
Protein Kinase C Regulates Expression and Function of the Cav3.2 T-Type Ca2+ Channel during Maturation of Neonatal Rat Cardiomyocyte

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Neonatal Rat Cardiomyocytes and Cell Culture
2.2. Stable Cell Lines for Cav3.1 and Cav3.2 Channels
2.3. Electrophysiology
2.4. Solutions and Chemicals
2.5. Quantitative Real-Time PCR
2.6. Preparation of Recombinant Adenoviruses
2.7. Measurement of PKC Activation
2.8. Action Potential Parameters
2.9. Data Acquisition and Statistical Analysis
3. Results
3.1. Nickel as a Selective Inhibitor of Cav3.2 T-Type Ca2+ Channel
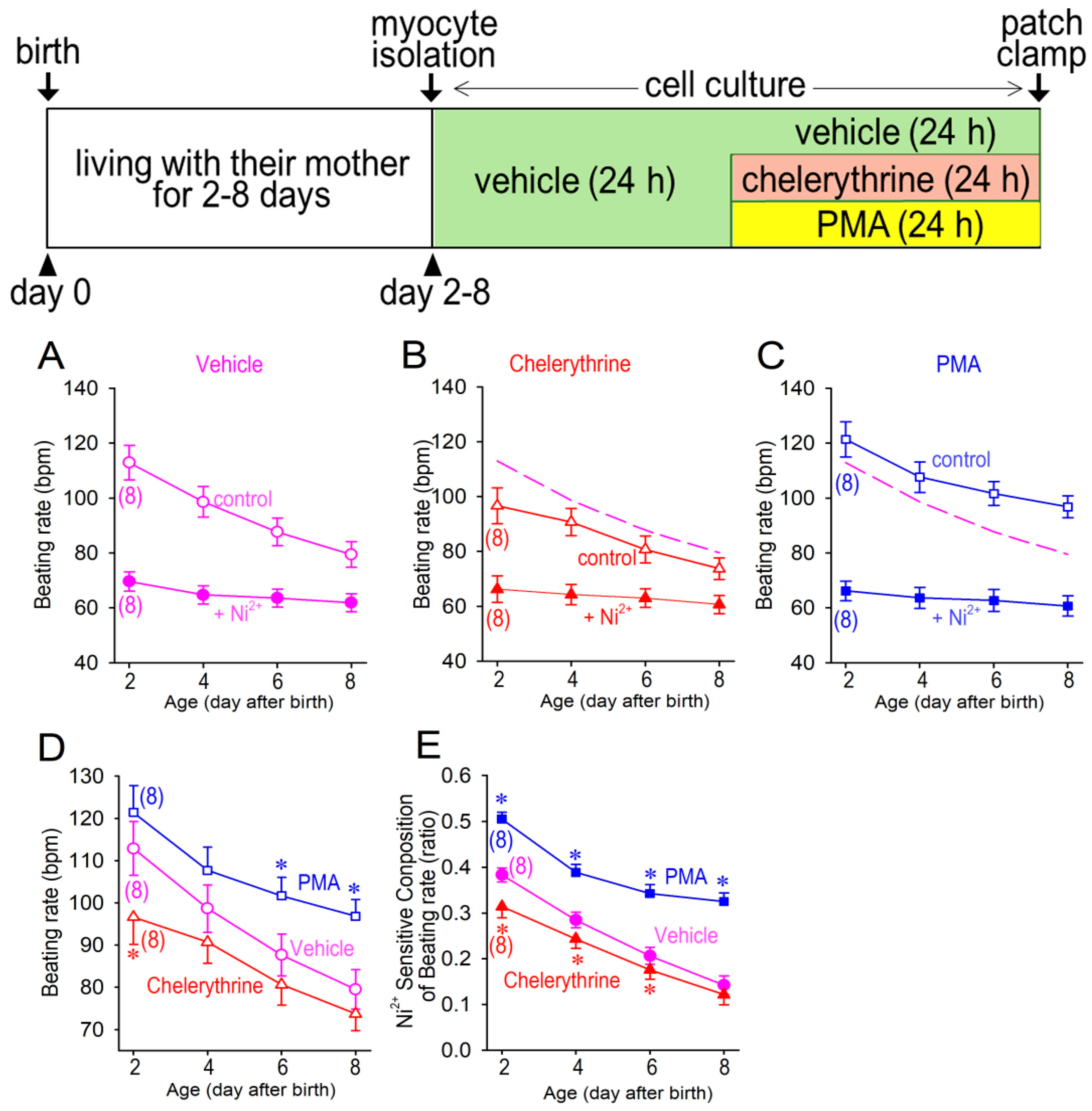
3.2. Cellular Automaticity in Neonatal Cardiomyocytes
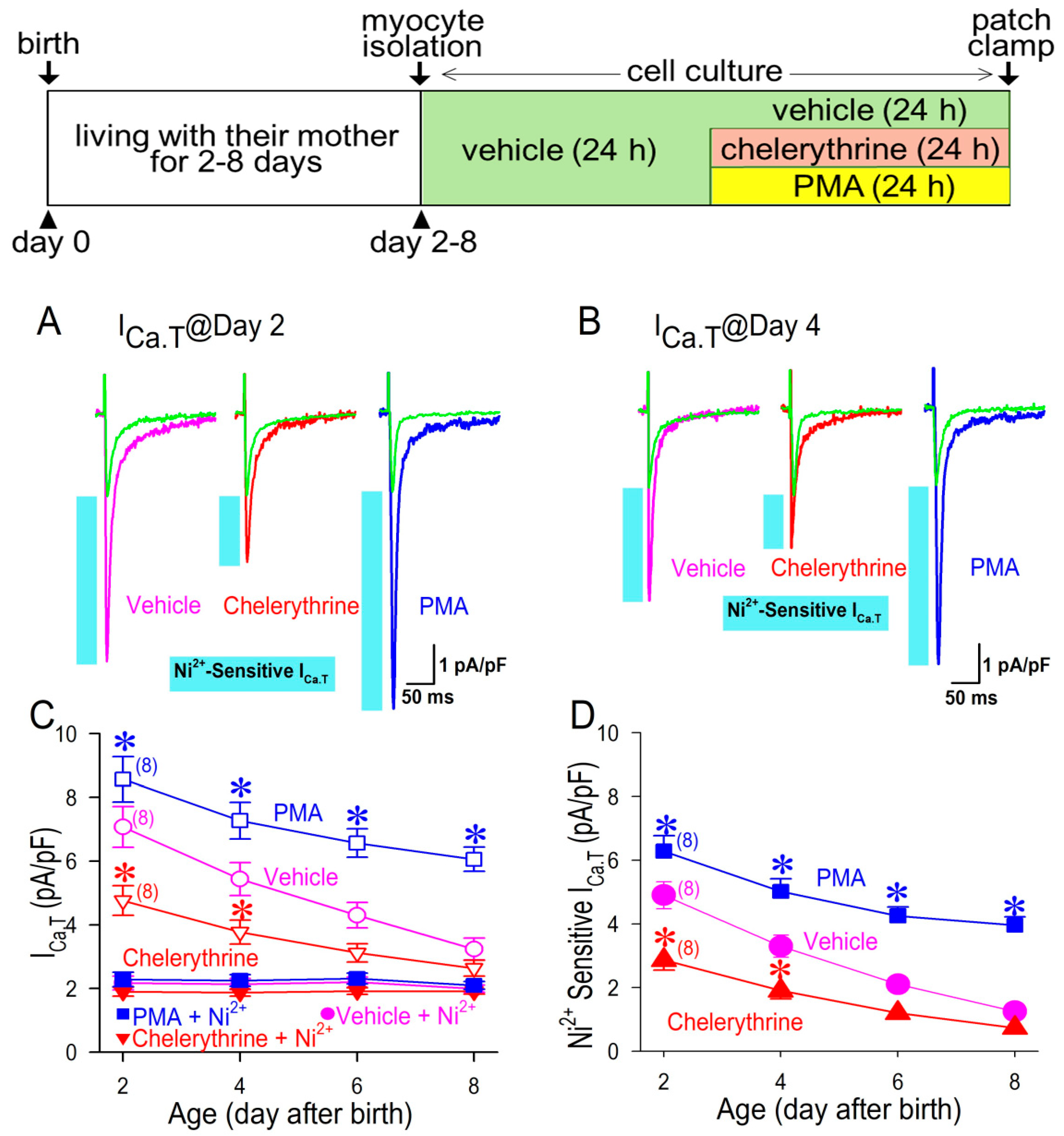
3.3. Ni2+-Sensitive and -Insensitive T-Type Ca2+ Current in Neonatal Cardiomyocytes
3.4. PKC Upregulates CaV3.2 mRNA
3.5. Cav3.1 Expression Is Negatively Associated with PKC Activity
3.6. Transcription Factor Nkx2.5 Is Responsible for PKC Actions for Cav3.2
4. Discussion
4.1. Isoform Switching of the T-Type Ca2+ Channels in the Neonatal Heart
4.2. Actions of PKC for the Cac3.2 Channel
4.3. Study Limitation
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ICa.T | T-type Ca2+ channel current |
| ICa.L | L-type Ca2+ channel current |
| MDP | maximum diastolic potentials |
| DDR | diastolic depolarization rate |
| Vmax | maximum dV/dt of action potentials |
| OS | overshoot of action potentials |
| TOP | take-off potential |
| APD50 | action potential duration at 50% repolarization |
| PKC | protein kinase C |
| Nkx2.5 | NK-2 transcription factor related, locus 5 |
| DAG | diacylglycerol |
| Ni2+ | nickle ion |
| HEK | human embryonic kidney |
| PCR | polymerase chain reaction |
| mRNA | messenger ribonucleic acid |
| cDNA | complementary deoxyribonucleic acid |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| IC50 | half maximal (50%) inhibitory concentration |
| h | hour |
| Ad-Nkx | recombinant adenovirus encoding NK-2 transcription factor related, locus 5 |
| Ad-GFP | recombinant adenovirus encoding green fluorescent protein |
| DMEM | Dulbecco’s modified Eagle’s medium |
| PMA | phorbol 12-myristate 13-acetate |
| Gö6976 | 5,6,7,13-tetrahydro-13-methyl-5-oxo-12H-indolo[2,3-a]pyrrolo[3,4-c]carbazole-12-propanenitrile |
| Ro-32-0432 | (3-(8-((dimethylamino)methyl)-6,7,8,9-tetrahydropyrido [1,2-a]indol-10-y1)-4-(1-methyl-1H-indol-3-y1)-1H-pyrrole-2,5-dione hydrochloride) |
| IYIAP | 5,6-Bis[(4-fluorophenyl)amino]-2H-isoindole-1,3-dione |
| CGP41251 | 4′-N-benzoyl staurosporine |
| IYIAP | 3-anilino-4-[1-(3-imidazol-1-ylpropyl)indol-3-yl]pyrrole-2,5-dione |
| Rsqr | the coefficient of determination |
| iPS: | induced pluripotent stem |
References
- Mesirca, P.; Torrente, A.G.; Mangoni, M.E. Functional role of voltage gated Ca2+ channels in heart automaticity. Front. Physiol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takebayashi, S.; Li, Y.; Kaku, T.; Inagaki, S.; Hashimoto, Y.; Kimura, K.; Miyamoto, S.; Hadama, T.; Ono, K. Remodeling excitation-contraction coupling of hypertrophied ventricular myocytes is dependent on T-type calcium channels expression. Biochem. Biophys. Res. Commun. 2006, 345, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Berthier, C.; Monteil, A.; Lory, P.; Strube, C. α1H mRNA in single skeletal muscle fibres accounts for T-type calcium current transient expression during fetal development in mice. J. Physiol. 2002, 539, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.K.; Mitchell, J.W.; Best, P.M. Quantitative analysis of the expression and distribution of calcium channel α1 subunit mRNA in the atria and ventricles of the rat heart. J. Mol. Cell. Cardiol. 2002, 34, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Wang, Y.; Kang, L.; Shimaoka, T.; Marni, F.; Ono, K. Intracellular Ca2+- and PKC-dependent upregulation of T-type Ca2+ channels in LPC-stimulated cardiomyocytes. J. Mol. Cell. Cardiol. 2010, 48, 131–139. [Google Scholar] [CrossRef]
- Kline, R.; Jiang, T.; Rybin, V.O.; Steinberg, S.F. Abnormal Ca and εPKC signaling in hypertrophied atrial tumor myocytes (AT-1 cells). Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1761–H1769. [Google Scholar] [CrossRef] [Green Version]
- Mende, U.; Kagen, A.; Meister, M.; Neer, E.J. Signal transduction in atria and ventricles of mice with transient cardiac expression of activated G protein αq. Circ. Res. 1999, 85, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.A.; Mochly-Rosen, D. Inhibition of the spontaneous rate of contraction of neonatal cardiac myocytes by protein kinase C isozymes. Circ. Res. 1995, 76, 654–663. [Google Scholar] [CrossRef]
- Marrocco, V.; Bogomolovas, J.; Ehler, E.; Dos Remedios, C.G.; Yu, J.; Gao, C.; Lange, S. PKC and PKN in heart disease. J. Mol. Cell. Cardiol. 2019, 128, 212–226. [Google Scholar] [CrossRef] [Green Version]
- Morishima, M.; Fujita, T.; Osagawa, S.; Kubota, H.; Ono, K. Enhanced BDNF actions following acute hypoxia facilitate HIF-1α-dependent upregulation of Cav3-T-type Ca2+ channels in rat cardiomyocytes. Membranes 2021, 11, 470. [Google Scholar] [CrossRef]
- Morishima, M.; Tahara, S.; Wang, Y.; Ono, K. Oxytocin downregulates the CaV1.2 L-type Ca2+ channel via Gi/cAMP/PKA/CREB signaling pathway in cardiomyocytes. Membranes 2021, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Uchino, T.; Kaku, T.; Kang, L.; Wang, Y.; Takebayashi, S.; Ono, K. Lysophosphatidylcholine augments CaV3.2 but not CaV3.1 T-type Ca2+ channel current expressed in HEK-293 cells. Pharmacology 2006, 76, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Kaku, T.; Uchino, T.; Isomoto, S.; Yoshimatsu, H.; Ono, K. Short- and long-term amiodarone treatments regulate Cav3.2 low-voltage-activated T-type Ca2+ channel through distinct mechanisms. Mol. Pharmacol. 2006, 69, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Kohl, R.; Preiss, S.; von Knethen, A.; Brune, B. Oxidized low-density lipoprotein depletes PKCα and attenuates reactive oxygen species formation in monocytes/macrophages. Cardiovasc. Res. 2006, 71, 574–585. [Google Scholar] [CrossRef]
- Hambleton, M.; Hahn, H.; Pleger, S.T.; Kuhn, M.C.; Klevitsky, R.; Carr, A.N.; Kimball, T.F.; Hewett, T.E.; Dorn, G.W., II; Koch, W.J.; et al. Pharmacological- and gene therapy-based inhibition of protein kinase Cα/β enhances cardiac contractility and attenuates heart failure. Circulation 2006, 114, 574–582. [Google Scholar] [CrossRef]
- Sampson, S.R.; Lupowitz, Z.; Braiman, L.; Zisapel, N. Role of protein kinase Cα in melatonin signal transduction. Mol. Cell. Endocrinol. 2006, 252, 82–87. [Google Scholar] [CrossRef]
- Kouroedov, A.; Eto, M.; Joch, H.; Volpe, M.; Luscher, T.F.; Cosentino, F. Selective inhibition of protein kinase Cβ2 prevents acute effects of high glucose on vascular cell adhesion molecule-1 expression in human endothelial cells. Circulation 2004, 110, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Morishima, M.; Zheng, M.; Uchino, T.; Mannen, K.; Takahashi, A.; Nakaya, Y.; Komuro, I.; Ono, K. Transcription factors Csx/Nkx2.5 and GATA4 distinctly regulate expression of Ca2+ channels in neonatal rat heart. J. Mol. Cell. Cardiol. 2007, 42, 1045–1053. [Google Scholar] [CrossRef]
- Wang, Y.; Morishima, M.; Li, D.; Takahashi, N.; Saikawa, T.; Nattel, S.; Ono, K. Binge alcohol exposure triggers atrial fibrillation through T-type Ca2+ channel upregulation via protein kinase C (PKC)/glycogen synthesis kinase 3β (GSK3β)/nuclear factor of activated T-cells (NFAT) signaling—An experimental account of holiday heart syndrome. Circ. J. 2020, 84, 1931–1940. [Google Scholar]
- Panáková, D.; Werdich, A.A.; Macrae, C.A. Wnt11 patterns a myocardial electrical gradient through regulation of the L-type Ca2+ channel. Nature 2010, 466, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, E.; Shirai, M.; Arakawa, K.; Hidaka, K.; Miake, J.; Ninomiya, H.; Kato, M.; Shigemasa, C.; Shirayoshi, Y.; Hisatome, I.; et al. Different distribution of Cav3.2 and Cav3.1 transcripts encoding T-type Ca2+ channels in the embryonic heart of mice. Biomed. Res. 2010, 31, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cribbs, L.L.; Lee, J.H.; Yang, J.; Satin, J.; Zhang, Y.; Daud, A.; Barclay, J.; Williamson, M.P.; Fox, M.; Rees, M.; et al. Cloning and characterization of α1H from human heart, a member of the T-type Ca2+ channel gene family. Circ. Res. 1998, 83, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manabe, K.; Miake, J.; Sasaki, N.; Furuichi, H.; Yano, S.; Mizuta, E.; Yamamoto, Y.; Hoshikawa, Y.; Yamazaki, H.; Tajima, F.; et al. Developmental changes of Ni2+ sensitivity and automaticity in Nkx2.5-positive cardiac precursor cells from murine embryonic stem cell. Circ. J. 2004, 68, 724–726. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, E.; Miake, J.; Yano, S.; Furuichi, H.; Manabe, K.; Sasaki, N.; Igawa, O.; Hoshikawa, Y.; Shigemasa, C.; Nanba, E.; et al. Subtype switching of T-type Ca2+ channels from Cav3.2 to Cav3.1 during differentiation of embryonic stem cells to cardiac cell lineage. Circ. J. 2005, 69, 1284–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.K.; Oyola, M.G. Progesterone signaling mechanisms in brain and behavior. Front. Endocrinol. 2012, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marni, F.; Wang, Y.; Morishima, M.; Shimaoka, T.; Uchino, T.; Zheng, M.; Kaku, T.; Ono, K. 17β-estradiol modulates expression of low-voltage-activated CaV3.2 T-type calcium channel via extracellularly regulated kinase pathway in cardiomyocytes. Endocrinology 2009, 150, 879–888. [Google Scholar] [CrossRef]
- Singh, R.M.; Cummings, E.; Pantos, C.; Singh, J. Protein kinase C and cardiac dysfunction: A review. Heart Fail. Rev. 2017, 22, 843–859. [Google Scholar] [CrossRef] [Green Version]
- Nishizuka, Y. Protein kinase C and lipid signaling for sustained cellular responses. FASEB J. 1995, 9, 484–496. [Google Scholar] [CrossRef]
- Goldberg, M.; Steinberg, S.F. Tissue-specific developmental regulation of protein kinase C isoforms. Biochem. Pharmacol. 1996, 51, 1089–1093. [Google Scholar] [CrossRef]
- Radaszkiewicz, K.A.; Beckerová, D.; Woloszczuková, L.; Radaszkiewicz, T.W.; Lesáková, P.; Blanářová, O.V.; Kubala, L.; Humpolíček, P.; Pachernik, J. 12-O-Tetradecanoylphorbol-13-acetate increases cardiomyogenesis through PKC/ERK signaling. Sci. Rep. 2020, 10, 15922. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Morishima, M.; Ono, K. Protein Kinase C Regulates Expression and Function of the Cav3.2 T-Type Ca2+ Channel during Maturation of Neonatal Rat Cardiomyocyte. Membranes 2022, 12, 686. https://doi.org/10.3390/membranes12070686
Wang Y, Morishima M, Ono K. Protein Kinase C Regulates Expression and Function of the Cav3.2 T-Type Ca2+ Channel during Maturation of Neonatal Rat Cardiomyocyte. Membranes. 2022; 12(7):686. https://doi.org/10.3390/membranes12070686
Chicago/Turabian StyleWang, Yan, Masaki Morishima, and Katsushige Ono. 2022. "Protein Kinase C Regulates Expression and Function of the Cav3.2 T-Type Ca2+ Channel during Maturation of Neonatal Rat Cardiomyocyte" Membranes 12, no. 7: 686. https://doi.org/10.3390/membranes12070686