
Investigating the Permeation Mechanism of Typical Phthalic Acid Esters (PAEs) and Membrane Response Using Molecular Dynamics Simulations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Simulation Details
2.1.1. System Preparation
2.1.2. Conventional MD Simulations (cMD)
2.1.3. Steered MD (SMD) Simulation and Umbrella Sampling
2.2. Trajectory Analysis
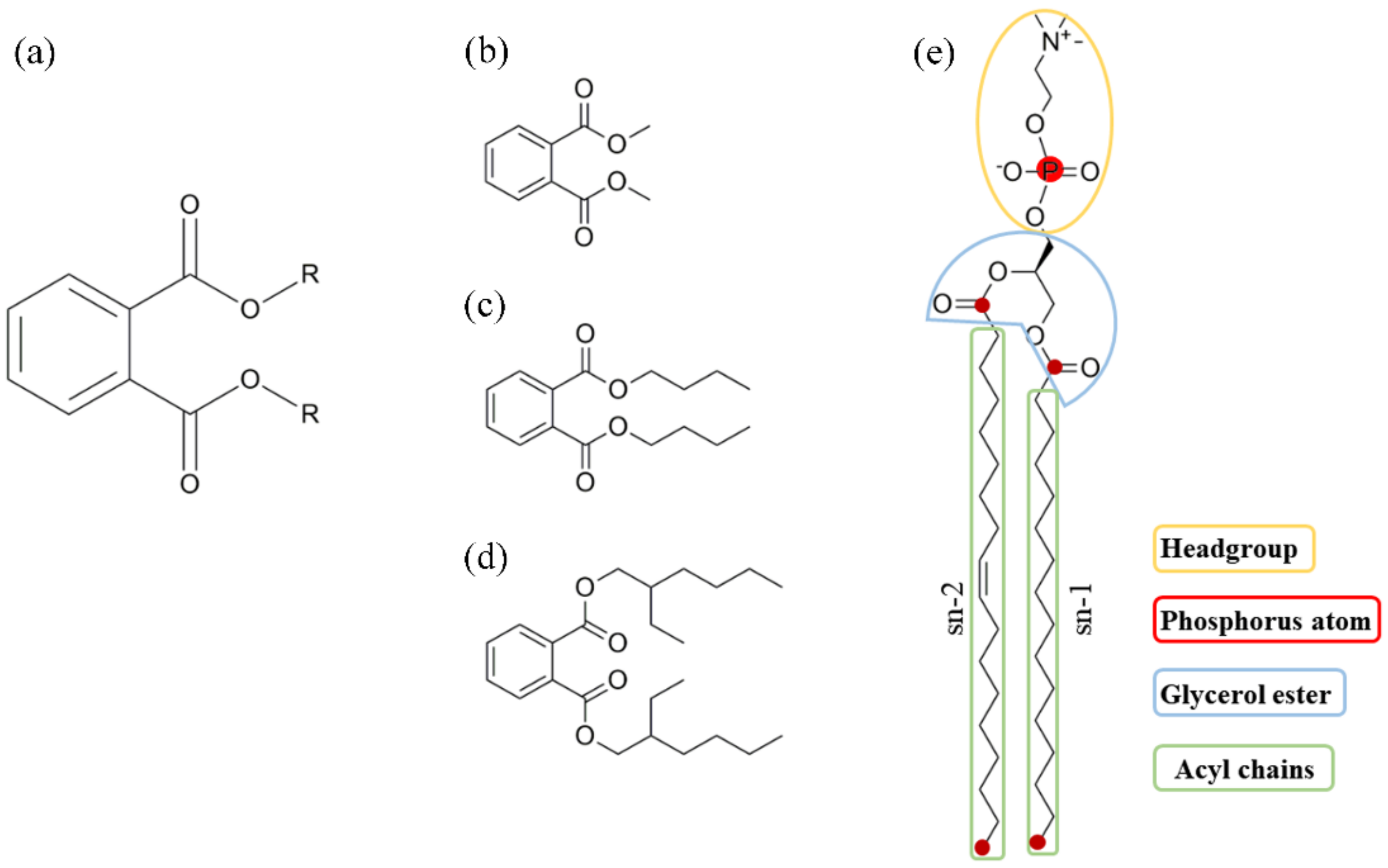
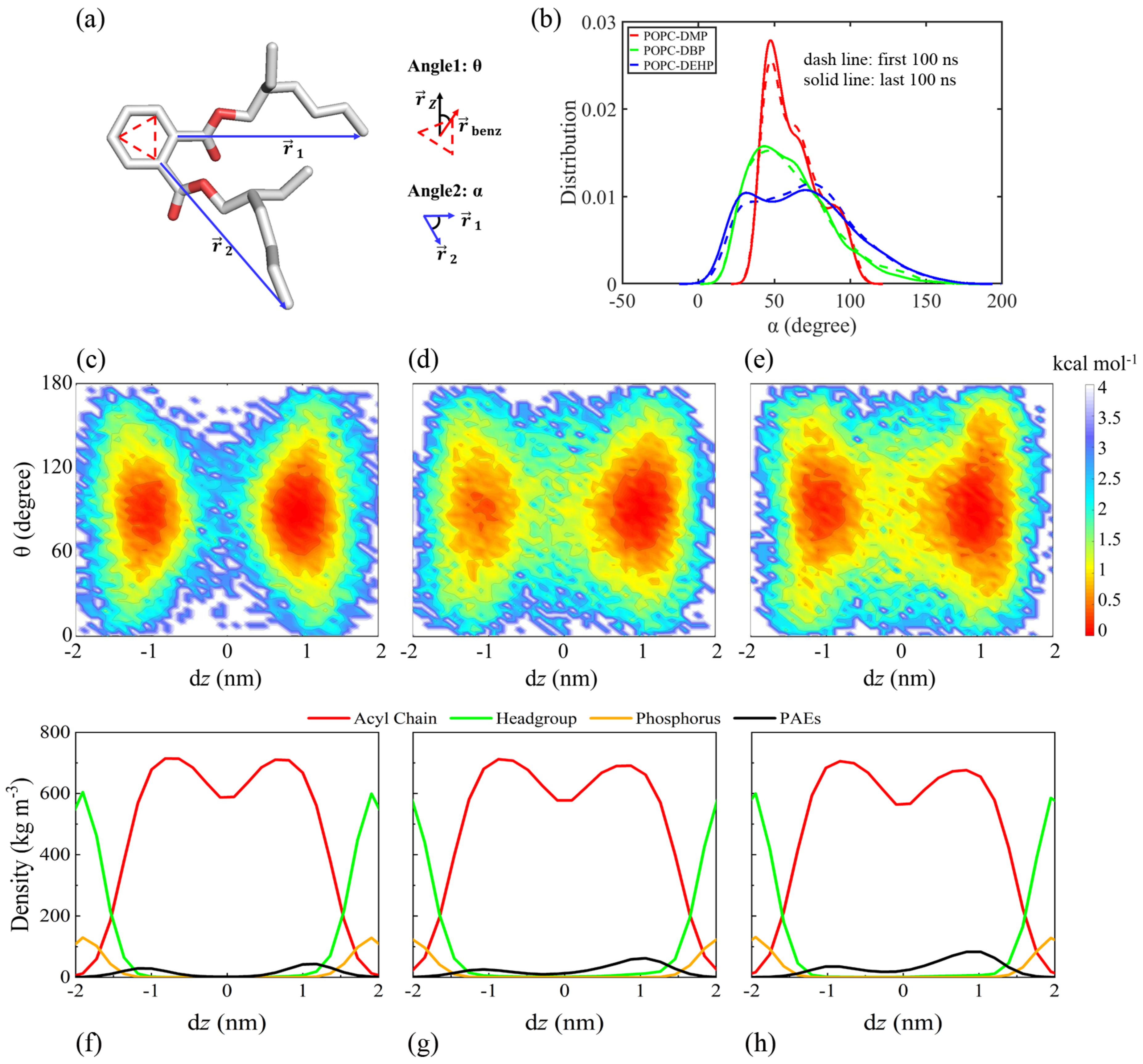
2.2.1. PAEs Configuration
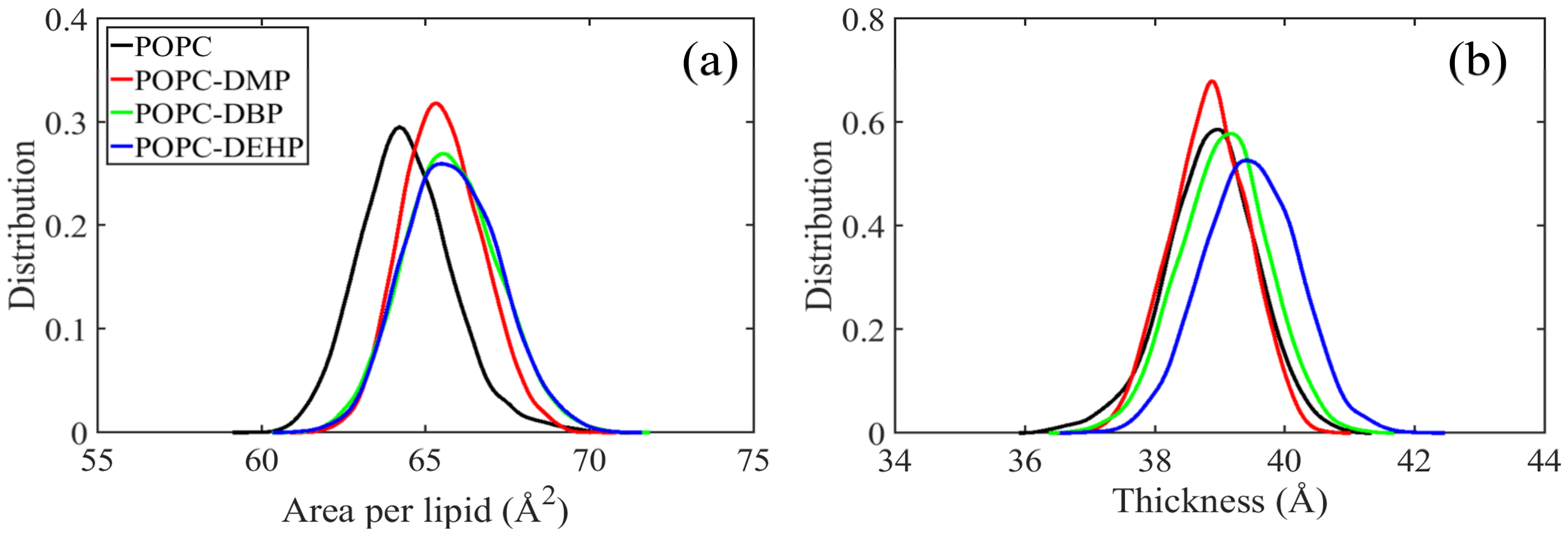
2.2.2. Area Per Lipid and Bilayer Thickness
2.2.3. Deuterium Order Parameters (Scd)
2.2.4. Lateral Diffusion
3. Results
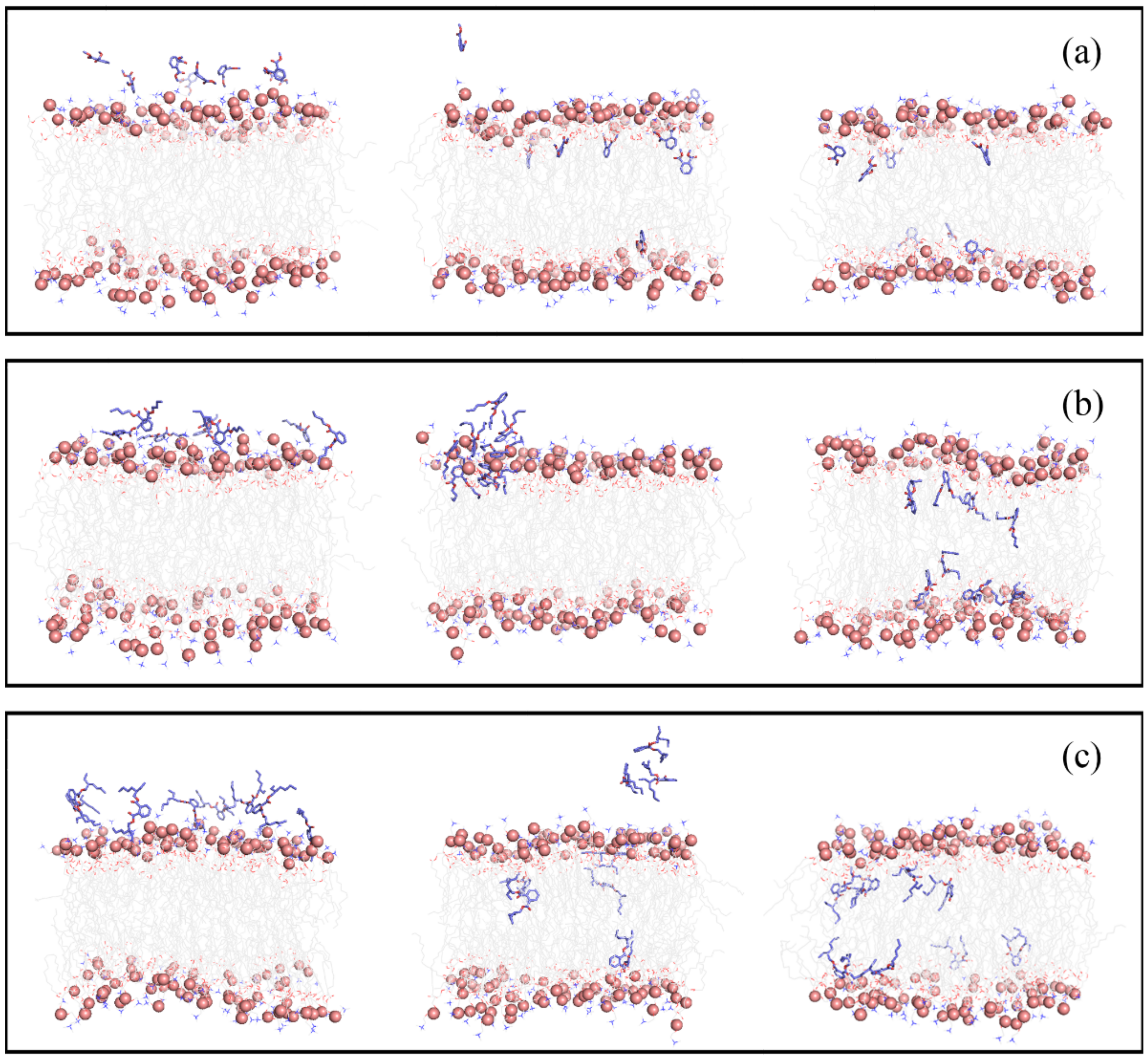
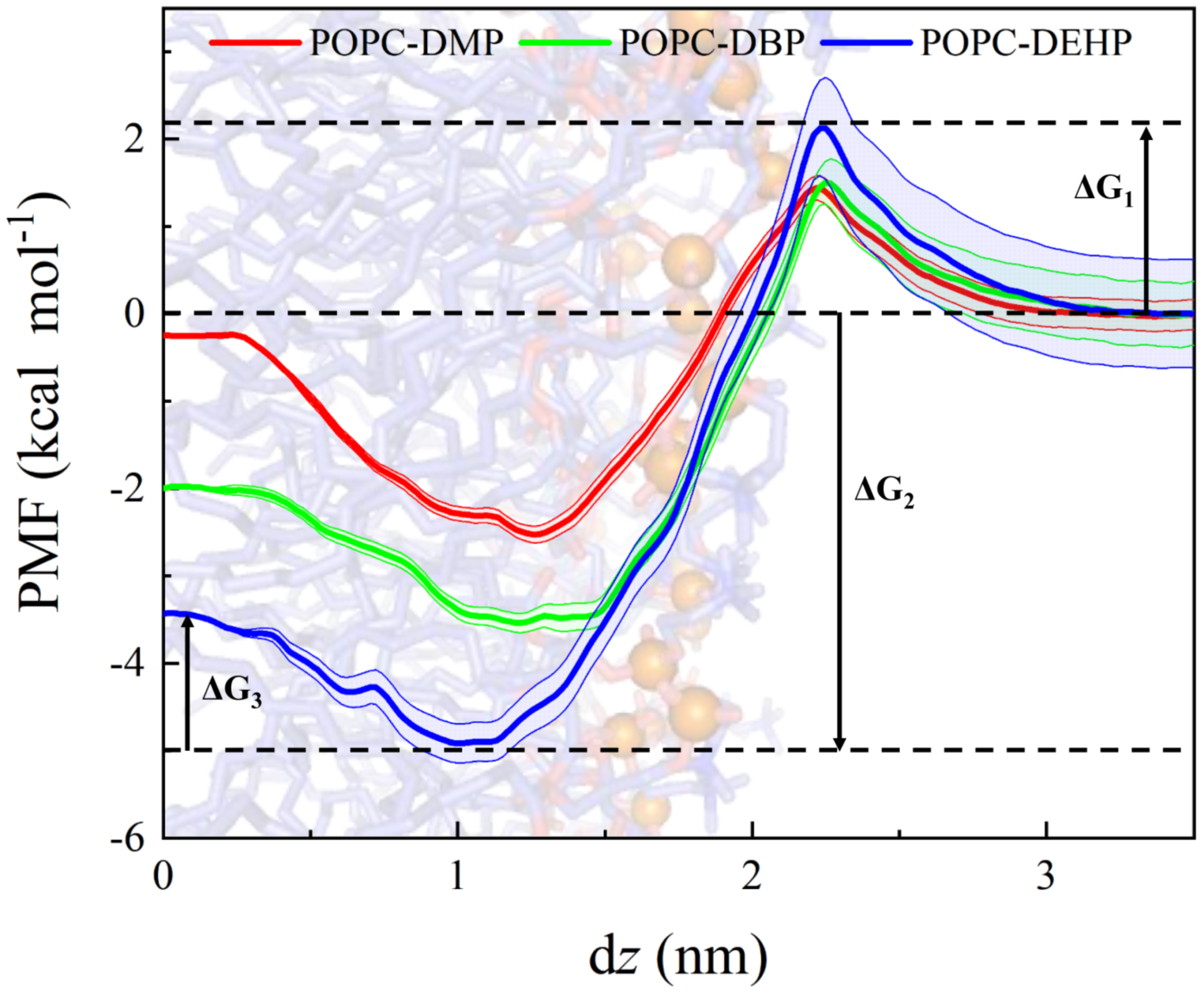
3.1. Permeation and Diffusion of Different Types of PAEs through the POPC Bilayer
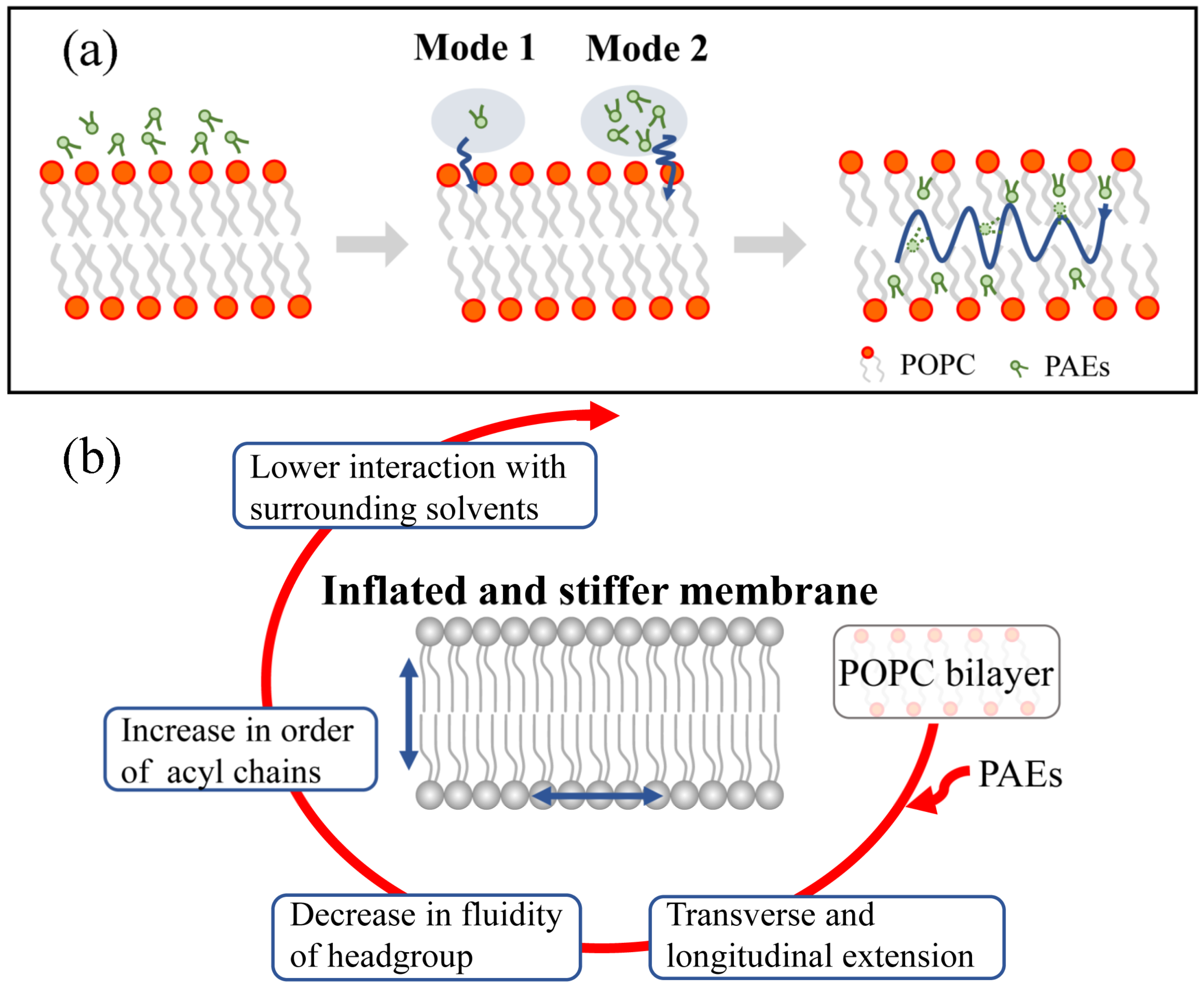
3.2. Partitioning of PAEs into the Lipid Bilayer Affects the Membrane Properties
3.2.1. PAEs Penetration Induces Transverse and Longitudinal Extension of Lipid Membranes
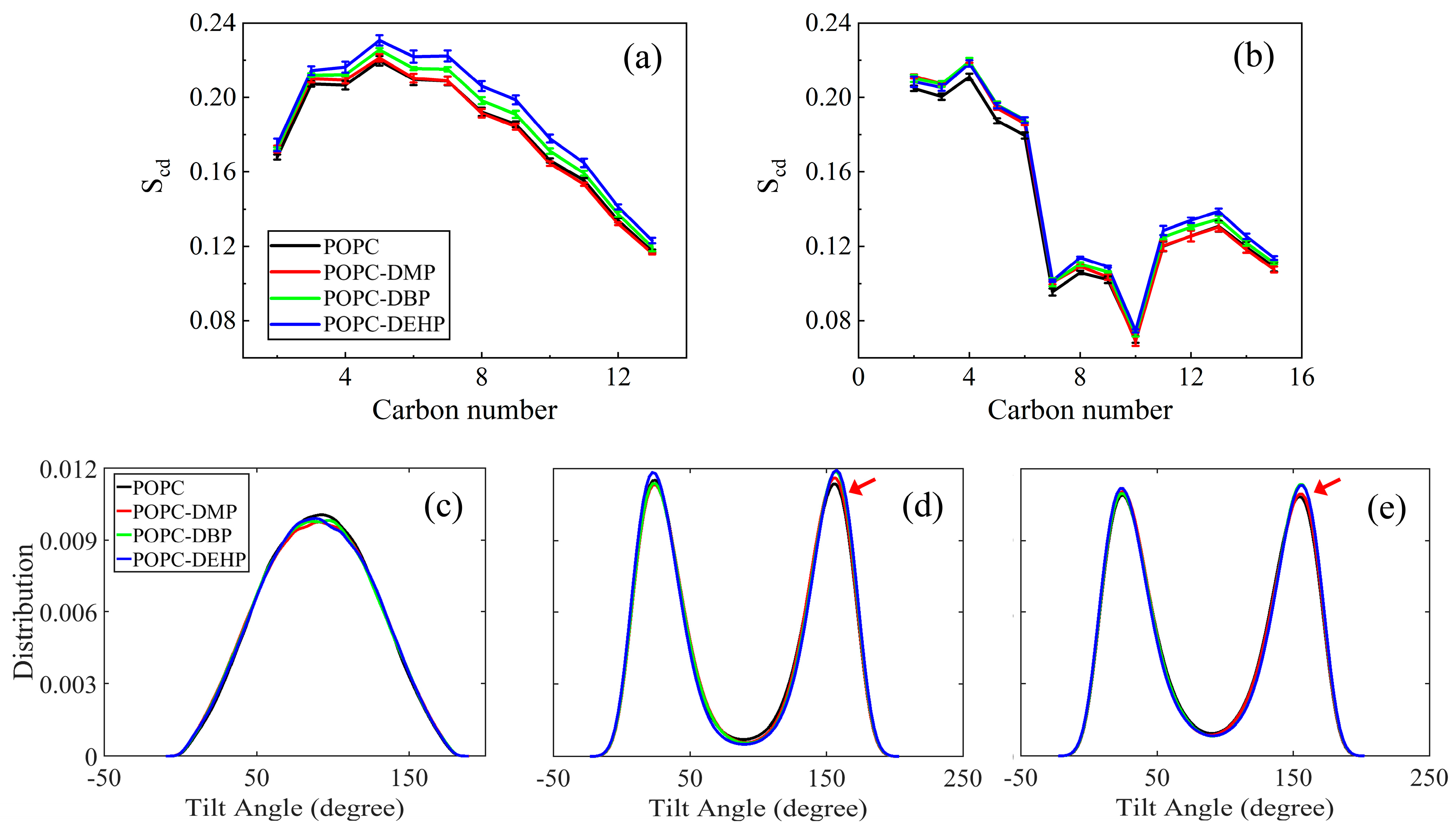
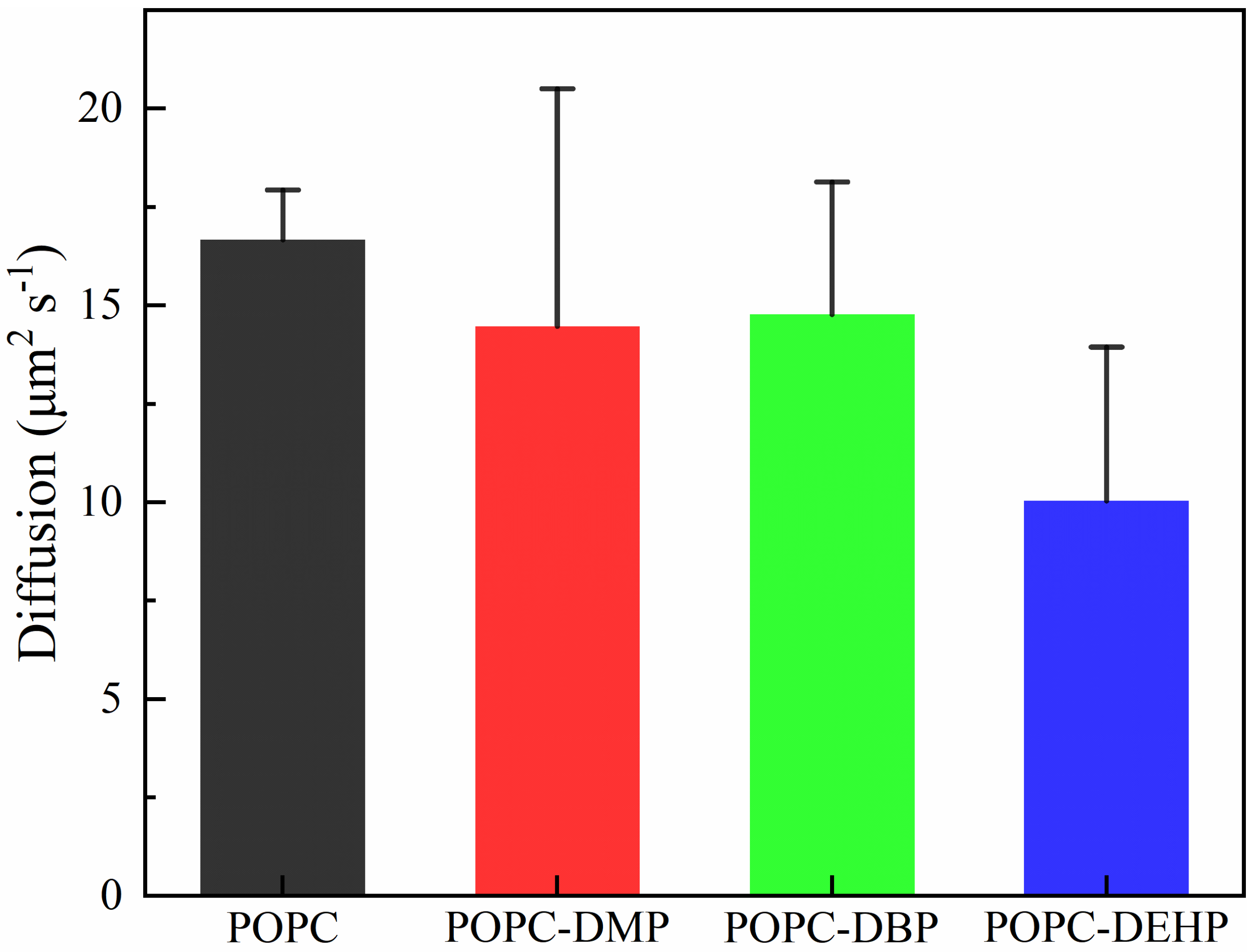
3.2.2. PAEs Insertion Reduces Lipid Fluidity
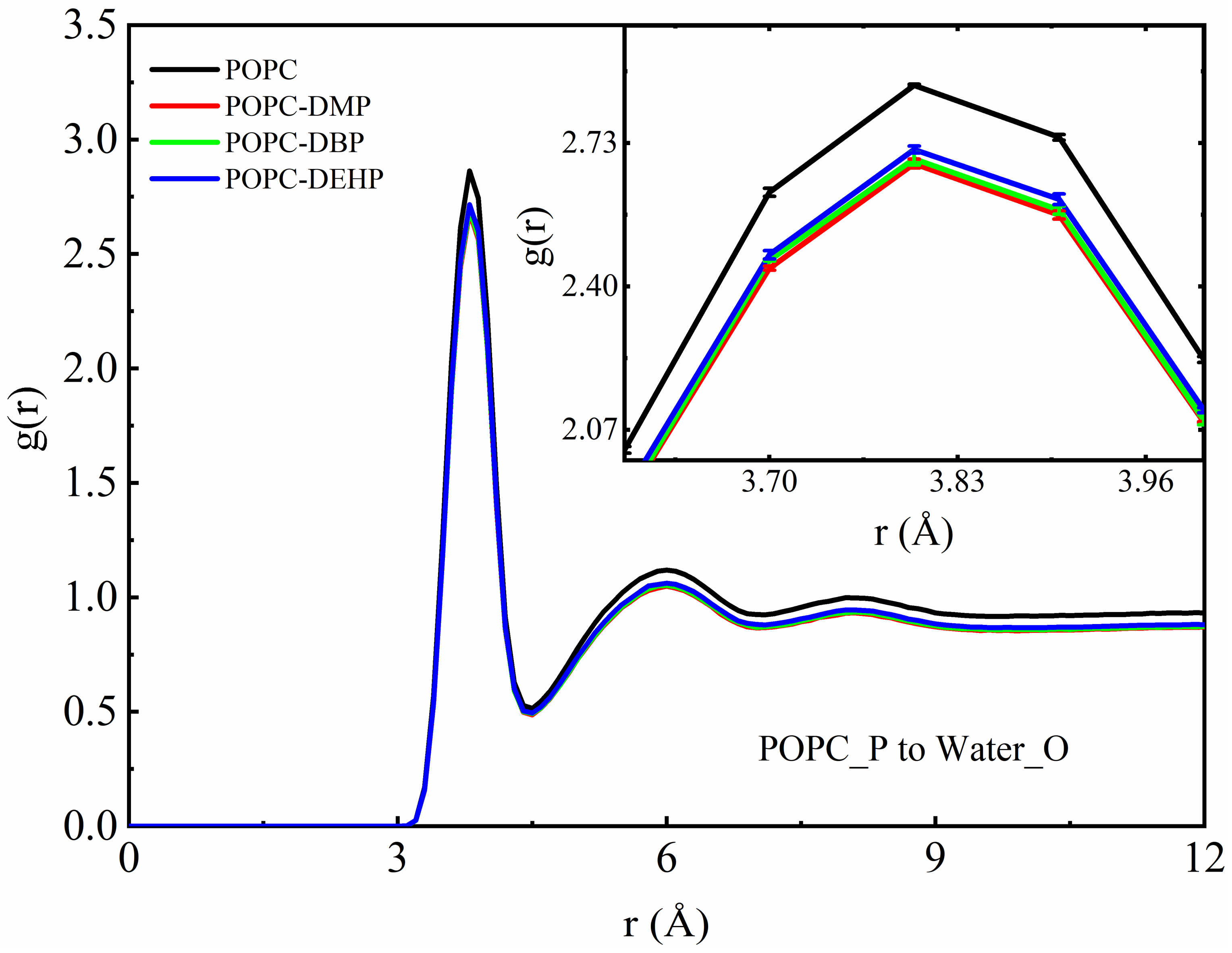
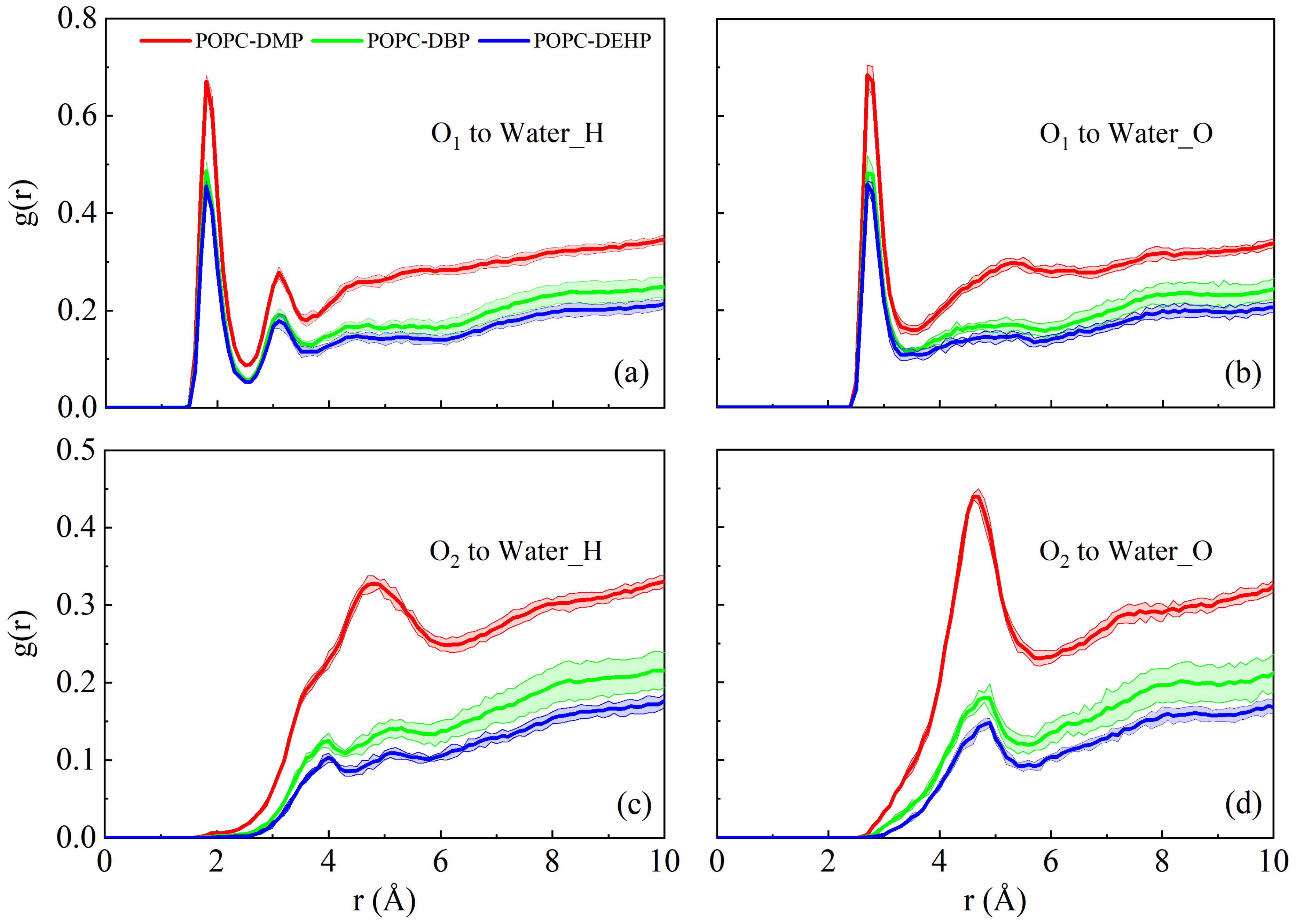
3.2.3. PAEs Decrease the Interaction of POPC Phosphate Groups with Surrounding Solvents
3.3. Membrane Permeability of PAEs Negatively Correlates with Their Side-Chain Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rongqi, C. Harmfulness of Phthalic Acid Ester Plasticizers to Human Health. Dyest. Color. 2015, 52, 52–58. [Google Scholar]
- Gao, D.W.; Wen, Z.D. Phthalate esters in the environment: A critical review of their occurrence, biodegradation, and removal during wastewater treatment processes. Sci. Total Environ. 2016, 541, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Kimber, I.; Dearman, R.J. An assessment of the ability of phthalates to influence immune and allergic responses. Toxicology 2010, 271, 73–82. [Google Scholar] [CrossRef]
- Fu, P.; Kawamura, K.; Barrie, L.A. Photochemical and Other Sources of Organic Compounds in the Canadian High Arctic Aerosol Pollution during Winter-Spring. Environ. Sci. Technol. 2009, 43, 286–292. [Google Scholar] [CrossRef]
- Dargnat, C.; Teil, M.J.; Chevreuil, M.; Blanchard, M. Phthalate removal throughout wastewater treatment plant: Case study of Marne Aval station (France). Sci. Total Environ. 2009, 407, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Net, S.; Dumoulin, D.; El-Osmani, R.; Rabodonirina, S.; Ouddane, B. Case study of PAHs, Me-PAHs, PCBs, Phthalates and Pesticides Contamination in the Somme River water, France. Int. J. Environ. Res. 2014, 8, 1159–1170. [Google Scholar]
- Gao, D.; Li, Z.; Wen, Z.; Ren, N. Occurrence and fate of phthalate esters in full-scale domestic wastewater treatment plants and their impact on receiving waters along the Songhua River in China. Chemosphere 2014, 95, 24–32. [Google Scholar] [CrossRef]
- Wang, X.; Tao, W.; Xu, Y.; Feng, J.; Wang, F. Indoor phthalate concentration and exposure in residential and office buildings in Xi’an, China. Atmos. Environ. 2014, 87, 146–152. [Google Scholar] [CrossRef]
- Bergh, C.; Magnus Aberg, K.; Svartengren, M.; Emenius, G.; Ostman, C. Organophosphate and phthalate esters in indoor air: A comparison between multi-storey buildings with high and low prevalence of sick building symptoms. J. Environ. Monit. 2011, 13, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Akingbemi, B.T.; Ge, R.; Klinefelter, G.R.; Zirkin, B.R.; Hardy, M.P. Phthalate-induced Leydig cell hyperplasia is associated with multiple endocrine disturbances. Proc. Natl. Acad. Sci. USA 2003, 101, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Leung, A.O.W.; Chu, L.H.; Wong, M.H. Phthalates contamination in China: Status, trends and human exposure-with an emphasis on oral intake. Environ. Pollut. 2018, 238, 771–782. [Google Scholar] [CrossRef]
- Chi, Z.; Zhao, J.; You, H.; Wang, M. Study on the Mechanism of Interaction between Phthalate Acid Esters and Bovine Hemoglobin. J. Agric. Food Chem. 2016, 64, 6035–6041. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Z.; Nakanishi, T.; Wan, Y.; Hiromori, Y.; Nagase, H.; Hu, J. Structure-dependent activity of phthalate esters and phthalate monoesters binding to human constitutive androstane receptor. Chem. Res. Toxicol. 2015, 28, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, G.; Wang, L. Potential toxicity of phthalic acid esters plasticizer: Interaction of dimethyl phthalate with trypsin in vitro. J. Agric. Food Chem. 2015, 63, 75–84. [Google Scholar] [CrossRef]
- Chi, Z.; Zhao, J.; Li, W.; Araghi, A.; Tan, S. In vitro assessment of phthalate acid esters-trypsin complex formation. Chemosphere 2017, 185, 29–35. [Google Scholar] [CrossRef]
- Lv, X.; Jiang, Z.; Zeng, G.; Zhao, S.; Li, N.; Chen, F.; Huang, X.; Yao, J.; Tuo, X. Comprehensive insights into the interactions of dicyclohexyl phthalate and its metabolite to human serum albumin. Food Chem. Toxicol. 2021, 155, 112407. [Google Scholar] [CrossRef]
- Wu, Z.; Yi, Z.; Dong, L.; Zhang, A. Molecular simulation study of the specific combination between four kinds of phthalic acid esters and human serum albumin. Environ. Toxicol. Pharmacol. 2016, 41, 259–265. [Google Scholar] [CrossRef]
- Sicinska, P. Di-n-butyl phthalate, butylbenzyl phthalate, and their metabolites exhibit different apoptotic potential in human peripheral blood mononuclear cells. Food Chem. Toxicol. 2019, 133, 110750. [Google Scholar] [CrossRef] [PubMed]
- Bider, R.C.; Lluka, T.; Himbert, S.; Khondker, A.; Qadri, S.M.; Sheffield, W.P.; Rheinstadter, M.C. Stabilization of Lipid Membranes through Partitioning of the Blood Bag Plasticizer Di-2-ethylhexyl phthalate (DEHP). Langmuir 2020, 36, 11899–11907. [Google Scholar] [CrossRef]
- Guo, J.; Ho, J.C.S.; Chin, H.; Mark, A.E.; Zhou, C.; Kjelleberg, S.; Liedberg, B.; Parikh, A.N.; Cho, N.J.; Hinks, J.; et al. Response of microbial membranes to butanol: Interdigitation vs. disorder. Phys. Chem. Chem. Phys. 2019, 21, 11903–11915. [Google Scholar] [CrossRef]
- Sharma, P.; Parthasarathi, S.; Patil, N.; Waskar, M.; Raut, J.S.; Puranik, M.; Ayappa, K.G.; Basu, J.K. Assessing Barriers for Antimicrobial Penetration in Complex Asymmetric Bacterial Membranes: A Case Study with Thymol. Langmuir 2020, 36, 8800–8814. [Google Scholar] [CrossRef]
- Faulkner, C.; Santos-Carballal, D.; Plant, D.F.; de Leeuw, N.H. Atomistic Molecular Dynamics Simulations of Propofol and Fentanyl in Phosphatidylcholine Lipid Bilayers. ACS Omega 2020, 5, 14340–14353. [Google Scholar] [CrossRef]
- Dickson, C.J.; Hornak, V.; Bednarczyk, D.; Duca, J.S. Using Membrane Partitioning Simulations to Predict Permeability of Forty-Nine Drug-Like Molecules. J. Chem. Inf. Model. 2019, 59, 236–244. [Google Scholar] [CrossRef]
- Gupta, R.; Sridhar, D.B.; Rai, B. Molecular Dynamics Simulation Study of Permeation of Molecules through Skin Lipid Bilayer. J. Phys. Chem. B 2016, 120, 8987–8996. [Google Scholar] [CrossRef]
- Gupta, R.; Rai, B. Molecular dynamics simulation study of translocation of fullerene C60 through skin bilayer: Effect of concentration on barrier properties. Nanoscale 2017, 9, 4114–4127. [Google Scholar] [CrossRef]
- Nunes, R.S.; Vila-Vicosa, D.; Costa, P.J. Halogen Bonding: An Underestimated Player in Membrane-Ligand Interactions. J. Am. Chem. Soc. 2021, 143, 4253–4267. [Google Scholar] [CrossRef]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Davila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M.; et al. CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; He, X.; Vanommeslaeghe, K.; MacKerell, A.D., Jr. Extension of the CHARMM General Force Field to sulfonyl-containing compounds and its utility in biomolecular simulations. J. Comput. Chem. 2012, 33, 2451–2468. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Tribello, G.A.; Bonomi, M.; Branduardi, D.; Camilloni, C.; Bussi, G. PLUMED 2: New feathers for an old bird. Comput. Phys. Commun. 2014, 185, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Hub, J.S.; de Groot, B.L.; van der Spoel, D. g_wham-A Free Weighted Histogram Analysis Implementation Including Robust Error and Autocorrelation Estimates. J. Chem. Theory Comput. 2010, 6, 3713–3720. [Google Scholar] [CrossRef] [Green Version]
- Van der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.L.; Engstrom, O.; Jo, S.; Stuhlsatz, D.; Yeom, M.S.; Klauda, J.B.; Widmalm, G.; Im, W. Molecular dynamics and NMR spectroscopy studies of E. coli lipopolysaccharide structure and dynamics. Biophys. J. 2013, 105, 1444–1455. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zheng, J.; Yang, Q.; Dang, Z.; Zhang, L. Effect of carbon chain structure on the phthalic acid esters (PAEs) adsorption mechanism by mesoporous cellulose biochar. Chem. Eng. J. 2019, 362, 383–391. [Google Scholar] [CrossRef]
- Sanchez-Iglesias, A.; Grzelczak, M.; Altantzis, T.; Goris, B.; Perez-Juste, J.; Bals, S.; Van Tendeloo, G.; Donaldson, S.H., Jr.; Chmelka, B.F.; Israelachvili, J.N.; et al. Hydrophobic interactions modulate self-assembly of nanoparticles. ACS Nano 2012, 6, 11059–11065. [Google Scholar] [CrossRef] [PubMed]
- Ersan, G.; Apul, O.G.; Perreault, F.; Karanfil, T. Adsorption of organic contaminants by graphene nanosheets: A review. Water Res. 2017, 126, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Cousins, L.; Mackay, D. Correlating the physical ± chemical properties of phthalate esters using the ‘three solubility’ approach. Chemosphere 2000, 41, 1389–1399. [Google Scholar] [CrossRef]
- Zuo, X.; Yi, P.; Chen, Q.; Wu, M.; Zhang, L.; Pan, B.; Xing, B. Inter-molecular interactions of phthalic acid esters and multi-stage sorption revealed by experimental investigations and computation simulations. Chem. Eng. J. 2022, 431, 134018. [Google Scholar] [CrossRef]
- Horowitzd, B.; StrykeP, M.H.; Waldmana, A.A.; Woods, K.R.; Gassh, J.D.; Drago, J. Stabilization of red blood cells by the plasticizer, Diethylhexylphthalate. Vox Sang. 1985, 48, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Farrag, H.A.; Abdallah, N.; Shehata, M.M.K.; Awad, E.M. Natural outer membrane permeabilizers boost antibiotic action against irradiated resistant bacteria. J. Biomed. Sci. 2019, 26, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, M.; Payasi, A. Changing Trends of Commonly Used Intensive Care Unit Antibiotics Due to Differential Membrane Permeability in Resistant Escherichia coli Collected in EASE Programme. J. Microb. Biochem. Technol. 2013, 5, 84–87. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation Type | Systems | NPAEs | Simulation Time |
|---|---|---|---|
| cMD simulations | Pure POPC | 0 | 200 ns × 3 |
| POPC + DMP | 8 | 500 ns × 3 | |
| POPC + DBP | 8 | ||
| POPC + DEHP | 8 | ||
| SMD simulations and umbrella sampling | POPC + DMP | 1 | 50 ns/window |
| POPC + DBP | 1 | ||
| POPC + DEHP | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Y.; Li, M.; Xie, Y.; Guo, J. Investigating the Permeation Mechanism of Typical Phthalic Acid Esters (PAEs) and Membrane Response Using Molecular Dynamics Simulations. Membranes 2022, 12, 596. https://doi.org/10.3390/membranes12060596
Bao Y, Li M, Xie Y, Guo J. Investigating the Permeation Mechanism of Typical Phthalic Acid Esters (PAEs) and Membrane Response Using Molecular Dynamics Simulations. Membranes. 2022; 12(6):596. https://doi.org/10.3390/membranes12060596
Chicago/Turabian StyleBao, Yiqiong, Mengrong Li, Yanjie Xie, and Jingjing Guo. 2022. "Investigating the Permeation Mechanism of Typical Phthalic Acid Esters (PAEs) and Membrane Response Using Molecular Dynamics Simulations" Membranes 12, no. 6: 596. https://doi.org/10.3390/membranes12060596