
Acid-Sensing Ion Channel 2: Function and Modulation


Abstract
:1. Introduction
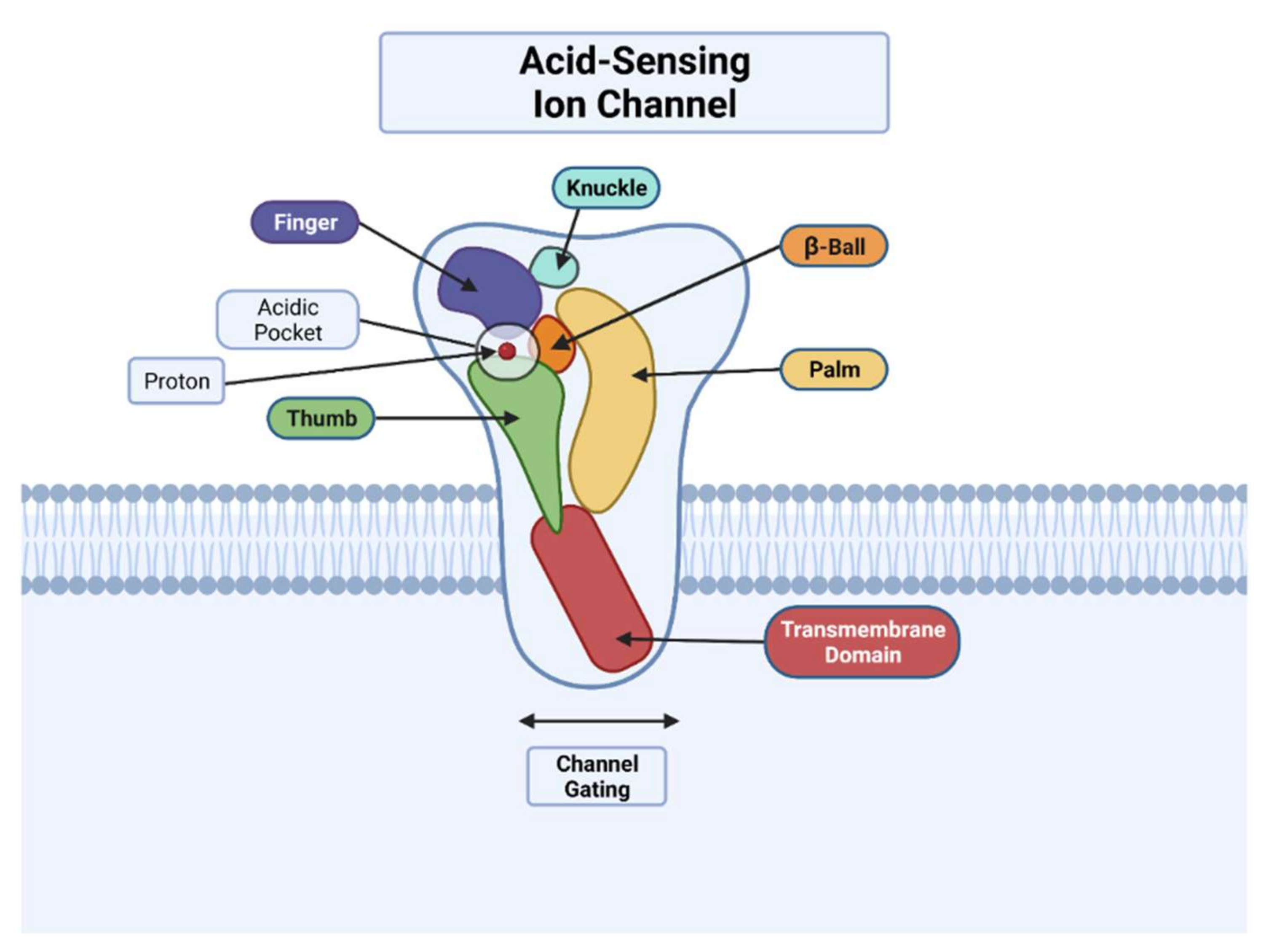
2. Function
2.1. Physiological Function
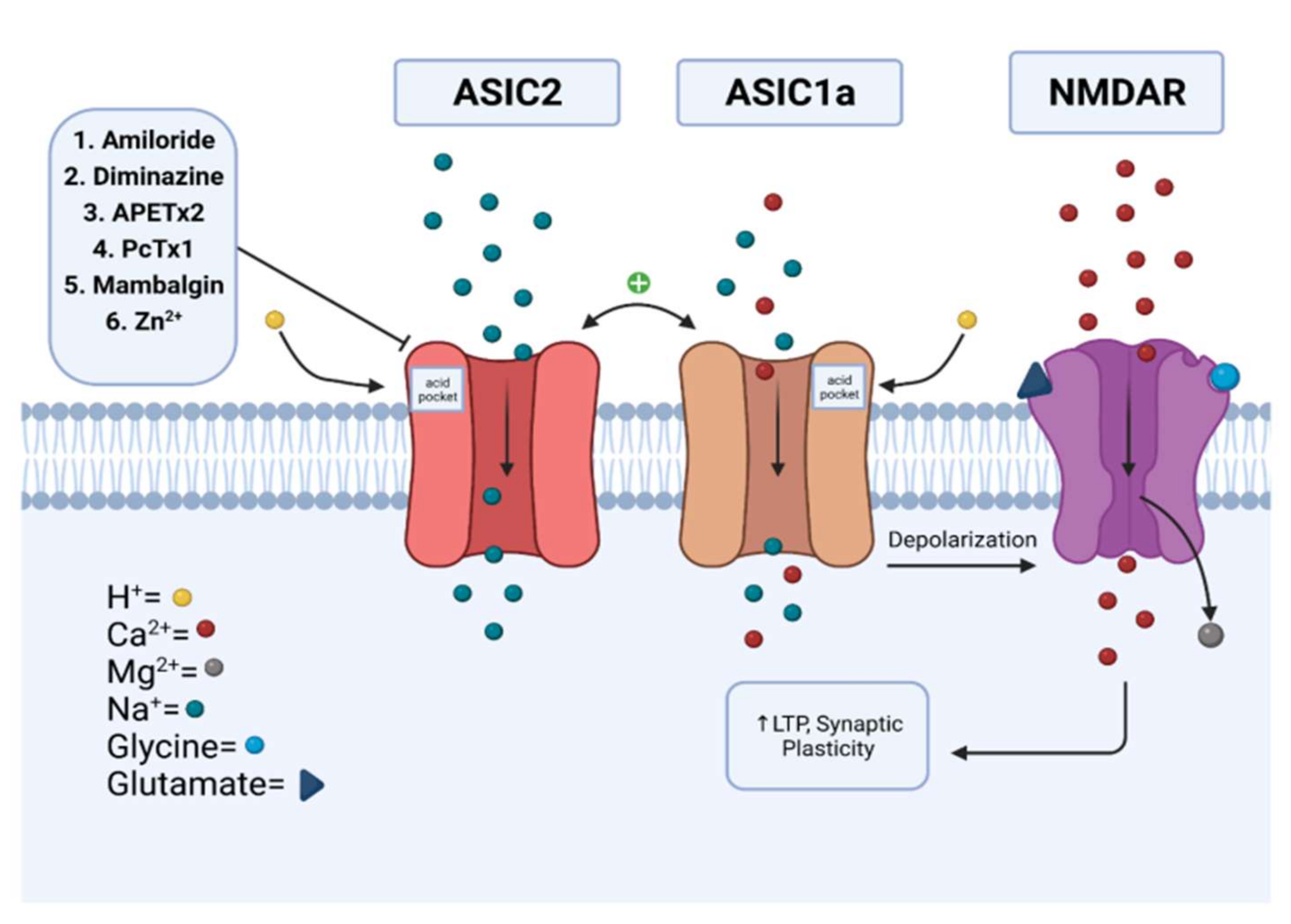
2.1.1. Synaptic Role of ASIC2
2.1.2. Mechanosensation of ASIC2
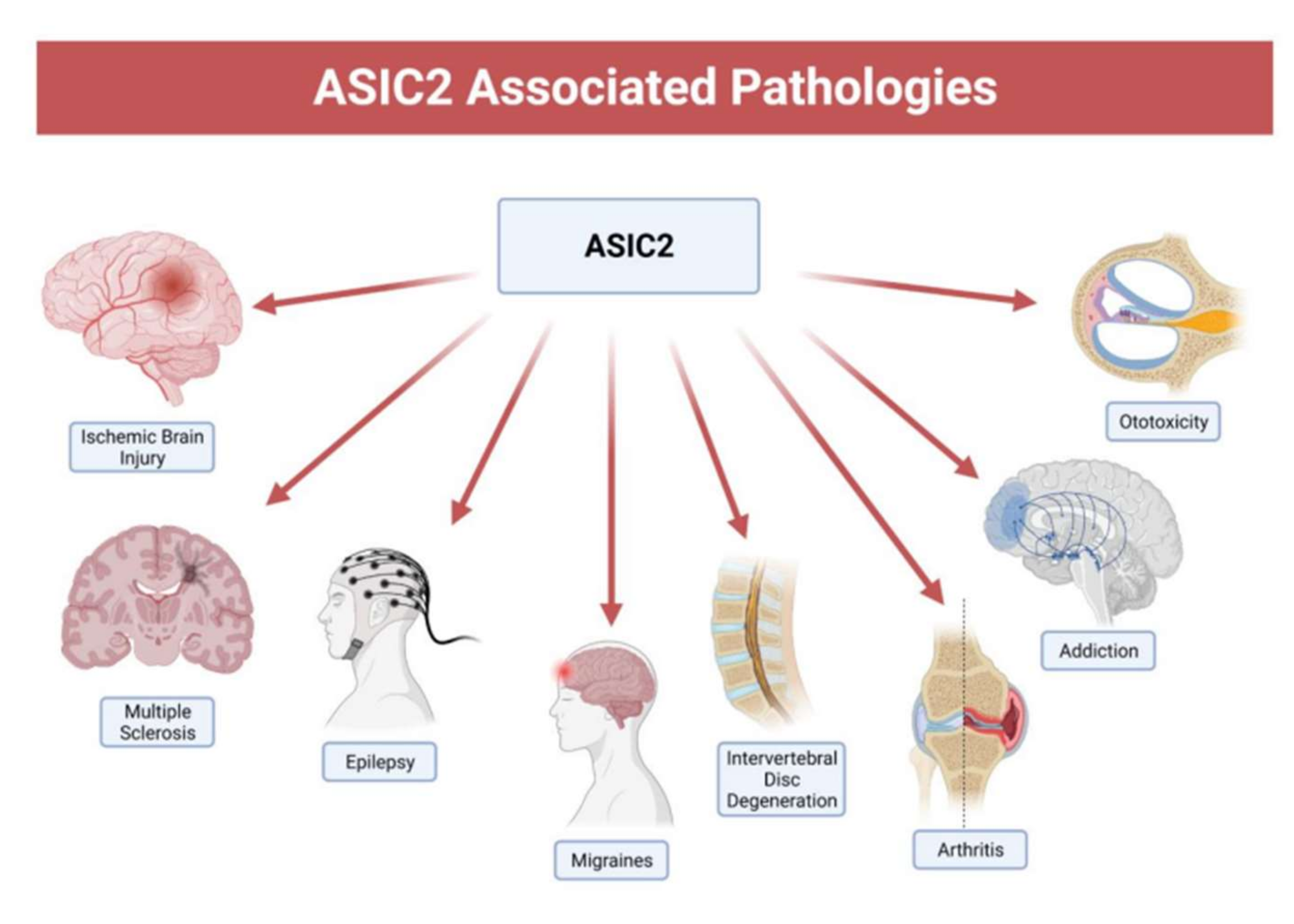
2.2. Pathology of ASIC2
2.2.1. Ischemic Brain Injury
2.2.2. Multiple Sclerosis
2.2.3. Epilepsy
2.2.4. Migraines
2.2.5. Intervertebral Disc Degeneration
2.2.6. Arthritis
2.2.7. Addiction
2.2.8. Aminoglycoside-Induced Hearing Loss
3. Modulation
3.1. Amiloride
3.2. Diminazene
3.3. APETx2
3.4. Psalmotoxin (PcTx1)
3.5. Mambalgins
3.6. Zinc
4. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AMPAR | α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor |
| ASICs | acid-sensing ion channels |
| ASOR | acid-sensitive outward rectifying anion channel |
| CNS | central nervous system |
| DEG/ENaC | degenerin/epithelial sodium channel |
| DRG | dorsal root ganglion |
| EAE | experimental autoimmune encephalitis |
| GPCR | G-protein-coupled receptor |
| GPR | G-protein-coupled proton-sensing receptor |
| HD | Huntingdon’s disease |
| IVDD | intervertebral disc degeneration |
| KO | Knock-out |
| LTD | long-term depression |
| LTP | long-term potentiation |
| MCAO | middle cerebral artery occlusion |
| mPFC | medial prefrontal cortex |
| MS | multiple sclerosis |
| NAc | nucleus accumbens |
| NMDAR | N-methyl-D-aspartate receptor |
| PAC | proton-activated chloride channel |
| PcTx1 | psalmotoxin-1 |
| PD | Parkinson’s disease |
| PDB | protein data bank |
| RTN | retrotrapezoid nucleus |
| SGNs | spiral ganglion neurons |
| TM | transmembrane domain |
| WT | wild-type |
References
- Groc, L.; Choquet, D. Linking glutamate receptor movements and synapse function. Science 2020, 368, eaay4631. [Google Scholar] [CrossRef]
- Pagonabarraga, J.; Tinazzi, M.; Caccia, C.; Jost, W.H. The role of glutamatergic neurotransmission in the motor and non-motor symptoms in Parkinsons disease: Clinical cases and a review of the literature. J. Clin. Neurosci. 2021, 90, 178–183. [Google Scholar] [CrossRef]
- Gruol, D.L.; Barker, J.L.; Li-Yen, H.M.; MacDonald, J.F.; Smith, T.G., Jr. Hydrogen ions have multiple effects on the excitability of cultured mammalian neurons. Brain Res. 1980, 183, 247–252. [Google Scholar] [CrossRef]
- Krishtal, O.A.; Pidoplichko, V.I. A receptor for protons in the nerve cell membrane. Neuroscience 1980, 5, 2325–2327. [Google Scholar] [CrossRef]
- Krishtal, O.A.; Pidoplichko, V.I. A receptor for protons in the membrane of sensory neurons may participate in nociception. Neuroscience 1981, 6, 2599–2601. [Google Scholar] [CrossRef]
- Cheng, Y.; Jiang, B.; Chen, C. Acid-sensing ion channels: Dual function proteins for chemo-sensing and mechano-sensing. J. Biomed. Sci. 2018, 25, 46. [Google Scholar] [CrossRef]
- Waldmann, R.; Champigny, G.; Bassilana, F.; Heurteaux, C.; Lazdunski, M. A proton-gated cation channel involved in acid-sensing. Nature 1997, 386, 173–177. [Google Scholar] [CrossRef]
- Krishtal, O. The ASICs: Signaling molecules? Modulators? Trends Neurosci. 2003, 26, 477–483. [Google Scholar] [CrossRef]
- Bartoi, T.; Augustinowski, K.; Polleichtner, G.; Gründer, S.; Ulbrich, M.H. Acid-sensing ion channel (ASIC) 1a/2a heteromers have a flexible 2:1/1:2 stoichiometry. Proc. Natl. Acad. Sci. USA 2014, 111, 8281–8286. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Xu, Y.; Jiang, Y.-Q.; Xu, J.; Hu, Y.; Zha, X. ASIC subunit ratio and differential surface trafficking in the brain. Mol. Brain. 2016, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Kellenberger, S.; Schild, L. International Union of Basic and Clinical Pharmacology. XCI. Structure, function, and pharmacology of acid-sensing ion channels and the epithelial Na+ channel. Pharmacol. Rev. 2015, 67, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gründer, S.; Pusch, M. Biophysical properties of acid-sensing ion channels (ASICs). Neuropharmacology 2015, 94, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, T.W.; Frey, E.N.; Askwith, C.C. Structure and activity of the acid-sensing ion channels. Am. J. Physiol. Cell Physiol. 2012, 303, 699–710. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, E.B.; Kawate, T.; Gouaux, E. Pore architecture and ion sites in acid-sensing ion channels and P2X receptors. Nature 2009, 460, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Ruan, N.; Tribble, J.; Peterson, A.M.; Jiang, Q.; Wang, J.Q.; Chu, X.P. Acid-sensing ion channels and mechanosensation. Int. J. Mol. Sci. 2021, 22, 4810. [Google Scholar] [CrossRef] [PubMed]
- Jasti, J.; Furukawa, H.; Gonzales, E.B.; Gouaux, E. Structure of acid-sensing ion channel 1 at 1.9 A resolution and low pH. Nature 2007, 449, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wu, J.; Leng, T.; Yang, T.; Zhou, Y.; Jiang, Q.; Wang, B.; Hu, Y.; Ji, Y.H.; Simon, R.P.; et al. Region specific contribution of ASIC2 to acidosis-and ischemia-induced neuronal injury. J. Cereb. Blood Flow Metab. 2017, 37, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gao, G.; Zhang, Y.; Sun, Y.; Li, H.; Dong, S.; Ma, W.; Liu, B.; Wang, W.; Wu, H.; et al. Glucose deficiency elevates acid-sensing ion channel 2a expression and increases seizure susceptibility in temporal lobe epilepsy. Sci. Rep. 2017, 7, 5870. [Google Scholar] [CrossRef] [Green Version]
- Karsan, N.; Gonzales, E.B.; Dussor, G. Targeted acid-sensing ion channel therapies for migraine. Neurotherapeutics 2018, 15, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, A.; del Valle, M.E.; Garcia-Suarez, O.; Vina, E.; Cabo, R.; Vazquez, G.; Cobo, J.L.; Murcia, A.; Alvarez-Vega, M.; García-Cosamalón, J.; et al. Acid-sensing ion channels in healthy and degenerated human intervertebral disc. Connect. Tissue Res. 2014, 55, 197–204. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, X.; Wang, Z.; Ge, J.; Chen, F. Interleukin-6 enhances acid-induced apoptosis via upregulating acid-sensing ion channel 1a expression and function in rat articular chondrocytes. Int. Immunopharmacol. 2015, 29, 748–760. [Google Scholar] [CrossRef] [Green Version]
- Kreple, C.J.; Lu, Y.; Taugher, R.J.; Schwager-Gutman, A.L.; Du, J.; Stump, M.; Wemmie, J.A.; Wang, Y.; Ghobbeh, A.; Fan, R.; et al. Acid-sensing ion channels contribute to synaptic transmission and inhibit cocaine-evoked plasticity. Nat. Neurosci. 2014, 17, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Suman, A.; Mehta, B.; Guo, M.; Chu, X.; Fibuch, E.E.; Mao, L.; Wang, J.Q. Alterations in subcellular expression of acid-sensing ion channels in the rat forebrain following chronic amphetamine administration. Neurosci. Res. 2010, 68, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Wang, F.; Zhang, C.; Mao, C.J.; Yang, J.; Yang, Y.P.; Liu, S.; Hu, L.F.; Liu, C.F. Neuroprotective effects of paeoniflorin on 6-OHDA-lesioned rat model of Parkinson’s disease. Neurochem. Res. 2016, 41, 2923–2936. [Google Scholar] [CrossRef]
- Mango, D.; Nisticò, R. Role of ASIC1a in Aβ-induced synaptic alterations in the hippocampus. Pharmacol. Res. 2018, 131, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Mango, D.; Nisticò, R. Neurodegenerative disease: What potential therapeutic role of acid-sensing ion channels? Front. Cell. Neurosci. 2021, 15, 730641. [Google Scholar] [CrossRef] [PubMed]
- Staniland, A.A.; McMahon, S.B. Mice lacking acid-sensing ion channels (ASIC) 1 or 2, but not ASIC3, show increased pain behaviour in the formalin test. Eur. J. Pain 2009, 13, 554–563. [Google Scholar] [CrossRef]
- Storozhuka, M.; Cherninskyia, A.; Maximyuka, O.; Isaeva, D.; Krishtala, O. Acid-sensing ion channels: Focus on physiological and some pathological roles in the brain. Curr. Neuropharmacol. 2021, 19, 1570–1589. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, T.W.; Lee, K.G.; Gormley, M.G.; Askwith, C.C. Heteromeric acid-sensing ion channels (ASICs) composed of ASIC2b and ASIC1a display novel channel properties and contribute to acidosis-induced neuronal death. J. Neurosci. 2011, 31, 9723–9734. [Google Scholar] [CrossRef] [Green Version]
- Deval, E.; Salinas, M.; Baron, A.; Lingueglia, E.; Lazdunski, M. ASIC2b-dependent regulation of ASIC3, an essential acid-sensing ion channel subunit in sensory neurons via the partner protein PICK-1. J. Biol. Chem. 2004, 279, 19531–19539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunay, A.; Gasull, X.; Salinas, M.; Noel, J.; Friend, V.; Lingueglia, E.; Deval, E. Human ASIC3 channel dynamically adapts its activity to sense the extracellular pH in both acidic and alkaline directions. Proc. Natl. Acad. Sci. USA 2012, 109, 13124–13129. [Google Scholar] [CrossRef] [Green Version]
- Vralsted, V.C.; Price, M.P.; Du, J.; Schnizler, M.; Wunsch, A.M.; Ziemann, A.E.; Welsh, M.J.; Wemmie, J.A. Expressing acid-sensing ion channel 3 in the brain alters acid-evoked currents and impairs fear conditioning. Genes Brain Behav. 2011, 10, 444–450. [Google Scholar] [CrossRef]
- Akopian, A.N.; Chen, C.C.; Ding, Y.; Cesare, P.; Wood, J.N. A new member of the acid-sensing ion channel family. Neuroreport 2000, 11, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Grunder, S.; Geissler, H.S.; Bassler, E.L.; Ruppersberg, J.P. A new member of acid sensing ion channels from pituitary gland. Neuroreport 2000, 11, 1607–1611. [Google Scholar] [CrossRef]
- Lin, S.H.; Chien, Y.C.; Chiang, W.W.; Liu, Y.Z.; Lien, C.C.; Chen, C.C. Genetic mapping of ASIC4 and contrasting phenotype to ASIC1a in modulating innate fear and anxiety. Eur. J. Neurosci. 2015, 41, 1553–1568. [Google Scholar] [CrossRef]
- Gautschi, I.; van Bemmelen, M.X.; Schild, L. Proton and non-proton activation of ASIC channels. PLoS ONE 2017, 12, e0175293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, A.M.; Kusama, N.; Hattori, T.; Gautam, M.; Benson, C.J. ASIC2 subunits facilitate expression at the cell surface and confer regulation by PSD-95. PLoS ONE 2014, 9, e93797. [Google Scholar] [CrossRef]
- Miao, Y.; Zhang, W.; Lin, Y.; Lu, X.; Qiu, Y. Neuroprotective effects of ischemic preconditioning on global brain ischemia through up-regulation of acid-sensing ion channel 2a. Int. J. Mol. Sci. 2010, 11, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Chen, J.; Harding, A.M.; Price, M.P.; Lu, Y.; Abboud, F.M.; Benson, C.J. ASIC2a and ASIC3 heteromultimerize to form pH-sensitive channels in mouse cardiac dorsal root ganglia neurons. Circ. Res. 2009, 105, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immke, D.C.; McCleskey, E.W. Lactate enhances the acid-sensing Na+ channel on ischemia-sensing neurons. Nat. Neurosci. 2001, 4, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jiang, N.; Li, J.; Ji, Y.H.; Xiong, Z.G.; Zha, X.M. Two aspects of ASIC function: Synaptic plasticity and neuronal injury. Neuropharmacology 2015, 94, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchitel, O.D.; González-Inchauspe, C.; Weissmann, C. Synaptic signals mediated by protons and acid-sensing ion channels. Synapse 2019, 73, e22120. [Google Scholar] [CrossRef] [PubMed]
- González-Inchauspe, C.; Gobetto, M.N.; Uchitel, O.D. Modulation of acid sensing ion channel dependent protonergic neurotransmission at the mouse calyx of Held. Neuroscience 2020, 439, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Gobetto, M.N.; González-Inchauspe, C.; Uchitel, O.D. Histamine and corticosterone modulate acid sensing ion channels (ASICs) dependent long-term potentiation at the mouse anterior cingulate cortex. Neuroscience 2021, 460, 145–160. [Google Scholar] [CrossRef]
- Li, H.S.; Su, X.Y.; Song, X.L.; Qi, X.; Li, Y.; Wang, R.Q.; Maximyuk, O.; Krishtal, O.; Wang, T.; Fang, H.; et al. Protein kinase c lambda mediates acid-sensing ion channel 1a-dependent cortical synaptic plasticity and pain hypersensitivity. J. Neurosci. 2019, 39, 5773–5793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mango, D.; Nisticò, R. Acid-sensing ion channel 1a is involved in N-methyl D-aspartate receptor-dependent long-term depression in the hippocampus. Front. Pharmacol. 2019, 10, 555. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Reznikov, L.R.; Price, M.P.; Zha, X.M.; Lu, Y.; Moninger, T.O.; Wemmie, J.A.; Welsh, M.J. Protons are a neurotransmitter that regulates synaptic plasticity in the lateral amygdala. Proc. Natl. Acad. Sci. USA 2014, 111, 8961–8966. [Google Scholar] [CrossRef] [Green Version]
- Zha, X.M.; Costa, V.; Harding, A.M.; Reznikov, L.; Benson, C.J.; Welsh, M.J. ASIC2 subunits target acid-sensing ion channels to the synapse via an association with PSD-95. J. Neurosci. 2009, 29, 8438–8446. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Bredt, D.S.; Nicoll, R.A. AMPA receptor trafficking at excitatory synapses. Neuron 2003, 40, 361–379. [Google Scholar] [CrossRef] [Green Version]
- Price, M.P.; Gong, H.; Parsons, M.G.; Kundert, J.R.; Reznikov, L.R.; Bernardinelli, L.; Chaloner, K.; Buchanan, G.F.; Wemmie, J.A.; Richerson, G.B.; et al. Localization and behaviors in null mice suggest that ASIC1 and ASIC2 modulate responses to aversive stimuli. Genes Brain Behav. 2014, 13, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taugher, R.J.; Lu, Y.; Fan, R.; Ghobbeh, A.; Kreple, C.J.; Faraci, F.M.; Wemmie, J.A. ASIC1A in neurons is critical for fear-related behaviors. Genes Brain Behav. 2017, 16, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.J.; Price, M.P.; Xie, J. Biochemical basis of touch perception: Mechanosensory functions of degenerin/epithelial Na+ channels. J. Biol. Chem. 2002, 277, 2369–2372. [Google Scholar] [CrossRef] [Green Version]
- Price, M.; Lewin, G.; McIlwrath, S.; Cheung, C.; Xie, J.; Heppenstall, P.A.; Stucky, C.L.; Mannsfeldt, A.G.; Brennan, T.J.; Drummond, H.A. The mammalian sodium channel BNC1 is required for normal touch sensation. Nature 2000, 407, 1007–1011. [Google Scholar] [CrossRef]
- Cabo, R.; Alonso, P.; Viña, E.; Vázquez, G.; Gago, A.; Feito, J.; Pérez-Moltó, F.J.; García-Suárez, O.; Vega, J.A. ASIC2 is present in human mechanosensory neurons of the dorsal root ganglia and in mechanoreceptors of the glabrous skin. Histochem. Cell Biol. 2015, 143, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Wong, C.W. Neurosensory mechanotransduction through acid-sensing ion channels. J. Cell. Mol. Med. 2013, 17, 337–349. [Google Scholar] [CrossRef]
- Calavia, M.G.; Montaño, J.A.; García-Suárez, O.; Feito, J.; Guervós, M.A.; Germanà, A.; del Valle, M.; Pérez-Piñera, P.; Cobo, J.; Vega, J.A. Differential localization of acid-sensing ion channels 1 and 2 in human cutaneous pacinian corpuscles. Cell. Mol. Neurobiol. 2010, 30, 841–848. [Google Scholar] [CrossRef]
- Simon, A.; Shenton, F.; Hunter, I.; Banks, R.W.; Bewick, G.S. Amiloride-sensitive channels are a major contributor to mechanotransduction in mammalian muscle spindles. J. Physiol. 2010, 588, 171–185. [Google Scholar] [CrossRef]
- Barth, D.; Fronius, M. Shear force modulates the activity of acid-sensing ion channels at low pH or in the presence of non-proton ligands. Sci. Rep. 2019, 9, 6781. [Google Scholar] [CrossRef]
- Peng, B.G.; Ahmad, S.; Chen, S.; Chen, P.; Price, M.P.; Lin, X. Acid-sensing ion channel 2 contributes a major component to acid-evoked excitatory responses in spiral ganglion neurons and plays a role in noise susceptibility of Mice. J. Neurosci. 2004, 24, 10167–10175. [Google Scholar] [CrossRef]
- Roza, C.; Puel, J.L.; Kress, M.; Baron, A.; Diochot, S.; Lazdunski, M.; Waldmann, R. Knockout of the ASIC2 channel in mice does not impair cutaneous mechanosensation, visceral mechanonociception and hearing. J. Physiol. 2004, 558, 659–669. [Google Scholar] [CrossRef] [PubMed]
- González-Garrido, A.; Vega, R.; Mercado, F.; López, I.A.; Soto, E. Acid-sensing ion channels expression, identity and role in the excitability of the cochlear afferent neurons. Front. Cell. Neurosci. 2015, 9, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Ma, X.; Sabharwal, R.; Snitsarev, V.; Morgan, D.; Rahmouni, K.; Drummond, H.A.; Whiteis, C.A.; Costa, V.; Price, M.; et al. The ion channel ASIC2 is required for baroreceptor and autonomic control of the circulation. Neuron 2009, 24, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Gannon, K.; McKey, S.; Stec, D.; Drummond, H. Altered myogenic vasoconstriction and regulation of whole kidney blood flow in the asic2 knockout mouse. Am. J. Physiol. Renal Physiol. 2015, 308, F339–F348. [Google Scholar] [CrossRef] [Green Version]
- Laude, D.; Baudrie, V.; Elghozi, J.L. Applicability of recent methods used to estimate spontaneous baroreflex sensitivity to resting mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R142–R150. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.P.; Grasing, K.A.; Wang, J.Q. Acid-sensing ion channels contribute to neurotoxicity. Transl. Stroke Res. 2014, 5, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siesjo, B.K.; Katsura, K.; Mellergard, P.; Ekholm, A.; Lundgren, J.; Smith, M.L. Acidosis related brain damage. Prog. Brain Res. 1993, 96, 23–48. [Google Scholar]
- Xiong, Z.G.; Zhu, X.M.; Chu, X.P.; Minami, M.; Hey, J.; Wen, W.L.; MacDonald, J.; Wemmie, J.A.; Price, M.P.; Welsh, M.J.; et al. Neuroprotection in ischemia: Blocking calcium-permeable acid-sensing ion channels. Cell 2004, 118, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Pignataro, G.; Simon, R.P.; Xiong, Z.G. Prolonged activation of ASIC1a and the time window for neuroprotection in cerebral ischaemia. Brain 2007, 130, 151–158. [Google Scholar] [CrossRef]
- Gao, J.; Duan, B.; Wang, D.G.; Deng, X.H.; Zhang, G.Y.; Xu, L.; Xu, T.L. Coupling between NMDA receptor and acid-sensing ion channel contributes to ischemic neuronal death. Neuron 2005, 48, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.B.; Jin, K.; Minami, M.; Chen, D.; Simon, R.P. Global ischemia induces expression of acid-sensing ion channel 2a in rat brain. J. Cereb. Blood Flow Metab. 2001, 21, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.M.; Wallin, M.T.; Marrie, R.A.; Culpepper, W.J.; Langer-Gould, A.; Campbell, J.D.; Buka, S.; Tremlett, H.; Cutter, G.; Kaye, W. The prevalence of MS in the United States. Neurology 2019, 92, 469–480. [Google Scholar] [CrossRef]
- Friese, M.A.; Craner, M.J.; Etzensperger, R.; Vergo, S.; Wemmie, J.A.; Welsh, M.J. Acid-sensing ion channel-1 contributes to axonal degeneration in autoimmune inflammation of the central nervous system. Nat. Med. 2007, 13, 1483–1489. [Google Scholar] [CrossRef]
- Vergo, S.; Craner, M.J.; Etzensperger, R.; Attfield, K.; Friese, M.A.; Newcombe, J.; Esiri, M.; Fugger, L. Acid-sensing ion channel 1 is involved in both axonal injury and demyelination in multiple sclerosis and its animal model. Brain 2011, 134, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.P.; Xiong, Z.G. Physiological and pathological functions of acid-sensing ion channels in the central nervous system. Curr. Drug Targets 2012, 13, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Wu, X.; Wang, Z.; Xie, Y.; Ge, J.; Chen, F. Novel insights into acid-sensing ion channels: Implications for degenerative diseases. Aging Dis. 2016, 7, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardinelli, L.; Murgia, S.B.; Bitti, P.P.; Focco, L.; Musu, L.; Prokopenko, I.; Pastorino, R.; Saddi, V.; Ticca, A.; Piras, M.L. Association between the ACCN1 gene and multiple sclerosis in Central East Sardinia. PLoS ONE 2007, 2, e480. [Google Scholar] [CrossRef] [PubMed]
- Fazia, T.; Pastorino, R.; Notartomaso, S.; Busceti, C.; Imbriglio, T.; Cannella, M.; Gentilini, D.; Morani, G.; Ticca, A.; Bitti, P.; et al. Acid sensing ion channel 2: A new potential player in the pathophysiology of multiple sclerosis. Eur. J. Neurosci. 2019, 49, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, A.E.; Schnizler, M.K.; Albert, G.W.; Severson, M.A.; Howard, M.A., 3rd; Welsh, M.J.; Wemmie, J.A. Seizure termination by acidosis depends on ASIC1a. Nat Neurosci. 2008, 11, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.; Xiao, Z.M.; Wang, X.; Weng, C.; Ding, M.; Zhu, F.; Lu, Z.N. Inhibition of acid sensing ion channel 3 aggravates seizures by regulating NMDAR function. Neurochem. Res. 2018, 43, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Jones-Muhammad, M.; Shao, Q.; Cain-Shields, L.; Shaffery, J.P.; Warrington, J.P. Acid sensing ion channel 2a is reduced in the reduced uterine perfusion pressure mouse model and increases seizure susceptibility in pregnant mice. Cells 2021, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- N’Gouemo, P. Amiloride delays the onset of pilocarpine-induced seizures in rats. Brain Res. 2008, 1222, 230–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levglevskyi, O.; Isaev, D.; Netsyk, O.; Romanov, A.; Fedoriuk, M.; Maximyuk, O.; Isaeva, E.; Akaike, N.; Krishtal, O. Acid-sensing ion channels regulate spontaneous inhibitory activity in the hippocampus: Possible implications for epilepsy. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, R.J.; He, J.S.; Fu, Y.H.; Zhang, Y.Q.; Shao, X.Q.; Wu, L.W.; Lu, Q.; Jin, L.R.; Liu, H. ASIC1a polymorphism is associated with temporal lobe epilepsy. Epilepsy Res. 2011, 96, 74–80. [Google Scholar] [CrossRef]
- Wu, H.; Wang, C.; Liu, B.; Li, H.; Zhang, Y.; Dong, S.; Gao, G.; Zhang, H. Altered expression pattern of acid-sensing ion channel isoforms in piriform cortex after seizures. Mol. Neurobiol. 2016, 53, 1782–1793. [Google Scholar] [CrossRef]
- Wang, S.; Wu, B.X.; Liu, C.Y.; Qin, G.C.; Yan, W.H.; Zhou, J.Y.; Chen, L.X. Expression of ASIC3 in the trigeminal nucleus caudalis plays a role in a rat model of recurrent migraine. J. Mol. Neurosci. 2018, 66, 44–52. [Google Scholar] [CrossRef]
- Verkest, C.; Piquet, E.; Diochot, S.; Dauvois, M.; Lanteri-Minet, M.; Lingueglia, E.; Baron, A. Effects of systemic inhibitors of acid-sensing ion channels 1 (ASIC1) against acute and chronic mechanical allodynia in a rodent model of migraine. Br. J. Pharmacol. 2018, 175, 4154–4166. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Edelmayer, R.M.; Wei, X.; De Felice, M.; Porreca, F.; Dussor, G. Dural afferents express acid-sensing ion channels: A role for decreased meningeal pH in migraine headache. Pain 2011, 152, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Fontana, G.; See, E.; Pandit, A. Current trends in biologics delivery to restore intervertebral disc anabolism. Adv. Drug Deliv. Rev. 2015, 84, 146–158. [Google Scholar] [CrossRef]
- Sakai, D.; Grad, S. Advancing the cellular and molecular therapy for intervertebral disc disease. Adv. Drug Deliv. Rev. 2015, 84, 159–171. [Google Scholar] [CrossRef]
- Tetlow, L.C.; Adlam, D.J.; Woolley, D.E. Matrix metalloproteinase and proinflammatory cytokine production by chondrocytes of human osteoarthritic cartilage: Associations with degenerative changes. Arthritis Rheum. 2001, 44, 585–594. [Google Scholar] [CrossRef]
- Wuertz, K.; Godburn, K.; Iatridis, J.C. MSC response to pH levels found in degenerating intervertebral discs. Biochem. Biophys. Res. Commun. 2009, 379, 824–829. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, F.R.; Xu, R.S.; Hu, W.; Jiang, D.L.; Ji, C.; Chen, F.H.; Yuan, F.L. Acid-sensing ion channel 1a-mediated calcium influx regulates apoptosis of endplate chondrocytes in intervertebral discs. Expert Opin. Ther. Targets 2014, 18, 1–14. [Google Scholar] [CrossRef]
- Yuan, F.L.; Chen, F.H.; Lu, W.G.; Li, X.; Li, J.P.; Li, C.W.; Xu, R.S.; Wu, F.R.; Hu, W.; Zhang, T.Y. Inhibition of acid-sensing ion channels in articular chondrocytes by amiloride attenuates articular cartilage destruction in rats with adjuvant arthritis. Inflamm. Res. 2010, 59, 939–947. [Google Scholar] [CrossRef]
- Razaq, S.; Wilkins, R.J.; Urban, J.P. The effect of extracellular pH on matrix turnover by cells of the bovine nucleus pulposus. Eur. Spine J. 2003, 12, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Boyle, M. Neuroscience of addiction: Relevance to prevention and treatment. Am. J. Psychiatry 2018, 175, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F. Drug addiction: Hyperkatifeia/negative reinforcement as a framework for medications development. Pharmacol. Rev. 2021, 73, 163–201. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T.W. Drug addiction: Updating actions to habits to compulsions ten Years on. Annu. Rev. Psychol. 2016, 67, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Buck, S.A.; Torregrossa, M.M.; Logan, R.W.; Freyberg, Z. Roles of dopamine and glutamate co-release in the nucleus accumbens in mediating the actions of drugs of abuse. FEBS J. 2021, 288, 1462–1474. [Google Scholar] [CrossRef] [PubMed]
- Gutman, A.L.; Cosme, C.V.; Noterman, M.F.; Worth, W.R.; Wemmie, J.A.; Lalumiere, R.T. Overexpression of ASIC1A in the nucleus accumbens of rats potentiates cocaine-seeking behavior. Addict. Biol. 2018, 25, e12690. [Google Scholar] [CrossRef] [PubMed]
- Kreple, C.J.; Lu, Y.; LaLumiere, R.T.; Wemmie, J.A. Drug abuse and the simplest neurotransmitter. ACS Chem. Neurosci. 2014, 5, 746–748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.C.; Mao, L.M.; Wang, J.Q.; Chu, X.P. Upregulation of acid-sensing ion channel 1 protein expression by chronic administration of cocaine in the mouse striatum in vivo. Neurosci. Lett. 2009, 459, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Wang, C.; Fibuch, E.; Wang, J.; Chu, X.P. Differential regulation of locomotor activity to acute and chronic cocaine administration by acid-sensing ion channel 1a and 2 in adult mice. Neuroscience 2013, 246, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Selimoglu, E. Aminoglycoside-induced ototoxicity. Curr. Pharm. Des. 2007, 13, 119–126. [Google Scholar] [CrossRef]
- Strupp, M.; Arbusow, V. Acute vestibulopathy. Curr. Opin. Neurol. 2001, 14, 11–20. [Google Scholar] [CrossRef]
- Garza, A.; López-Ramírez, O.; Vega, R.; Soto, E. The aminoglycosides modulate the acid-sensing ionic-channel (ASIC) currents in dorsal-root ganglion neurons from the rat. J. Pharmacol. Exp. Ther. 2010, 332, 489–499. [Google Scholar] [CrossRef]
- Diering, G.H.; Numata, M. Endosomal pH in neuronal signaling and synaptic transmission: Role of Na+/H+ exchanger NHE5. Front. Physiol. 2014, 4, 412. [Google Scholar] [CrossRef] [Green Version]
- Soto, E.; Ortega-Ramírez, A.; Vega, R. Protons as messengers of intercellular communication in the nervous system. Front. Cell. Neurosci. 2018, 12, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vullo, S.; Kellenberger, S. A molecular view of the function and pharmacology of acid-sensing ion channels. Pharmacol. Res. 2020, 154, 104166. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.P.; Saez, N.J.; Cristofori-Armstrong, B.; Anangi, R.; King, G.F.; Smith, M.T.; Rash, L.D. Inhibition of acid-sensing ion channels by diminazene and APETx2 evoke partial and highly variable anti-hyperalgesia in a rat model of inflammatory pain. Br. J. Pharmacol. 2018, 175, 2204–2218. [Google Scholar] [CrossRef]
- Schmidt, A.; Rossetti, G.; Joussen, S.; Gründer, S. Diminazene is a slow pore blocker of acid-sensing ion channel 1a (ASIC1a). Mol. Pharmacol. 2017, 92, 665–675. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef]
- Holton, C.M.; Strother, L.C.; Dripps, I.; Pradhan, A.A.; Goadsby, P.J.; Holland, P.R. Acid-sensing ion channel 3 blockade inhibits durovascular and nitric oxide-mediated trigeminal pain. Br. J. Pharmacol. 2020, 177, 2478–2486. [Google Scholar] [CrossRef]
- Yen, L.T.; Hsieh, C.L.; Hsu, H.C.; Lin, Y.W. Preventing the induction of acid saline-induced fibromyalgia pain in mice by electroacupuncture or APETx2 injection. Acupunct. Med. 2020, 38, 188–193. [Google Scholar] [CrossRef]
- Baron, A.; Lingueglia, E. Pharmacology of acid-sensing ion channels—Physiological and therapeutical perspectives. Neuropharmacology 2015, 94, 19–35. [Google Scholar] [CrossRef]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venom. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Joeres, N.; Augustinowski, K.; Neuhof, A.; Assmann, M.; Gründer, S. Functional and pharmacological characterization of two different ASIC1a/2a heteromers reveals their sensitivity to the spider toxin PcTx1. Sci. Rep. 2016, 6, 27647. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Gao, B.; Xiong, Q.J.; Wang, Y.C.; Huang, D.K.; Wu, W.N. Acid-sensing ion channels contribute to the effect of extracellular acidosis on proliferation and migration of A549 cells. Tumour Biol. 2017, 39, 1010428317705750. [Google Scholar] [CrossRef] [Green Version]
- Brzezicki, M.A.; Zakowicz, P.T. Mambalgins, the venom-origin peptides as a potentially novel group of analgesics: Mini review. CNS Neurol. Disord. Drug Targets 2018, 17, 87–97. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Salinas, M.; Kessler, P.; Douguet, D.; Sarraf, D.; Tonali, N.; Thai, R.; Servent, D.; Lingueglia, E. Mambalgin-1 pain-relieving peptide locks the hinge between α4 and α5 helices to inhibit rat acid-sensing ion channel 1a. Neuropharmacology 2021, 185, 108453. [Google Scholar] [CrossRef]
- Sun, D.; Liu, S.; Li, S.; Zhang, M.; Yang, F.; Wen, M.; Shi, P.; Wang, T.; Pan, M.; Chang, S.; et al. Structural insights into human acid-sensing ion channel 1a inhibition by snake toxin mambalgin1. eLife 2020, 9, e57096. [Google Scholar] [CrossRef]
- Diochot, S.; Alloui, A.; Rodrigues, P.; Dauvois, M.; Friend, V.; Aissouni, Y.; Eschalier, A.; Lingueglia, E.; Baron, A. Analgesic effects of mambalgin peptide inhibitors of acid-sensing ion channels in inflammatory and neuropathic pain. Pain 2016, 157, 552–559. [Google Scholar] [CrossRef]
- Salinas, M.; Besson, T.; Delettre, Q.; Diochot, S.; Boulakirba, S.; Douguet, D.; Lingueglia, E. Binding site and inhibitory mechanism of the mambalgin-2 pain-relieving peptide on acid-sensing ion channel 1a. J. Biol. Chem. 2014, 289, 13363–13373. [Google Scholar] [CrossRef] [Green Version]
- Baron, A.; Schaefer, L.; Lingueglia, E.; Champigny, G.; Lazdunski, M. Zn2+ and H+ are coactivators of acid-sensing ion channels. J. Biol. Chem. 2001, 276, 35361–35367. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.P.; Wemmie, J.A.; Wang, W.Z.; Zhu, X.M.; Saugstad, J.A.; Price, M.P.; Simon, R.P.; Xiong, Z.G. Subunit-dependent high-affinity zinc inhibition of acid-sensing ion channels. J. Neurosci. 2004, 24, 8678–8689. [Google Scholar] [CrossRef]
- Ortega-Ramírez, A.; Vega, R.; Soto, E. Acid-sensing ion channels as potential therapeutic targets in neurodegeneration and neuroinflammation. Mediat. Inflamm. 2017, 2017, 3728096. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Barajas, C.; Coronel, I.; Floran, B. Dopamine receptors and neurodegeneration. Aging. Dis. 2015, 6, 349–368. [Google Scholar]
- Arias, R.L.; Sung, M.L.; Vasylyev, D.; Zhang, M.Y.; Albinson, K.; Kubek, K.; Kagan, N.; Beyer, C.; Lin, Q.; Dwyer, J.M.; et al. Amiloride is neuroprotective in an MPTP model of Parkinson’s disease. Neurobiol. Dis. 2008, 31, 334–341. [Google Scholar] [CrossRef]
- Komnig, D.; Imgrund, S.; Reich, A.; Gründer, S.; Falkenburger, B.H. ASIC1a deficient mice show unaltered neurodegeneration in the subacute MPTP model of Parkinson disease. PLoS ONE 2016, 11, e0165235. [Google Scholar] [CrossRef]
- Joch, M.; Ase, A.R.; Chen, C.X.; MacDonald, P.A.; Kontogiannea, M.; Corera, A.T.; Brice, A.; Seguela, P.; Fon, E. Parkin-mediated monoubiquitination of the PDZ protein PICK1 regulates the activity of acid-sensing ion channels. Mol. Biol. Cell 2007, 18, 3105–3118. [Google Scholar] [CrossRef] [Green Version]
- Agostinho, L.A.; dos Santos, S.R.; Alvarenga, R.M.; Paiva, C.L. A systematic review of the intergenerational aspects and the diverse genetic profiles of Huntington’s disease. Genet. Mol. Res. 2013, 12, 1974–1981. [Google Scholar] [CrossRef]
- Tsang, T.M.; Woodman, B.; Mcloughlin, G.A.; Griffin, J.L.; Tabrizi, S.J.; Bates, G.P.; Holmes, E. Metabolic characterization of the R6/2 transgenic mouse model of Huntington’s disease by high-resolution MAS 1H NMR spectroscopy. J. Proteome Res. 2006, 5, 483–492. [Google Scholar] [CrossRef]
- Josefsen, K.; Nielsen, S.M.; Campos, A.; Seifert, T.; Hasholt, L.; Nielsen, J.E. Reduced gluconeogenesis and lactate clearance in Huntington’s disease. Neurobiol. Dis. 2010, 40, 656–662. [Google Scholar] [CrossRef]
- Wong, H.K.; Bauer, P.O.; Kurosawa, M.; Goswami, A.; Washizu, C.; Machida, Y.; Tosaki, A.; Yamada, M.; Knopfel, T.; Nakamura, T.; et al. Blocking acid sensing ion channel 1 alleviates Huntington’s disease pathology via an ubiquitin-proteasome system dependent mechanism. Hum. Mol. Genet. 2008, 17, 3223–3235. [Google Scholar] [CrossRef] [Green Version]
- Tikhonova, T.B.; Nagaeva, E.I.; Barygin, O.I.; Potapieva, N.M.; Bolshakov, K.V.; Tikhonov, D.B. Monoamine NMDA receptor channel blockers inhibit and potentiate native and recombinant proton-gated ion channels. Neuropharmacology 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Ullrich, F.; Blin, S.; Lazarow, K.; Daubitz, T.; Kries, J.P.; Jentsch, T.J. Identification of TMEM206 proteins as pore of PAORAC/ASOR acid-sensitive chloride channels. eLife 2019, 8, e49187. [Google Scholar] [CrossRef]
- Yang, J.; Chen, J.; Vitery, M.D.; Osei-Owusu, J.; Chu, J.; Yu, H.; Sun, S.; Qiu, Z. PAC, an evolutionarily conserved membrane protein, is a proton-activated chloride channel. Science 2019, 364, 395–399. [Google Scholar] [CrossRef]
- Hosford, P.; Mosienko, V.; Kishi, K.; Jurisic, G.; Seuwen, K.; Kinzel, B.; Ludwig, M.G.; Wells, J.A.; Christie, I.N.; Koolen, L.; et al. CNS distribution, signaling properties and central effects of G-protein coupled receptor 4. Neuropharmacology 2018, 138, 381–392. [Google Scholar] [CrossRef]
- Kumar, N.N.; Velic, A.; Soliz, J.; Shi, Y.; Li, K.; Wang, S.; Weaver, J.L.; Sen, J.; Abbott, S.B.G.; Lazarenko, R.M.; et al. Regulation of breathing by CO2 requires the proton-activated receptor GPR4 in retrotrapezoid nucleus neurons. Science 2015, 348, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Zhou, G.; He, M.; Xu, Y.; Rusyniak, W.G.; Xu, Y.; Ji, Y.; Simon, R.P.; Xiong, Z.G.; Zha, X. GPR68 is a neuroprotective proton receptor in brain ischemia. Stroke 2020, 51, 3690–3700. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, M.T.; Zha, X. GPR68 deletion impairs hippocampal long-term potentiation and passive avoidance behavior. Mol. Brain 2020, 13, 132. [Google Scholar] [CrossRef]
- Vollmer, L.L.; Ghosal, S.; Mcguire, J.L.; Ahlbrand, R.L.; Li, K.; Santin, J.M.; Ratliff-Rang, C.A.; Patrone, L.G.A.; Rush, J.; Lewkowich, I.P.; et al. Microglial acid sensing regulates carbon dioxide-evoked fear. Biol. Psychiatry 2016, 80, 541–551. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Modulators | Chemical and 3D Structures |
|---|---|
| Amiloride |  |
| Diminazene |  |
| APETx2 (PDB 2MUB) |  |
| PcTx1 (PDB 2KNI) |  |
| Mambalgin 2 (PDB 2MFA) |  |
| zinc |  |
| Pathologies | ASIC2 Association from Findings |
|---|---|
| Ischemic Brain Injury | |
| Multiple Sclerosis | |
| Epilepsy |
|
| Migraines |
|
| Intervertebral Disc Degeneration | |
| Arthritis | |
| Addiction |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivils, A.; Yang, F.; Wang, J.Q.; Chu, X.-P. Acid-Sensing Ion Channel 2: Function and Modulation. Membranes 2022, 12, 113. https://doi.org/10.3390/membranes12020113
Sivils A, Yang F, Wang JQ, Chu X-P. Acid-Sensing Ion Channel 2: Function and Modulation. Membranes. 2022; 12(2):113. https://doi.org/10.3390/membranes12020113
Chicago/Turabian StyleSivils, Andy, Felix Yang, John Q. Wang, and Xiang-Ping Chu. 2022. "Acid-Sensing Ion Channel 2: Function and Modulation" Membranes 12, no. 2: 113. https://doi.org/10.3390/membranes12020113