. 2008, 86, 471–484. [Google Scholar] [CrossRef]
- Schink, K.; Tan, K.; Spangenberg, H.; Martorana, D.; Sneeggen, M.; Campsteijn, S.; Raiborg, C.; Stenmark, H. The PtdIns3P-binding protein Phafin2 escorts macropinosomes through the cortical actin cytoskeleton. bioxRxiv, 2017; unpublished results. [Google Scholar] [CrossRef] [Green Version]
- Matsuda-Lennikov, M.; Suizu, F.; Hirata, N.; Hashimoto, M.; Kimura, K.; Nagamine, T.; Fujioka, Y.; Ohba, Y.; Iwanaga, T.; Noguchi, M. Lysosomal interaction of Akt with Phafin2: A critical step in the induction of autophagy. PLoS ONE 2014, 9, e79795. [Google Scholar] [CrossRef]
- Schink, K.O.; Tan, K.W.; Spangenberg, H.; Martorana, D.; Sneeggen, M.; Stévenin, V.; Enninga, J.; Campsteijn, C.; Raiborg, C.; Stenmark, H. The phosphoinositide coincidence detector Phafin2 promotes macropinocytosis by coordinating actin organisation at forming macropinosomes. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Lin, W.-J.; Yang, C.-Y.; Li, L.-L.; Yi, Y.-H.; Chen, K.-W.; Lin, Y.-C.; Liu, C.-C.; Lin, C.-H. Lysosomal targeting of phafin1 mediated by Rab7 induces autophagosome formation. Biochem. Biophys. Res. Commun. 2012, 417, 35–42. [Google Scholar] [CrossRef]
- Pedersen, N.M.; Raiborg, C.; Brech, A.; Skarpen, E.; Roxrud, I.; Platta, H.W.; Liestøl, K.; Stenmark, H. The PtdIns3P-binding protein phafin 2 mediates epidermal growth factor receptor degradation by promoting endosome fusion. Traffic 2012, 13, 1547–1563. [Google Scholar] [CrossRef]
- Gailite, I.; Egger-Adam, D.; Wodarz, A. The phosphoinositide-associated protein Rush hour regulates endosomal trafficking in Drosophila. Mol. Biol. Cell 2012, 23, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Saei, A.A.; Beusch, C.M.; Sabatier, P.; Wells, J.A.; Gharibi, H.; Meng, Z.; Chernobrovkin, A.; Rodin, S.; Näreoja, K.; Thorsell, A.-G.; et al. System-wide identification and prioritization of enzyme substrates by thermal analysis. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Hirata, N.; Suizu, F.; Matsuda-Lennikov, M.; Tanaka, T.; Edamura, T.; Ishigaki, S.; Donia, T.; Lithanatudom, P.; Obuse, C.; Iwanaga, T.; et al. Functional characterization of lysosomal interaction of Akt with VRK2. Oncogene 2018, 37, 5367–5386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.W.; Nähse, V.; Campsteijn, C.; Brech, A.; Schink, K.O.; Stenmark, H. JIP4 is recruited by the phosphoinositide-binding protein Phafin2 to promote recycling tubules on macropinosomes. J. Cell Sci. 2021, 134, jcs258495. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Sonnhammer, E.L.; Eddy, S.R.; Durbin, R. Pfam: A comprehensive database of protein domain families based on seed alignments. Proteins 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L.; et al. InterPro: The integrative protein signature database. Nucleic Acids Res. 2008, 37, D211–D215. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Prot. Bioinform. 2014, 48, 3.13.11–13.13.16. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mier, P.; Andrade-Navarro, M.A.; Perez-Pulido, A.J. Orthofind facilitates the discovery of homologous and orthologous proteins. PLoS ONE 2015, 10, e0143906. [Google Scholar] [CrossRef]
- Olsen, J.V.; Vermeulen, M.; Santamaria, A.; Kumar, C.; Miller, M.L.; Jensen, L.J.; Gnad, F.; Cox, J.; Jensen, T.S.; Nigg, E.A.; et al. Quantitative phosphoproteomics reveals widespread full phosphorylation site occupancy during mitosis. Sci. Signal. 2010, 3, ra3. [Google Scholar] [CrossRef]
- Xu, Q.; Bateman, A.; Finn, R.D.; Abdubek, P.; Astakhova, T.; Axelrod, H.L.; Bakolitsa, C.; Carlton, D.; Chen, C.; Chiu, H.-J.; et al. Bacterial pleckstrin homology domains: A prokaryotic origin for the PH domain. J. Mol. Biol. 2010, 396, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffzek, K.; Welti, S. Pleckstrin homology (PH) like domains—Versatile modules in protein-protein interaction platforms. FEBS Lett. 2012, 586, 2662–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.; Mandal, S.; Banerjee, S.; Ghosh, A.; Ganguly, S.; Sil, A.K.; Sarkar, S. Identification and Characterization of a FYVE domain from the early diverging eukaryote giardia lamblia. Curr. Microbiol. 2010, 62, 1179–1184. [Google Scholar] [CrossRef]
- Roach, T.G.; Lång, H.K.M.; Xiong, W.; Ryhänen, S.J.; Capelluto, D.G.S. Protein trafficking or cell signaling: A dilemma for the adaptor protein TOM1. Front. Cell Dev. Biol. 2021, 9, 643769. [Google Scholar] [CrossRef]
- Pruneda, J.N.; Durkin, C.H.; Geurink, P.P.; Ovaa, H.; Santhanam, B.; Holden, D.W.; Komander, D. The molecular basis for ubiquitin and ubiquitin-like specificities in bacterial effector proteases. Mol. Cell 2016, 63, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Su, V.; Lau, A.F. Ubiquitin-like and ubiquitin-associated domain proteins: Significance in proteasomal degradation. Cell. Mol. Life Sci. 2009, 66, 2819–2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zientara-Rytter, K.; Subramani, S. The roles of ubiquitin-binding protein shuttles in the degradative fate of ubiquitinated proteins in the ubiquitin-proteasome system and autophagy. Cells 2019, 8, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boura, E.; Nencka, R. Phosphatidylinositol 4-kinases: Function, structure, and inhibition. Exp. Cell Res. 2015, 337, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Roymans, D.; Slegers, H. Phosphatidylinositol 3-kinases in tumor progression. JBIC J. Biol. Inorg. Chem. 2001, 268, 487–498. [Google Scholar] [CrossRef]
- Tang, M.R.; Sternberg, D.; Behr, R.K.; Sloma, A.; Berka, R.M. Use of transcriptional profiling & bioinformatics to solve production problems: Eliminating red pigment production in a Bacillus subtilis strain producing hyaluronic acid. Ind. Biotech. 2006, 2, 66–74. [Google Scholar]
- Bunai, K.; Nozaki, M.; Hamano, M.; Ogane, S.; Inoue, T.; Nemoto, T.; Nakanishi, H.; Yamane, K. Proteomic analysis of acrylamide gel separated proteins immobilized on polyvinylidene difluoride membranes following proteolytic digestion in the presence of 80% acetonitrile. Proteomics 2003, 3, 1738–1749. [Google Scholar] [CrossRef]
- Cao, M.; Kobel, P.A.; Morshedi, M.M.; Wu, M.F.W.; Paddon, C.; Helmann, J.D. Defining the Bacillus subtilis σW regulon: A comparative analysis of promoter consensus search, run-off transcription/macroarray analysis (ROMA), and transcriptional profiling approaches. J. Mol. Biol. 2002, 316, 443–457. [Google Scholar] [CrossRef]
- Cao, M.; Wang, T.; Ye, R.; Helmann, J.D. Antibiotics that inhibit cell wall biosynthesis induce expression of the Bacillus subtilisσW and σM regulons. Mol. Microbiol. 2002, 45, 1267–1276. [Google Scholar] [CrossRef]
- Akimkina, T.; Vénien-Bryan, C.; Hodgkin, J. Isolation, characterization and complete nucleotide sequence of a novel temperate bacteriophage Min1, isolated from the nematode pathogen Microbacterium nematophilum. Res. Microbiol. 2007, 158, 582–590. [Google Scholar] [CrossRef]
- Labrie, S.; Moineau, S. Complete Genomic Sequence of Bacteriophage ul36: Demonstration of phage heterogeneity within the P335 quasi-species of lactococcal phages. Virology 2002, 296, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegert, T.; Homuth, G.; Versteeg, S.; Schumann, W. Alkaline shock induces the Bacillus subtilis sigma(W) regulon. Mol. Microbiol. 2001, 41, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Wecke, T.; Veith, B.; Ehrenreich, A.; Mascher, T. Cell envelope stress response in Bacillus licheniformis: Integrating comparative genomics, transcriptional profiling, and regulon mining to decipher a complex regulatory network. J. Bacteriol. 2006, 188, 7500–7511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botero, S.; Chiaroni-Clarke, R.; Simon, S.M. Escherichia coli as a platform for the study of phosphoinositide biology. Sci. Adv. 2019, 5, eaat4872. [Google Scholar] [CrossRef] [Green Version]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [Green Version]
- Pace, N.R. Mapping the tree of life: Progress and prospects. Microbiol. Mol. Biol. Rev. 2009, 73, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Guan, X.; Dong, C.; Jiao, Z. Rho GTPases and related signaling complexes in cell migration and invasion. Exp. Cell Res. 2020, 388, 111824. [Google Scholar] [CrossRef]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Jung, O.; Choi, S.; Jang, S.-B.; Lee, S.-A.; Lim, S.-T.; Choi, Y.-J.; Kim, H.-J.; Kim, D.-H.; Kwak, T.K.; Kim, H.; et al. Tetraspan TM4SF5-dependent direct activation of FAK and metastatic potential of hepatocarcinoma cells. J. Cell Sci. 2012, 125, 5960–5973. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhu, Y.; Ye, S.; Zhang, R. Crystal structure of kindlin-2 PH domain reveals a conformational transition for its membrane anchoring and regulation of integrin activation. Protein Cell 2012, 3, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Nurniwalis, A.W.; Zubaidah, R.; Akmar, A.S.N.; Zulkifli, H.; Arif, M.A.M.; Massawe, F.; Chan, K.L.; Parveez, G.K.A. Genomic structure and characterization of a lipase class 3 gene and promoter from oil palm. Biol. Plant. 2015, 59, 227–236. [Google Scholar] [CrossRef]
- Oguchi, Y.; Ishizuka, J.; Hitchcock-DeGregori, S.E.; Ishiwata, S.; Kawai, M. The role of tropomyosin domains in cooperative activation of the actin–myosin interaction. J. Mol. Biol. 2011, 414, 667–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Khodor, S.; Price, C.T.; Kalia, A.; Abu Kwaik, Y. Functional diversity of ankyrin repeats in microbial proteins. Trends Microbiol. 2010, 18, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Nakada-Tsukui, K.; Nozaki, T. Diversity of phosphoinositide binding proteins in Entamoeba histolytica. Parasitol. Int. 2021, 83, 102367. [Google Scholar] [CrossRef] [PubMed]
- Mertens, H.D.; Callaghan, J.M.; Swarbrick, J.D.; McConville, M.J.; Gooley, P.R. A high-resolution solution structure of a trypanosomatid FYVE domain. Protein Sci. 2007, 16, 2552–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattioni, A.; Boldt, K.; Auciello, G.; Komada, M.; Rappoport, J.Z.; Ueffing, M.; Castagnoli, L.; Cesareni, G.; Santonico, E. Ring finger protein 11 acts on ligand-activated EGFR via the direct interaction with the UIM region of ANKRD13 protein family. FEBS J. 2020, 287, 3526–3550. [Google Scholar] [CrossRef] [PubMed]
- Shertz, C.A.; Cardenas, M.E. Exploiting and subverting Tor signaling in the pathogenesis of fungi, parasites, and viruses. PLoS Pathog. 2011, 7, e1002269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef]
- Eltschinger, S.; Loewith, R. TOR complexes and the maintenance of cellular homeostasis. Trends Cell Biol. 2015, 26, 148–159. [Google Scholar] [CrossRef]
- Berchtold, D.; Walther, T.C. TORC2 plasma membrane localization is essential for cell viability and restricted to a distinct domain. Mol. Biol. Cell 2009, 20, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Mulet, J.M.; Martin, D.E.; Loewith, R.; Hall, M.N. Mutual antagonism of target of rapamycin and calcineurin signaling. J. Biol. Chem. 2006, 281, 33000–33007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelants, F.M.; Leskoske, K.L.; Pedersen, R.T.; Muir, A.; Liu, J.M.-H.; Finnigan, G.C.; Thorner, J. TOR complex 2-regulated protein kinase Fpk1 stimulates endocytosis via inhibition of Ark1/Prk1-related protein kinase Akl1 in Saccharomyces cerevisiae. Mol. Cell Biol. 2017, 37, e00627-00616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- deHart, A.K.; Schnell, J.D.; Allen, D.A.; Hicke, L. The conserved Pkh–Ypk kinase cascade is required for endocytosis in yeast. J. Cell Biol. 2002, 156, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Barman, A.; Gohain, D.; Bora, U.; Tamuli, R. Phospholipases play multiple cellular roles including growth, stress tolerance, sexual development, and virulence in fungi. Microbiol. Res. 2018, 209, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Irvine, R.F. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature 1984, 312, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.-A.; Takeuchi, H.; Kanematsu, T.; Allen, V.; Yagisawa, H.; Kikkawa, U.; Watanabe, Y.; Nakasima, A.; Katan, M.; Hirata, M. Involvement of EF hand motifs in the Ca(2+)-dependent binding of the pleckstrin homology domain to phosphoinositides. Eur. J. Biol. Inorg. Chem. 1999, 265, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Li, Y.; Li, D.; Bi, Y.; Prusky, D.B.; Dong, Y.; Wang, T.; Zhang, M.; Zhang, X.; Liu, Y. Phospholipase C from alternaria alternata is induced by physiochemical cues on the pear fruit surface that dictate infection structure differentiation and pathogenicity. Front. Microbiol. 2020, 11, 1279. [Google Scholar] [CrossRef]
- Rho, H.-S.; Jeon, J.; Lee, Y.-H. Phospholipase C-mediated calcium signalling is required for fungal development and pathogenicity in Magnaporthe oryzae. Mol. Plant Pathol. 2009, 10, 337–346. [Google Scholar] [CrossRef]
- Gresset, A.; Sondek, J.; Harden, T.K. The phospholipase C isozymes and their regulation. Subcell. Biochem. 2012, 58, 61–94. [Google Scholar]
- Bilkova, E.; Pleskot, R.; Rissanen, S.; Sun, S.; Czogalla, A.; Cwiklik, L.; Róg, T.; Vattulainen, I.; Cremer, P.S.; Jungwirth, P. Calcium directly regulates phosphatidylinositol 4, 5-bisphosphate headgroup conformation and recognition. J. Am. Chem. Soc. 2017, 139, 4019–4024. [Google Scholar] [CrossRef]
- Schumacher, J.; Viaud, M.; Simon, A.; Tudzynski, B. The Gα subunit BCG1, the phospholipase C (BcPLC1) and the calcineurin phosphatase co-ordinately regulate gene expression in the grey mould fungus Botrytis cinerea. Mol. Microbiol. 2008, 67, 1027–1050. [Google Scholar] [CrossRef] [PubMed]
- Pukkila, P.J. Coprinopsis cinerea. Curr. Biol. 2011, 21, R616–R617. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.T.; Ahn, C.-S.; Lee, K.-J.; Kim, J.-G.; Ro, H.-S.; Kim, J.W.; Lee, C.-W. The activity of phosphoinositide-specific phospholipase C is required for vegetative growth and cell wall regeneration in Coprinopsis cinerea. J. Microbiol. 2012, 50, 689–692. [Google Scholar] [CrossRef]
- Essen, L.-O.; Perisic, O.; Cheung, R.; Katan, M.; Williams, R.L. Crystal structure of a mammalian phosphoinositide-specific phospholipase Cδ. Nature 1996, 380, 595–602. [Google Scholar] [CrossRef] [PubMed]
- de Jong, F.; Munnik, T. Attracted to Membranes: Lipid-Binding Domains in Plants; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- de Bruijn, F.J. The Model Legume Medicago truncatula, 2 Volume Set; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Agorio, A.; Giraudat, J.; Bianchi, M.W.; Marion, J.; Espagne, C.; Castaings, L.; Lelièvre, F.; Curie, C.; Thomine, S.; Merlot, S. Phosphatidylinositol 3-phosphate–binding protein AtPH1 controls the localization of the metal transporter NRAMP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E3354–E3363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, D.; Pleskot, R.; Pejchar, P.; Potocký, M.; Trpkošová, P.; Cwiklik, L.; Vukašinović, N.; Sternberg, H.; Yalovsky, S.; Žárský, V. Exocyst SEC3 and phosphoinositides define sites of exocytosis in pollen tube initiation and growth. Plant Physiol. 2016, 172, 980–1002. [Google Scholar] [CrossRef] [Green Version]
- Simon, M.L.A.; Platre, M.P.; Marquès-Bueno, M.M.; Armengot, L.; Stanislas, T.; Bayle, V.; Caillaud, M.-C.; Jaillais, Y. A PtdIns(4)P-driven electrostatic field controls cell membrane identity and signalling in plants. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Di Lello, P.; Nguyen, B.D.; Jones, T.N.; Potempa, K.; Kobor, M.S.; Legault, P.; Omichinski, J.G. NMR Structure of the Amino-terminal domain from the Tfb1 subunit of TFIIH and characterization of its phosphoinositide and VP16 binding sites. Biochemistry 2005, 44, 7678–7686. [Google Scholar] [CrossRef]
- Fort, P.; Blangy, A. The evolutionary landscape of Dbl-Like RhoGEF families: Adapting eukaryotic cells to environmental signals. Genome Biol. Evol. 2017, 9, 1471–1486. [Google Scholar] [CrossRef] [Green Version]
- Cerione, R.A.; Zheng, Y. The Dbl family of oncogenes. Curr. Opin. Cell Biol. 1996, 8, 216–222. [Google Scholar] [CrossRef]
- Li, N.; Zheng, Y.; Chen, W.; Wang, C.; Liu, X.; He, W.; Xu, H.; Cao, X. Adaptor protein LAPF recruits phosphorylated p53 to lysosomes and triggers lysosomal destabilization in apoptosis. Cancer Res. 2007, 67, 11176–11185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Qin, K.; Li, N.; Han, C.; Cao, X. An endosomal LAPF is required for macrophage endocytosis and elimination of bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 12958–12963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, W.; Schilcher, P.; Zeitlmann, L.; Kolanus, W. The PH Domain and the polybasic c domain of cytohesin-1 cooperate specifically in plasma membrane association and cellular function. Mol. Biol. Cell 1998, 9, 1981–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Gan, W.; Chin, Y.R.; Ogura, K.; Guo, J.; Zhang, J.; Wang, B.; Blenis, J.; Cantley, L.C.; Toker, A. PtdIns(3,4,5)P3-dependent activation of the mTORC2 kinase complex. Cancer Discov. 2015, 5, 1194–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkateswarlu, K. Interaction protein for cytohesin exchange factors 1 (IPCEF1) binds cytohesin 2 and modifies its activity. J. Biol. Chem. 2003, 278, 43460–43469. [Google Scholar] [CrossRef] [Green Version]
- Hiester, K.G.; Santy, L.C. The cytohesin coiled-coil domain interacts with threonine 276 to control membrane association. PLoS ONE 2013, 8, e82084. [Google Scholar] [CrossRef]
- Ngo, M.; Ridgway, N.D. Oxysterol binding protein–related protein 9 (ORP9) is a cholesterol transfer protein that regulates golgi structure and function. Mol. Biol. Cell 2009, 20, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, A.; Perry, R.J.; Storey, M.K.; Cook, H.W.; Byers, D.M.; Ridgway, N.D. Golgi localization and phosphorylation of oxysterol binding protein in Niemann-Pick C and U18666A-treated cells. J. Lipid Res. 2001, 42, 1062–1071. [Google Scholar] [CrossRef]
- Singh, N.; Reyes-Ordoñez, A.; Compagnone, M.A.; Moreno, J.F.; Leslie, B.J.; Ha, T.; Chen, J. Redefining the specificity of phosphoinositide-binding by human PH domain-containing proteins. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; He, X.; Coleman, A.; Chen, Y.-J.; Dasari, P.; Liou, J.; Biederer, T.; Zhang, X. Structural analyses of FERM domain-mediated membrane localization of FARP1. Sci. Rep. 2018, 8, 10477. [Google Scholar] [CrossRef]
- He, X.; Kuo, Y.-C.; Rosche, T.J.; Zhang, X. Structural basis for autoinhibition of the guanine nucleotide exchange factor FARP2. Structure 2013, 21, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landgraf, K.E.; Pilling, C.; Falke, J.J. Molecular mechanism of an oncogenic mutation that alters membrane targeting: Glu17Lys Modifies the PIP lipid specificity of the AKT1 PH domain. Biochemistry 2008, 47, 12260–12269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, N.; Salguero, A.L.; Liu, A.Z.; Chen, Z.; Dempsey, D.R.; Ficarro, S.B.; Alexander, W.M.; Marto, J.A.; Li, Y.; Amzel, L.M.; et al. Akt kinase activation mechanisms revealed using protein semisynthesis. Cell 2018, 174, 897–907.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, N.; Viennet, T.; Bae, H.; Salguero, A.; Boeszoermenyi, A.; Arthanari, H.; A Cole, P. The structural determinants of PH domain-mediated regulation of Akt revealed by segmental labeling. eLife 2020, 9, e59151. [Google Scholar] [CrossRef]
- Shidal, C.; Inaba, J.-I.; Yaddanapudi, K.; Davis, K.R. The soy-derived peptide Lunasin inhibits invasive potential of melanoma initiating cells. Oncotarget 2017, 8, 25525–25541. [Google Scholar] [CrossRef] [Green Version]
- Okuda, M.; Araki, K.; Ohtani, K.; Nishimura, Y. The interaction mode of the acidic region of the cell cycle transcription factor DP1 with TFIIH. J. Mol. Biol. 2016, 428, 4993–5006. [Google Scholar] [CrossRef]
- Filimonenko, M.; Isakson, P.; Finley, K.D.; Anderson, M.; Jeong, H.; Melia, T.J.; Bartlett, B.J.; Myers, K.M.; Birkeland, H.C.; Lamark, T.; et al. The selective macroautophagic degradation of aggregated proteins requires the PI3P-binding protein alfy. Mol. Cell 2010, 38, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Clausen, T.H.; Lamark, T.; Isakson, P.; Finley, K.D.; Larsen, K.B.; Brech, A.; Øvervatn, A.; Stenmark, H.; Bjørkøy, G.; Simonsen, A. p62/SQSTM1 and ALFY interact to facilitate the formation of p62 bodies/ALIS and their degradation by autophagy. Autophagy 2010, 6, 330–344. [Google Scholar] [CrossRef] [Green Version]
- Chung, T. How phosphoinositides shape autophagy in plant cells. Plant Sci. 2019, 281, 146–158. [Google Scholar] [CrossRef]
- Teh, O.-K.; Hatsugai, N.; Tamura, K.; Fuji, K.; Tabata, R.; Yamaguchi, K.; Shingenobu, S.; Yamada, M.; Hasebe, M.; Sawa, S.; et al. BEACH-domain proteins act together in a cascade to mediate vacuolar protein trafficking and disease resistance in arabidopsis. Mol. Plant 2015, 8, 389–398. [Google Scholar] [CrossRef] [Green Version]
- De Lozanne, A. The role of BEACH proteins in dictyostelium. Traffic 2003, 4, 6–12. [Google Scholar] [CrossRef]
- Saedler, R.; Jakoby, M.; Marin, B.; Galiana-Jaime, E.; Hülskamp, M. The cell morphogenesis gene SPIRRIG in Arabidopsis encodes a WD/BEACH domain protein. Plant J. 2009, 59, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Steffens, A.; Jakoby, M.; Hülskamp, M. Physical, functional and genetic interactions between the BEACH domain protein SPIRRIG and LIP5 and SKD1 and its role in endosomal trafficking to the vacuole in arabidopsis. Front. Plant Sci. 2017, 8, 1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wywial, E.; Singh, S.M. Identification and structural characterization of FYVE domain-containing proteins of Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Wei, J.; Gao, C. Functional analysis of plant FYVE domain proteins in endosomal trafficking. Methods Mol. Biol. 2020, 2177, 83–94. [Google Scholar]
- Whitley, P.; Hinz, S.; Doughty, J. Arabidopsis FAB1/PIKfyve proteins are essential for development of viable pollen. Plant Physiol. 2009, 151, 1812–1822. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Matsuzawa, T.; Takegawa, K.; Sato, M.H. Loss-of-function and gain-of-function mutations in FAB1A/B impair endomembrane homeostasis, conferring pleiotropic developmental abnormalities in Arabidopsis. Plant Physiol. 2011, 155, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Zhuang, X.; Cui, Y.; Fu, X.; He, Y.; Zhao, Q.; Zeng, Y.; Shen, J.; Luo, M.; Jiang, L. Dual roles of an Arabidopsis ESCRT component FREE1 in regulating vacuolar protein transport and autophagic degradation. Proc. Natl. Acad. Sci. USA 2015, 112, 1886–1891. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Luo, M.; Zhao, Q.; Yang, R.; Cui, Y.; Zeng, Y.; Xia, J.; Jiang, L. A unique plant ESCRT component, FREE1, regulates multivesicular body protein sorting and plant growth. Curr. Biol. 2014, 24, 2556–2563. [Google Scholar] [CrossRef] [Green Version]
- Kolb, C.; Nagel, M.-K.; Kalinowska, K.; Hagmann, J.; Ichikawa, M.; Anzenberger, F.; Alkofer, A.; Sato, M.H.; Falter-Braun, P.; Isono, E. FYVE1 is essential for vacuole biogenesis and intracellular trafficking in arabidopsis. Plant Physiol. 2015, 167, 1361–1373. [Google Scholar] [CrossRef] [Green Version]
- Sutipatanasomboon, A.; Herberth, S.; Alwood, E.G.; Häweker, H.; Müller, B.; Shahriari, M.; Zienert, A.Y.; Marin, B.; Robatzek, S.; Praefcke, G.J.K.; et al. Disruption of the plant-specific CFS1 gene impairs autophagosome turnover and triggers EDS1-dependent cell death. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagishi, Y.; Matsuda, S. RUFY, rab and rap family proteins involved in a regulation of cell polarity and membrane trafficking. Int. J. Mol. Sci. 2013, 14, 6487–6498. [Google Scholar] [CrossRef] [Green Version]
- van Nocker, S.; Ludwig, P. The WD-repeat protein superfamily in Arabidopsis: Conservation and divergence in structure and function. BMC Genom. 2003, 4, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamalesh, K.; Trivedi, D.; Toscano, S.; Sharma, S.; Kolay, S.; Raghu, P. Phosphatidylinositol 5-phosphate 4-kinase regulates early endosomal dynamics during clathrin-mediated endocytosis. J. Cell Sci. 2017, 130, 2119–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, H.A.; Dionne, H.; Rusten, T.E.; Brech, A.; Fisher, W.W.; Pfeiffer, B.D.; Celniker, S.E.; Stenmark, H.; Bilder, D. Regulation of early endosomal entry by the Drosophila tumor suppressors rabenosyn and Vps45. Mol. Biol. Cell 2008, 19, 4167–4176. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Nickitenko, A.; Duan, X.; Lloyd, T.E.; Wu, M.N.; Bellen, H.; Quiocho, F.A. Crystal structure of the VHS and FYVE tandem domains of Hrs, a protein involved in membrane trafficking and signal transduction. Cell 2000, 100, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Consortium, C.e.S. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 1998, 282, 2012–2018. [Google Scholar]
- Reinhart, E.F.; Litt, N.A.; Katzenell, S.; Pellegrini, M.; Yamamoto, A.; Ragusa, M.J. A highly conserved glutamic acid in ALFY inhibits membrane binding to aid in aggregate clearance. Traffic 2021, 22, 23–37. [Google Scholar] [CrossRef]
- Posiri, P.; Thongsuksangcharoen, S.; Chaysri, N.; Panyim, S.; Ongvarrasopone, C. PmEEA1, the early endosomal protein is employed by YHV for successful infection in Penaeus monodon. Fish Shellfish Immunol. 2019, 95, 449–455. [Google Scholar] [CrossRef]
- Hermle, T.; Schneider, R.; Schapiro, D.; Braun, D.A.; van der Ven, A.T.; Warejko, J.K.; Daga, A.; Widmeier, E.; Nakayama, M.; Jobst-Schwan, T.; et al. GAPVD1 and ANKFY1 mutations implicate RAB5 regulation in nephrotic syndrome. J. Am. Soc. Nephrol. 2018, 29, 2123–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maekawa, M.; Tanigawa, K.; Sakaue, T.; Hiyoshi, H.; Kubota, E.; Joh, T.; Watanabe, Y.; Taguchi, T.; Higashiyama, S. Cullin-3 and its adaptor protein ANKFY1 determine the surface level of integrin beta1 in endothelial cells. Biol. Open 2017, 6, 1707–1719. [Google Scholar] [PubMed] [Green Version]
- Liu, K.; Jian, Y.; Sun, X.; Yang, C.; Gao, Z.; Zhang, Z.; Liu, X.; Li, Y.; Xu, J.; Jing, Y. Negative regulation of phosphatidylinositol 3-phosphate levels in early-to-late endosome conversionRegulation of endosomal PtdIns3P. J. Cell Biol. 2016, 212, 181–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.; Guo, M.; Gan, M.; Yu, B.; Tian, X.; Wang, J.-B. TRIM59 regulates autophagy through modulating both the transcription and the ubiquitination of BECN1. Autophagy 2018, 14, 2035–2048. [Google Scholar] [CrossRef] [Green Version]
- Maria Fimia, G.; Stoykova, A.; Romagnoli, A.; Giunta, L.; Di Bartolomeo, S.; Nardacci, R.; Corazzari, M.; Fuoco, C.; Ucar, A.; Schwartz, P.; et al. Ambra1 regulates autophagy and development of the nervous system. Nature 2007, 447, 1121–1125. [Google Scholar] [CrossRef] [Green Version]
- Usman, R.M.; Razzaq, F.; Akbar, A.; Farooqui, A.A.; Iftikhar, A.; Latif, A.; Hassan, H.; Zhao, J.; Carew, J.S.; Nawrocki, S.T.; et al. Role and mechanism of autophagy-regulating factors in tumorigenesis and drug resistance. Asia-Pac. J. Clin. Oncol. 2020, 17, 193–208. [Google Scholar] [CrossRef]
- Vergarajauregui, S.; Martina, J.; Puertollano, R. LAPTMs regulate lysosomal function and interact with mucolipin 1: New clues for understanding mucolipidosis type IV. J. Cell Sci. 2011, 124, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Karabiyik, C.; Vicinanza, M.; Son, S.; Rubinsztein, D. Glucose starvation induces autophagy via ULK1-mediated activation of PIKfyve in an AMPK-dependent manner. Dev. Cell 2021, 56, 1961–1975.e1965. [Google Scholar] [CrossRef]
- Desale, S.E.; Chinnathambi, S. Phosphoinositides signaling modulates microglial actin remodeling and phagocytosis in Alzheimer’s disease. Cell Commun. Signal. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Carr, S.M.; Munro, S.; A Sagum, C.; Fedorov, O.; Bedford, M.T.; La Thangue, N.B. Tudor-domain protein PHF20L1 reads lysine methylated retinoblastoma tumour suppressor protein. Cell Death Differ. 2017, 24, 2139–2149. [Google Scholar] [CrossRef] [Green Version]
- Sharp, L.; Brannigan, G. Spontaneous lipid binding to the nicotinic acetylcholine receptor in a native membrane. J. Chem. Phys. 2021, 154, 185102. [Google Scholar] [CrossRef] [PubMed]
- Okada, M. Can rodent models elucidate the pathomechanisms of genetic epilepsy? Br. J. Pharm. 2022, 179, 1620–1639. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Libertini, S.J.; George, M.; Dandekar, S.; Tepper, C.G.; Al-Bataina, B.; Kung, H.-J.; Ghosh, P.M.; Mudryj, M. Genome-wide analysis of androgen receptor binding and gene regulation in two CWR22-derived prostate cancer cell lines. Endocr. Relat. Cancer 2010, 17, 857–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Wang, S.; Zhang, L.; Liu, L.; Wen, Y.; Ma, M.; Cheng, S.; Li, P.; Cheng, B.; Du, Y.; et al. An integrative analysis of transcriptome-wide association study and mRNA expression profile identified candidate genes for attention-deficit/hyperactivity disorder. Psychiatry Res. 2019, 282, 112639. [Google Scholar] [CrossRef] [PubMed]
- Dhanoa, B.S.; Cogliati, T.; Satish, A.G.; A Bruford, E.; Friedman, J.S. Update on the Kelch-like (KLHL) gene family. Hum. Genom. 2013, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jipa, A.; Vedelek, V.; Merényi, Z.; Ürmösi, A.; Takáts, S.; Kovács, A.L.; Horváth, G.V.; Sinka, R.; Juhász, G. Analysis of Drosophila Atg8 proteins reveals multiple lipidation-independent roles. Autophagy 2021, 17, 2565–2575. [Google Scholar] [CrossRef]
- Rusten, T.E.; Vaccari, T.; Lindmo, K.; Rodahl, L.M.; Nezis, I.P.; Sem-Jacobsen, C.; Wendler, F.; Vincent, J.-P.; Brech, A.; Bilder, D.; et al. ESCRTs and Fab1 regulate distinct steps of autophagy. Curr. Biol. 2007, 17, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Randle, K.E.; Wu, L.P. Ird1 is a Vps15 homologue important for antibacterial immune responses in Drosophila. Cell Microbiol. 2007, 9, 1073–1085. [Google Scholar] [CrossRef]
- Tsun, Z.-Y.; Bar-Peled, L.; Chantranupong, L.; Zoncu, R.; Wang, T.; Kim, C.; Spooner, E.; Sabatini, D.M. The folliculin tumor suppressor is a GAP for the RagC/D GTPases that signal amino acid levels to mTORC1. Mol. Cell 2013, 52, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Yuan, C.; Chen, R.; Dawson, T.M.; Dawson, V.L. ArfGAP1 Is a GTPase Activating protein for LRRK2: Reciprocal regulation of ArfGAP1 by LRRK2. J. Neurosci. 2012, 32, 3877–3886. [Google Scholar] [CrossRef] [Green Version]
- Nahm, M.; Lee, M.; Baek, S.H.; Yoon, J.H.; Kim, H.H.; Lee, Z.H.; Lee, S. Drosophila RhoGEF4 encodes a novel RhoA-specific guanine exchange factor that is highly expressed in the embryonic central nervous system. Gene 2006, 384, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Ringrose, L.; Rehmsmeier, M.; Dura, J.-M.; Paro, R. Genome-wide prediction of polycomb/trithorax response elements in Drosophila melanogaster. Dev. Cell 2003, 5, 759–771. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Forms | UniProt ID | Gene | Organism | Proteins | Modules |
|---|---|---|---|---|---|
| Fungi | A0A2Z6RA14_9GLOM | RclHR1_00310010 | Rhizophagus clarus | Uncharacterized protein | RhoGEF; PH; FYVE |
| A0A1Y2IJD5_PYCCO | PYCCODRAFT_557045 | Trametes coccinea | FYVE domain-containing protein | FYVE | |
| A0A1Y1YYR8_9FUNG | K493DRAFT_311810 | Basidiobolusmeristosporus | Uncharacterized protein | PH; FYVE | |
| A0A2T2PC27_CORCC | BS50DRAFT_567895 | Corynespora cassiicola | Vacuolar protein sorting-associated protein 27 | VHS; FYVE; UIM | |
| A0A507BWX7_9FUNG | SmJEL517_g06146 | Synchytrium microbalum | alpha-1,2-Mannosidase | RhoGEF; Glycosyl hydrolase family 47 | |
| A0A197JU28_9FUNG | K457DRAFT_126461 | Linnemannia elongate | Uncharacterized protein | RhoGEF; FYVE | |
| F8Q904_SERL3 | SERLA73DRAFT_170932 | Serpula lacrymans | Uncharacterized protein | RhoGEF; FYVE | |
| A0A1Y1UY75_9FUNG | BCR36DRAFT_362273 | Piromyces finnis | Ankyrin | Ankyrin repeats; FYVE; Ring finger | |
| Plants | A0A015IGQ3_RHIIW | RirG_217080 | Rhizophagus irregularis | Rom2p | RhoGEF; PH; FYVE |
| A0A177WRZ8_BATDL | BDEG_26098 | Batrachochytrium dendrobatidis | Uncharacterized protein | RhoGEF; PH | |
| F4P650_BATDJ | BATDEDRAFT_35443 | Batrachochytrium dendrobatidis | Uncharacterized protein | RhoGEF; FYVE | |
| A0A507EFA4_9FUNG | PhCBS80983_g00823 | Powellomyces hirtus | Uncharacterized protein | RhoGEF; PH; FYVE | |
| A0A1Y2IJD5_PYCCO | PYCCODRAFT_557045 | Trametes coccinea | FYVE domain-containing protein | FYVE | |
| A0A6A6HW15_9PLEO | BU26DRAFT_494685 | Trematosphaeria pertusa | Vacuolar protein sorting-associated protein 27 | VHS; FYVE; UIM | |
| A0A6A6V864_9PLEO | M011DRAFT_469719 | Sporormia fimetaria | Vacuolar protein sorting-associated protein 27 | FYVE; UIM | |
| tr|A0A0D7A3M6|A0A0D7A3M6_9AGAR | FISHEDRAFT_67225 | Fistulina fimetaria | Uncharacterized protein | RhoGEF; FYVE | |
| tr|A0A1Y1UY75|A0A1Y1UY75_9FUNG | BCR36DRAFT_362273 | Piromyces finnis | Ankyrin | Ankyrin repeats; FYVE; Ring finger |
| UniProt ID | Gene | Organism | Protein | Modules |
|---|---|---|---|---|
| G3RXJ3_GORGO | PLEKHF2 | Gorilla gorilla | Pleckstrin homology and FYVE domain-containing 2 | PH; FYVE; polyD |
| F6Q9Z2_MONDO | NDUFAF6 | Monodelphis domestica | Uncharacterized protein | Squalene/phytoene synthase; PH; FYVE, polyD |
| A0A6J0AU76_VICPA | PLEKHF1 | Vicugna pacos | Pleckstrin homology domain-containing family F member 1 | PH; FYVE; polyD |
| A0A4X2MDN9_VOMUR | FGD6 | Vombatus ursinus | FYVE, RhoGEF and PH domain-containing 6 | RhoGEF; PH; FYVE |
| PKHF2_HUMAN | PKHF2 | Homo sapiens | Pleckstrin homology domain-containing family F member 2 | PH; FYVE; polyD |
| K7ELB8_HUMAN | PLEKHF1 | Homo sapiens | Pleckstrin homology domain-containing family F member 1 | PH; FYVE; polyD |
| A4FVC4_HUMAN | FGD6 | Homo sapiens | FGD6 protein | RhoGEF; PH; FYVE |
| G3V2D8_HUMAN | ZFYVE26 | Homo sapiens | Zinc finger FYVE domain-containing protein 26 | FYVE |
| RUFY2_HUMAN | RUFY2 | Homo sapiens | RUN and FYVE domain-containing protein 2 | RUN; FYVE |
| ZFYV9_HUMAN | ZFYVE9 | Homo sapiens | Zinc finger FYVE domain-containing protein 9 | FYVE; SARA; DUF |
| FARP1_HUMAN | FARP1 | Homo sapiens | FERM, ARHGEF, and pleckstrin domain-containing protein 1 | FERM N-terminal; FERM central; FERM C-terminal PH-like; FERM adjacent (FA); RhoGEF; PH |
| G5E953_HUMAN | MTMR3 | Homo sapiens | Phosphatidylinositol-3-phosphate phosphatase | Myotubularin-like phosphatase; FYVE |
| WDFY1_HUMAN | WDFY1 | Homo sapiens | WD repeat and FYVE domain-containing protein 1 | WD, G-b repeat; FYVE |
| A0A7I2YQD1_HUMAN | HGS | Homo sapiens | Hepatocyte growth factor-regulated tyrosine kinase substrate | VHS; FYVE; hepatocyte growth factor-regulated tyrosine kinase substrate |
| ANFY1_HUMAN | ANKFY1 | Homo sapiens | Rabankyrin-5 | Ankyrin repeats; FYVE |
| ZFY21_HUMAN | ZFYVE21 | Homo sapiens | Zinc finger FYVE domain-containing protein 21 | Zinc finger FYVE-containing protein 21 C-terminus |
| FYV1_HUMAN | PIKFYVE | Homo sapiens | 1-phosphatidylinositol 3-phosphate 5-kinase | FYVE; DEP; TCP-1/cpn60 chaperonin family; PtdIns4P 5-kinase |
| A7E293_HUMAN | WDFY3 | Homo sapiens | WDFY3 protein | PH domain associated with BEACH; BEACH; WD; G-b repeat; FYVE |
| RBNS5_HUMAN | RBSN | Homo sapiens | Rabenosyn-5 | FYVE; Rabenosyn Rab binding; Rabenosyn-5 repeating NPF sequence-motif |
| CYH4_HUMAN | CYTH4 | Homo sapiens | Cytohesin-4 | Sec7; PH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, M.; Capelluto, D.G.S. The PH Domain and C-Terminal polyD Motif of Phafin2 Exhibit a Unique Concurrence in Animals. Membranes 2022, 12, 696. https://doi.org/10.3390/membranes12070696
Hasan M, Capelluto DGS. The PH Domain and C-Terminal polyD Motif of Phafin2 Exhibit a Unique Concurrence in Animals. Membranes. 2022; 12(7):696. https://doi.org/10.3390/membranes12070696
Chicago/Turabian StyleHasan, Mahmudul, and Daniel G. S. Capelluto. 2022. "The PH Domain and C-Terminal polyD Motif of Phafin2 Exhibit a Unique Concurrence in Animals" Membranes 12, no. 7: 696. https://doi.org/10.3390/membranes12070696