
Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antimicrobial Peptide Preparations
2.2. Circular Dichroism Spectroscopy
2.3. Antibacterial Activity Assay
2.4. Selection of the Resistant Strains
2.5. Membrane Permeability Assays
2.6. Hemolysis Assay
2.7. Cytotoxicity Assay
2.8. Measurement of Cytokine Response by Human Monocytes
3. Results
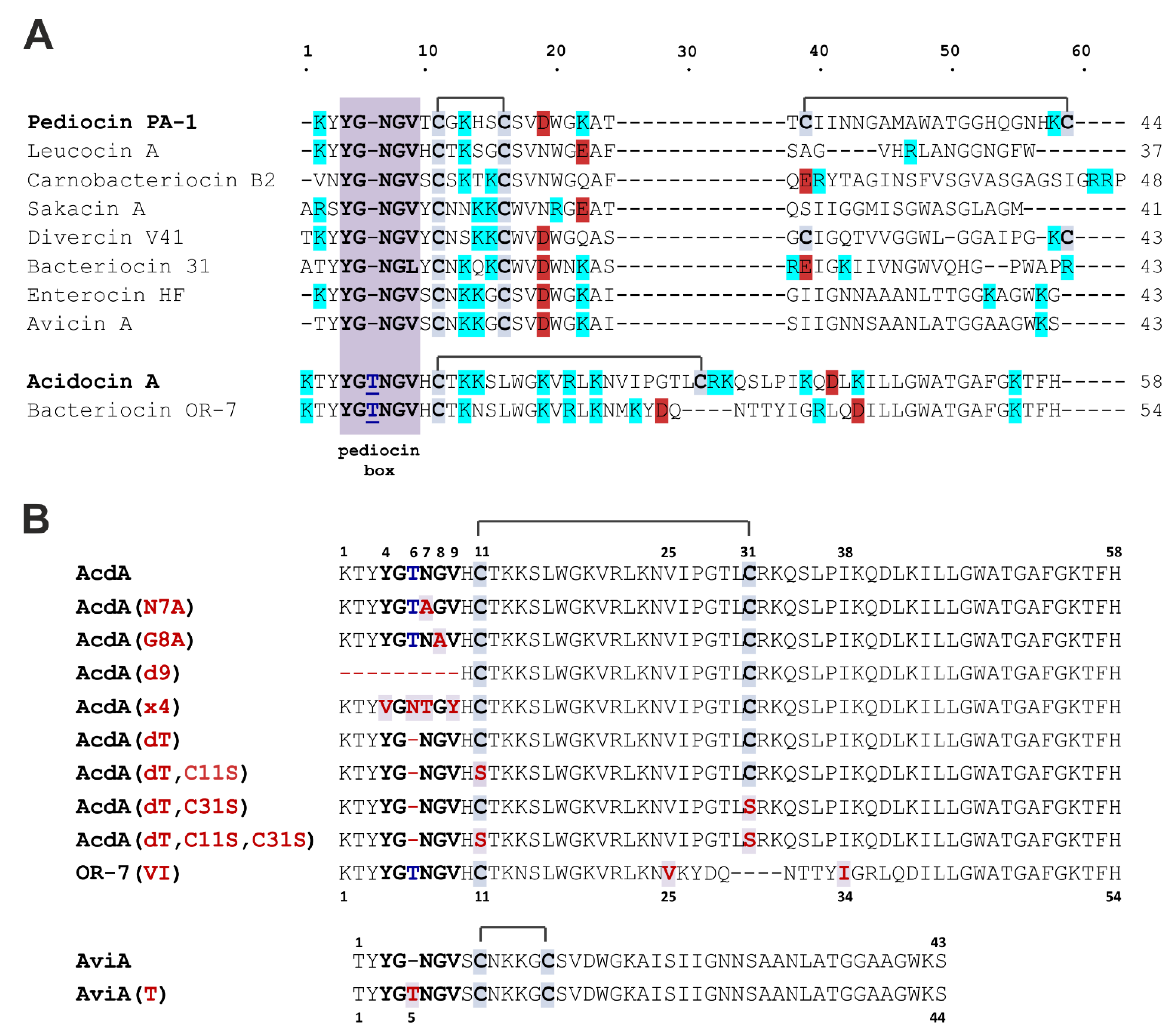
3.1. Expression and Purification of the Recombinant Peptides
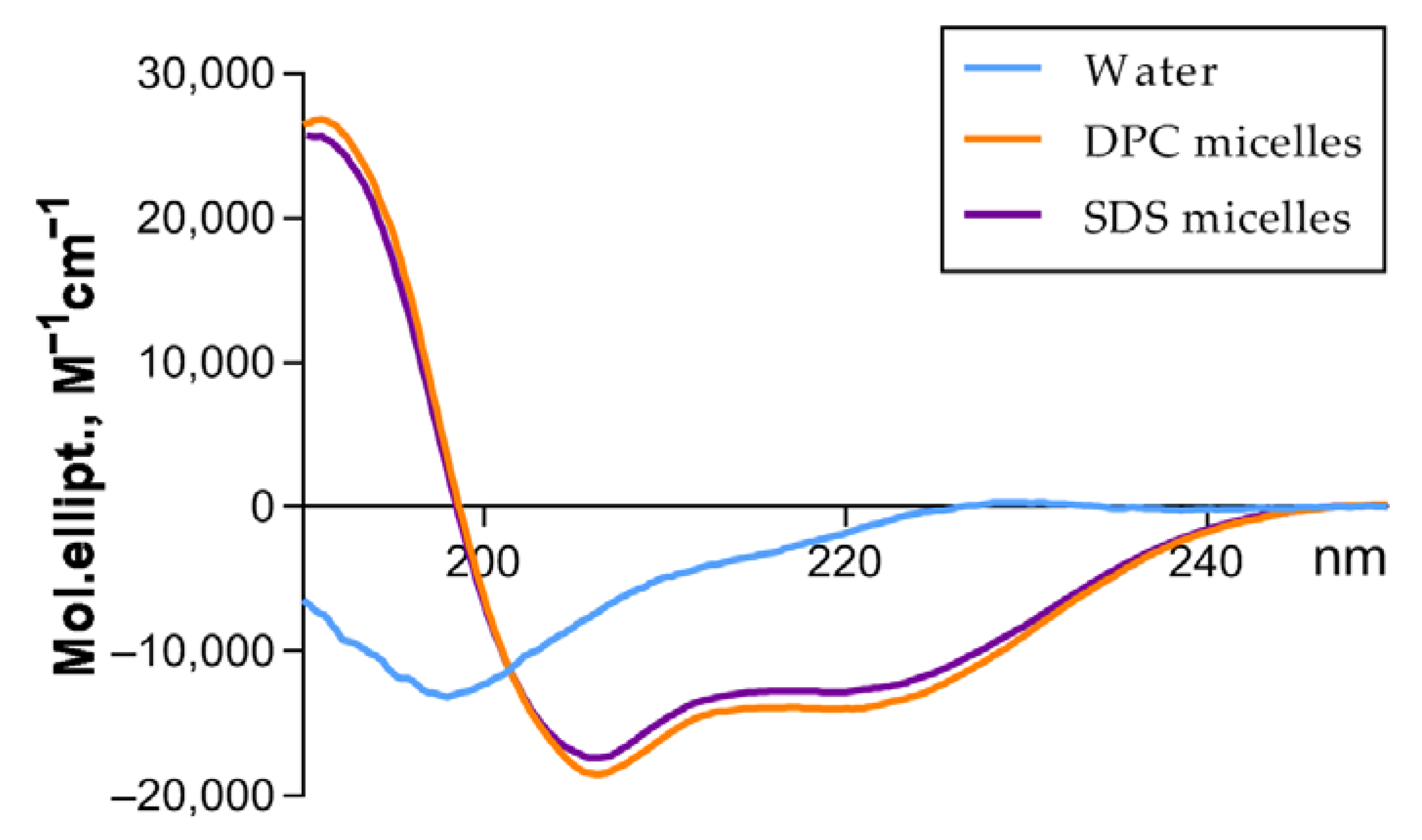
3.2. Acidocin A Secondary Structure
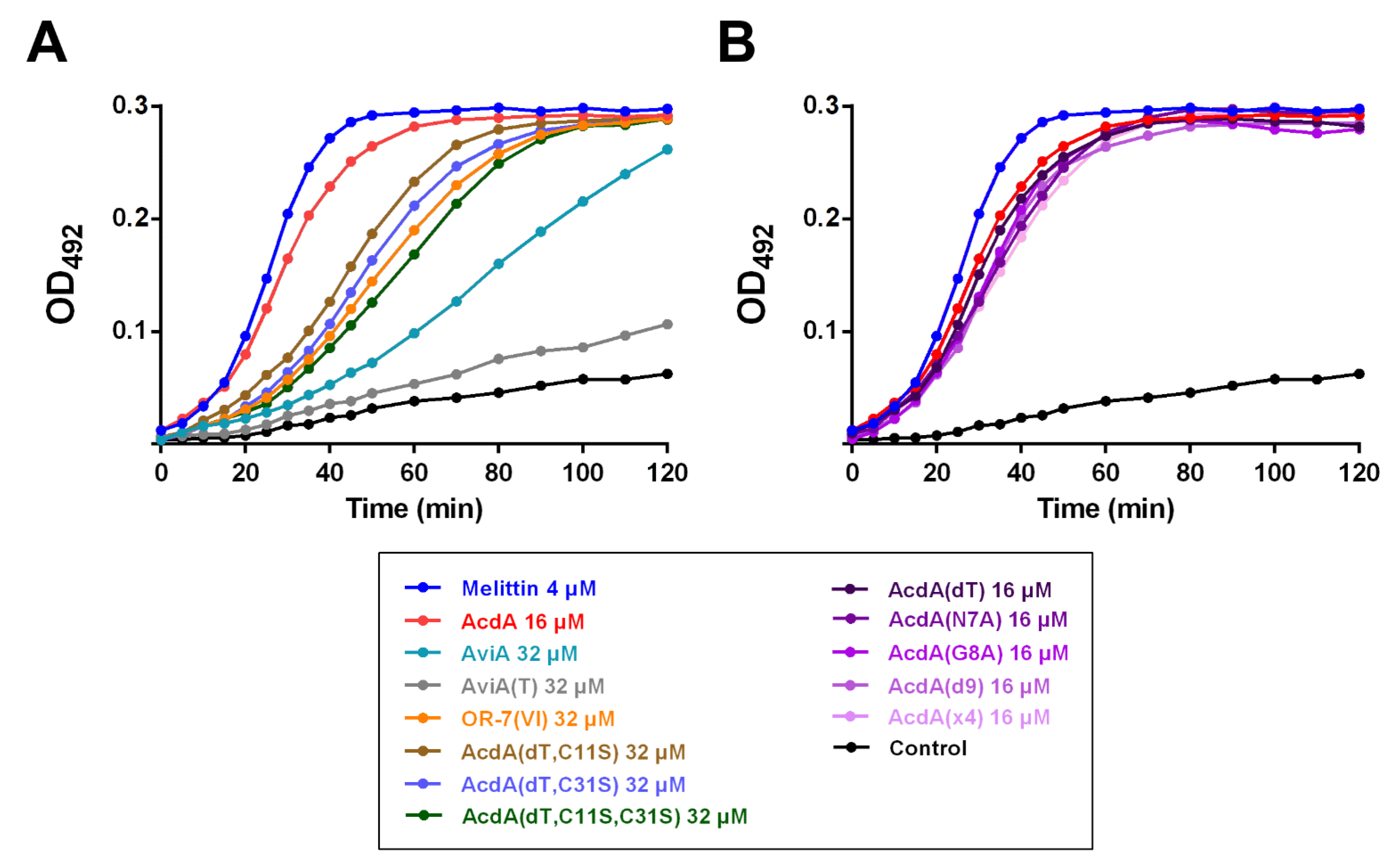
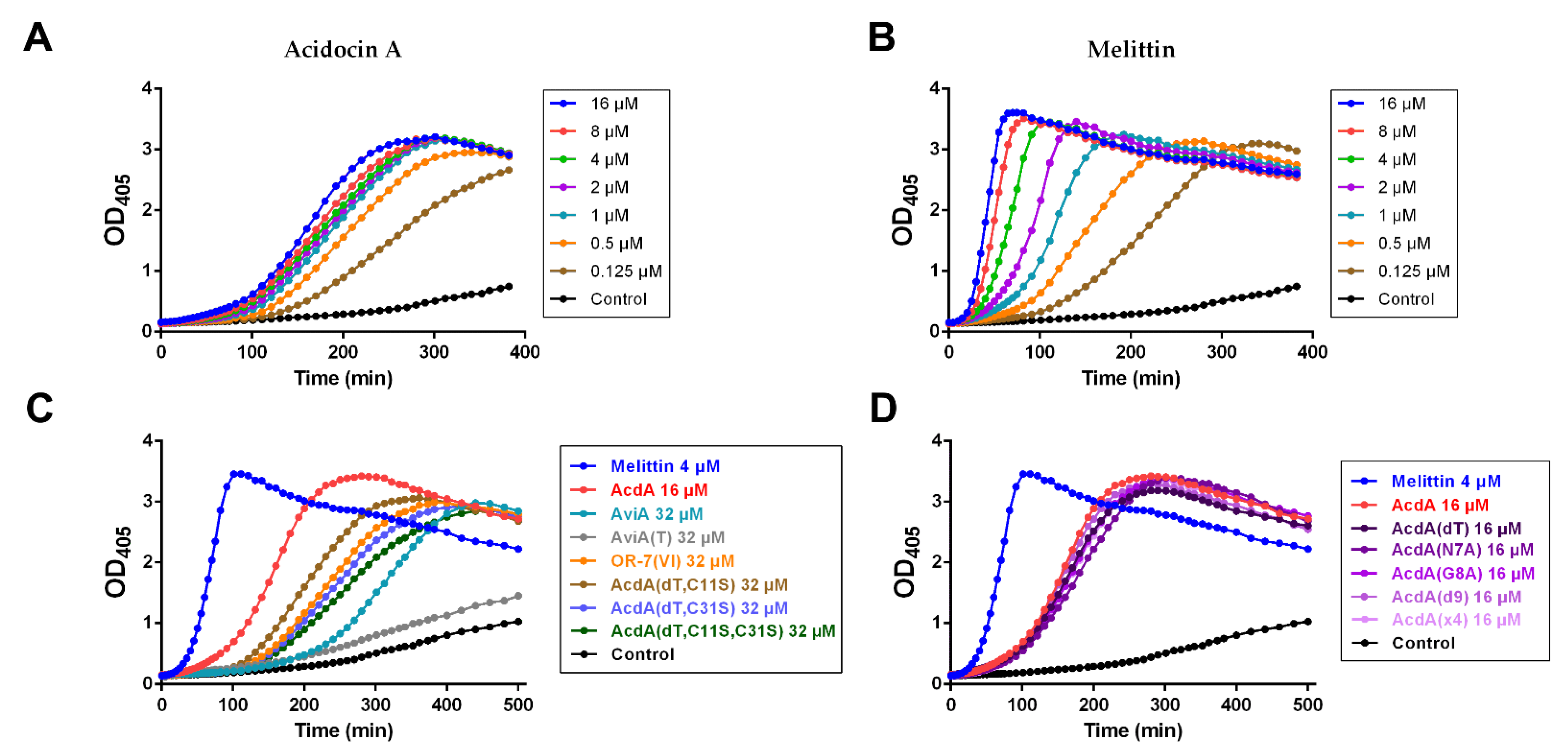
3.3. Antibacterial Activity
3.4. Bacterial Resistance to Acidocin A
3.5. Permeability of E. coli ML35p Outer and Inner Membranes
3.6. Assessment of the Hemolytic Activity and Cytotoxicity on the Human Cell Lines
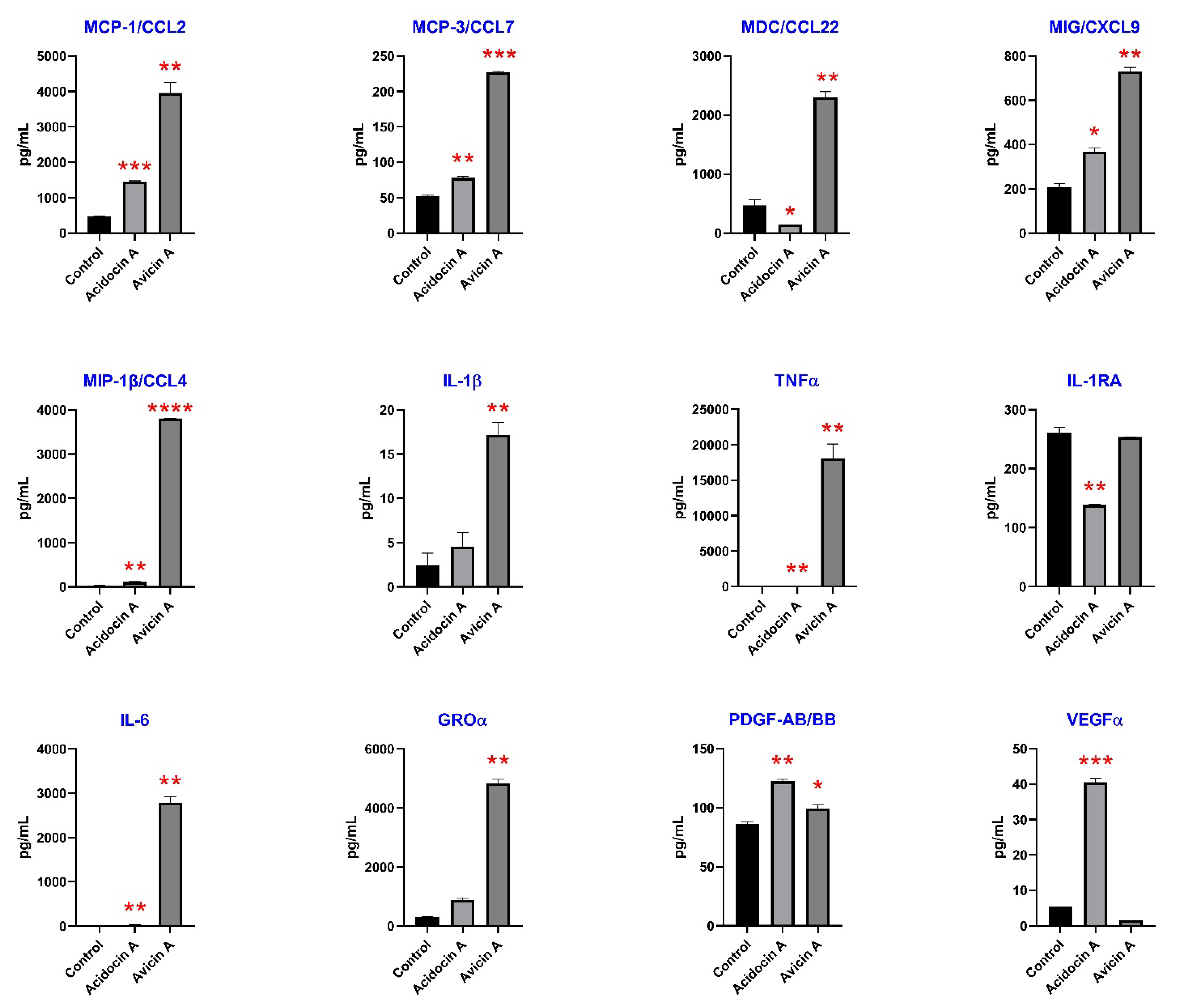
3.7. Immunomodulatory Effects on Human Immune Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yi, Y.; Li, P.; Zhao, F.; Zhang, T.; Shan, Y.; Wang, X.; Liu, B.; Chen, Y.; Zhao, X.; Lü, X. Current Status and Potentiality of Class II Bacteriocins from Lactic Acid Bacteria: Structure, Mode of Action and Applications in the Food Industry. Trends Food Sci. Technol. 2022, 120, 387–401. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.; Li, L.; Jiang, X.; Chen, Z.; Zhao, F.; Yi, Y. Biosynthesis and Production of Class II Bacteriocins of Food-Associated Lactic Acid Bacteria. Fermentation 2022, 8, 217. [Google Scholar] [CrossRef]
- Ríos Colombo, N.S.; Chalón, M.C.; Navarro, S.A.; Bellomio, A. Pediocin-like Bacteriocins: New Perspectives on Mechanism of Action and Immunity. Curr. Genet. 2018, 64, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Balandin, S.V.; Sheremeteva, E.V.; Ovchinnikova, T.V. Pediocin-Like Antimicrobial Peptides of Bacteria. Biochem. Biokhimiia 2019, 84, 464–478. [Google Scholar] [CrossRef]
- McClintock, M.K.; Kaznessis, Y.N.; Hackel, B.J. Enterocin A Mutants Identified by Saturation Mutagenesis Enhance Potency towards Vancomycin-Resistant Enterococci. Biotechnol. Bioeng. 2016, 113, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Farias, F.M.; Teixeira, L.M.; Vallim, D.C.; Bastos, M.D.C.D.F.; Miguel, M.A.L.; Bonelli, R.R. Characterization of Enterococcus Faecium E86 Bacteriocins and Their Inhibition Properties against Listeria Monocytogenes and Vancomycin-Resistant Enterococcus. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2021, 52, 1513–1522. [Google Scholar] [CrossRef]
- Zhu, L.; Zeng, J.; Wang, C.; Wang, J. Structural Basis of Pore Formation in the Mannose Phosphotransferase System by Pediocin PA-1. Appl. Environ. Microbiol. 2022, 88, e0199221. [Google Scholar] [CrossRef]
- Aymerich, T.; Jofré, A.; Bover-Cid, S. Enterocin A-Based Antimicrobial Film Exerted Strong Antilisterial Activity in Sliced Dry-Cured Ham Immediately and after 6 Months at 8 °C. Food Microbiol. 2022, 105, 104005. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a New Generation of Antimicrobials: Toxicity Aspects and Regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef]
- Kanatani, K.; Oshimura, M.; Sano, K. Isolation and Characterization of Acidocin A and Cloning of the Bacteriocin Gene from Lactobacillus Acidophilus. Appl. Environ. Microbiol. 1995, 61, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus Salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter Jejuni in the Chicken Gastrointestinal System. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birri, D.J.; Brede, D.A.; Forberg, T.; Holo, H.; Nes, I.F. Molecular and Genetic Characterization of a Novel Bacteriocin Locus in Enterococcus Avium Isolates from Infants. Appl. Environ. Microbiol. 2010, 76, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balandin, S.V.; Finkina, E.I.; Nurmukhamedova, E.K.-A.; Tagaev, A.A.; Umnyakova, E.S.; Kokryakov, V.N.; Shvets, V.I.; Ovchinnikova, T.V. Biotechnological Method of Preparation and Characterization of Recombinant Antimicrobial Peptide Avicin A from Enterococcus Avium. Dokl. Biochem. Biophys. 2019, 484, 42–44. [Google Scholar] [CrossRef]
- Puigbò, P.; Guzmán, E.; Romeu, A.; Garcia-Vallvé, S. OPTIMIZER: A Web Server for Optimizing the Codon Usage of DNA Sequences. Nucleic Acids Res. 2007, 35, W126–W131. [Google Scholar] [CrossRef] [Green Version]
- Shenkarev, Z.O.; Panteleev, P.V.; Balandin, S.V.; Gizatullina, A.K.; Altukhov, D.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S.; Ovchinnikova, T.V. Recombinant Expression and Solution Structure of Antimicrobial Peptide Aurelin from Jellyfish Aurelia Aurita. Biochem. Biophys. Res. Commun. 2012, 429, 63–69. [Google Scholar] [CrossRef]
- Jones, D.H.; Howard, B.H. A Rapid Method for Recombination and Site-Specific Mutagenesis by Placing Homologous Ends on DNA Using Polymerase Chain Reaction. BioTechniques 1991, 10, 62–66. [Google Scholar]
- Suttapitugsakul, S.; Xiao, H.; Smeekens, J.; Wu, R. Evaluation and Optimization of Reduction and Alkylation Methods to Maximize Peptide Identification with MS-Based Proteomics. Mol. Biosyst. 2017, 13, 2574–2582. [Google Scholar] [CrossRef]
- Magalhães, P.O.; Lopes, A.M.; Mazzola, P.G.; Rangel-Yagui, C.; Penna, T.C.V.; Pessoa, A. Methods of Endotoxin Removal from Biological Preparations: A Review. J. Pharm. Pharm. Sci. Publ. Can. Soc. Pharm. Sci. Soc. Can. Sci. Pharm. 2007, 10, 388–404. [Google Scholar]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Nadezhdin, K.D.; Balandin, S.V.; Zhmak, M.N.; Kudelina, I.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S. Recombinant Expression, Synthesis, Purification, and Solution Structure of Arenicin. Biochem. Biophys. Res. Commun. 2007, 360, 156–162. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to Study Proteins by Circular Dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.T. Circular Dichroism and Its Use in Protein-Folding Studies. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 752, pp. 59–72. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Tagg, J.R.; Dajani, A.S.; Wannamaker, L.W. Bacteriocins of Gram-Positive Bacteria. Bacteriol. Rev. 1976, 40, 722–756. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Hu, J.; Ke, F. Experimental Induction of Bacterial Resistance to the Antimicrobial Peptide Tachyplesin I and Investigation of the Resistance Mechanisms. Antimicrob. Agents Chemother. 2016, 60, 6067–6075. [Google Scholar] [CrossRef] [Green Version]
- Shamova, O.V.; Orlov, D.S.; Zharkova, M.S.; Balandin, S.V.; Yamschikova, E.V.; Knappe, D.; Hoffmann, R.; Kokryakov, V.N.; Ovchinnikova, T.V. Minibactenecins ChBac7.Nα and ChBac7. Nβ-Antimicrobial Peptides from Leukocytes of the Goat Capra Hircus. Acta Nat. 2016, 8, 136–146. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Kalashnikov, A.À.; Kokryakov, V.N.; Shamova, O.V.; Emelianova, A.A.; Balandin, S.V.; Ovchinnikova, T.V. Combined Antibacterial Effects of Goat Cathelicidins with Different Mechanisms of Action. Front. Microbiol. 2018, 9, 2983. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (Resazurin) Fluorescent Dye for the Assessment of Mammalian Cell Cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Rampersad, S.N. Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Wahl, L.M.; Smith, P.D. Isolation of Monocyte/Macrophage Populations. Curr. Protoc. Immunol. 1995, 16, 7.6.1–7.6.8. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Mantovani, A.; Gray, P.A.; Van Damme, J.; Sozzani, S. Macrophage-Derived Chemokine (MDC). J. Leukoc. Biol. 2000, 68, 400–404. [Google Scholar] [PubMed]
- Vries, M.H.M.; Wagenaar, A.; Verbruggen, S.E.L.; Molin, D.G.M.; Dijkgraaf, I.; Hackeng, T.H.; Post, M.J. CXCL1 Promotes Arteriogenesis through Enhanced Monocyte Recruitment into the Peri-Collateral Space. Angiogenesis 2015, 18, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, G.M.; Davidson, B.E.; Hillier, A.J. Novel Expression System for Large-Scale Production and Purification of Recombinant Class IIa Bacteriocins and Its Application to Piscicolin 126. Appl. Environ. Microbiol. 2004, 70, 3292–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazazic, M.; Nissen-Meyer, J.; Fimland, G. Mutational Analysis of the Role of Charged Residues in Target-Cell Binding, Potency and Specificity of the Pediocin-like Bacteriocin Sakacin P. Microbiol. Read. Engl. 2002, 148, 2019–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial Peptides: Interaction With Model and Biological Membranes and Synergism With Chemical Antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldhuizen, E.J.A.; Schneider, V.A.F.; Agustiandari, H.; van Dijk, A.; Tjeerdsma-van Bokhoven, J.L.M.; Bikker, F.J.; Haagsman, H.P. Antimicrobial and Immunomodulatory Activities of PR-39 Derived Peptides. PLoS ONE 2014, 9, e95939. [Google Scholar] [CrossRef]
- Cui, G.; Pan, C.; Xu, P.; Li, Y.; Wang, L.; Gong, B.; Li, X.; Huang, S. Purification and Characterization of a Novel Bacteriocin Produced by Enterococcus Faecalis CG-9 from Human Saliva. Biotechnol. Biotechnol. Equip. 2020, 34, 1224–1233. [Google Scholar] [CrossRef]
- Kjos, M.; Nes, I.F.; Diep, D.B. Class II One-Peptide Bacteriocins Target a Phylogenetically Defined Subgroup of Mannose Phosphotransferase Systems on Sensitive Cells. Microbiol. Read. Engl. 2009, 155, 2949–2961. [Google Scholar] [CrossRef] [Green Version]
- Jeckelmann, J.-M.; Erni, B. The Mannose Phosphotransferase System (Man-PTS)-Mannose Transporter and Receptor for Bacteriocins and Bacteriophages. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183412. [Google Scholar] [CrossRef]
- Zhu, L.; Zeng, J.; Wang, J. Structural Basis of the Immunity Mechanisms of Pediocin-like Bacteriocins. Appl. Environ. Microbiol. 2022, 88, e0048122. [Google Scholar] [CrossRef]
- Tymoszewska, A.; Diep, D.B.; Wirtek, P.; Aleksandrzak-Piekarczyk, T. The Non-Lantibiotic Bacteriocin Garvicin Q Targets Man-PTS in a Broad Spectrum of Sensitive Bacterial Genera. Sci. Rep. 2017, 7, 8359. [Google Scholar] [CrossRef] [PubMed]
- Tymoszewska, A.; Walczak, P.; Aleksandrzak-Piekarczyk, T. BacSJ-Another Bacteriocin with Distinct Spectrum of Activity That Targets Man-PTS. Int. J. Mol. Sci. 2020, 21, 7860. [Google Scholar] [CrossRef] [PubMed]
- Assoni, L.; Milani, B.; Carvalho, M.R.; Nepomuceno, L.N.; Waz, N.T.; Guerra, M.E.S.; Converso, T.R.; Darrieux, M. Resistance Mechanisms to Antimicrobial Peptides in Gram-Positive Bacteria. Front. Microbiol. 2020, 11, 2362. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, T.; Hatakeyama, Y. Determination of Essential and Variable Residues in Pediocin PA-1 by NNK Scanning. Appl. Environ. Microbiol. 2006, 72, 1141–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, X.; Chen, X.; Du, L.; Wu, X.; Liu, F.; Yuan, J. Alanine-Scanning Mutational Analysis of Durancin GL Reveals Residues Important for Its Antimicrobial Activity. J. Agric. Food Chem. 2015, 63, 6402–6409. [Google Scholar] [CrossRef]
- Flynn, J.; Ryan, A.; Hudson, S.P. Pre-Formulation and Delivery Strategies for the Development of Bacteriocins as next Generation Antibiotics. Eur. J. Pharm. Biopharm. Off. J. Arb. Pharm. Verfahr. EV 2021, 165, 149–163. [Google Scholar] [CrossRef]
- Soltani, S.; Zirah, S.; Rebuffat, S.; Couture, F.; Boutin, Y.; Biron, E.; Subirade, M.; Fliss, I. Gastrointestinal Stability and Cytotoxicity of Bacteriocins From Gram-Positive and Gram-Negative Bacteria: A Comparative in Vitro Study. Front. Microbiol. 2021, 12, 780355. [Google Scholar] [CrossRef]
- Baindara, P.; Chaudhry, V.; Mittal, G.; Liao, L.M.; Matos, C.O.; Khatri, N.; Franco, O.L.; Patil, P.B.; Korpole, S. Characterization of the Antimicrobial Peptide Penisin, a Class Ia Novel Lantibiotic from Paenibacillus Sp. Strain A3. Antimicrob. Agents Chemother. 2016, 60, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Shang, L.; Yang, G.; Dai, Z.; Zeng, X.; Qiao, S. Biosynthetic Microcin J25 Exerts Strong Antibacterial, Anti-Inflammatory Activities, Low Cytotoxicity without Increasing Drug-Resistance to Bacteria Target. Front. Immunol. 2022, 13, 811378. [Google Scholar] [CrossRef]
- Niklison Chirou, M.; Bellomio, A.; Dupuy, F.; Arcuri, B.; Minahk, C.; Morero, R. Microcin J25 Induces the Opening of the Mitochondrial Transition Pore and Cytochrome c Release through Superoxide Generation. FEBS J. 2008, 275, 4088–4096. [Google Scholar] [CrossRef]
- Makino, A.; Baba, T.; Fujimoto, K.; Iwamoto, K.; Yano, Y.; Terada, N.; Ohno, S.; Sato, S.B.; Ohta, A.; Umeda, M.; et al. Cinnamycin (Ro 09-0198) Promotes Cell Binding and Toxicity by Inducing Transbilayer Lipid Movement. J. Biol. Chem. 2003, 278, 3204–3209. [Google Scholar] [CrossRef] [PubMed]
- Śmiałek, J.; Bzowska, M.; Hinz, A.; Mężyk-Kopeć, R.; Sołtys, K.; Mak, P. Bacteriocin BacSp222 and Its Succinylated Forms Exhibit Proinflammatory Activities Toward Innate Immune Cells. J. Inflamm. Res. 2022, 15, 4601–4621. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of Innate Immunity by a Bacterial Secreted Peptide: Lantibiotic Nisin Z Is Selectively Immunomodulatory. Innate Immun. 2013, 19, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Rękawek, W.; Siwicki, A.K. In Vitro Immunomodulatory Effect of Nisin on Porcine Leucocytes. J. Anim. Physiol. Anim. Nutr. 2019, 103, 882–893. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.; Eubank, T.D.; Doseff, A.I. Monocytes and Macrophages Regulate Immunity through Dynamic Networks of Survival and Cell Death. J. Innate Immun. 2010, 2, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouritzen, M.V.; Andrea, A.; Qvist, K.; Poulsen, S.S.; Jenssen, H. Immunomodulatory Potential of Nisin A with Application in Wound Healing. Wound Repair Regen. 2019, 27, 650–660. [Google Scholar] [CrossRef]
- Guo, S.; Dipietro, L.A. Factors Affecting Wound Healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The Role of Vascular Endothelial Growth Factor in Wound Healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Orelle, C.; Carlson, S.; Kaushal, B.; Almutairi, M.M.; Liu, H.; Ochabowicz, A.; Quan, S.; Pham, V.C.; Squires, C.L.; Murphy, B.T.; et al. Tools for Characterizing Bacterial Protein Synthesis Inhibitors. Antimicrob. Agents Chemother. 2013, 57, 5994–6004. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteriocin | Condition | α-Helix, % | β-Sheet, % | β-Turn, % | Random, % | NRMSD |
|---|---|---|---|---|---|---|
| Acidocin A | Aqueous solution | 5.7 | 33.7 | 22.6 | 37.9 | 0.02 |
| DPC micelles | 44.0 | 8.1 | 19.2 | 28.6 | 0.01 | |
| SDS micelles | 40.2 | 10.6 | 20.3 | 28.8 | 0.01 | |
| Avicin A * | Aqueous solution | 5.8 | 32.8 | 22.1 | 39.3 | 0.02 |
| DPC micelles | 19.4 | 28.6 | 21.6 | 27.8 | 0.02 | |
| SDS micelles | 19.7 | 29.3 | 23.1 | 30.4 | 0.02 |
| Bacteriocins | Minimum Inhibitory Concentration (µM) * | |||||||
|---|---|---|---|---|---|---|---|---|
| L. monocytogenes EGD | L. lactis MK66 | L. lactis MK43 | B. subtilis B-886 | B. licheniformis B-511 | M. phlei Ac-1221 | M. luteus Ac-2229 | S. aureus 209P | |
| Acidocin A | >128 | 0.5 | 1 | 2 | 2 | 2 | 8 | 8 |
| AcdA(dT) | >128 | 0.5 | 1 | 2 | 2 | 2 | 8 | 8 |
| AcdA (DTT reduced) | >32 | 1 | 2 | 4 | 4 | 4 | 16 | 16 |
| AcdA(dT) (DTT reduced) | >32 | 1 | 2 | 4 | 4 | 4 | 16 | 16 |
| AcdA(dT,C11S) | >32 | 1 | 4 | 4 | 4 | 4 | 16 | 32 |
| AcdA(dT,C31S) | >32 | 1 | 2 | 4 | 4 | 4 | 16 | 32 |
| AcdA(dT,C11S,C31S) | >32 | 1 | 4 | 4 | 4 | 8 | 32 | >32 |
| AcdA(N7A) | >32 | 2 | 8 | 8 | 4 | 2 | 16 | 8 |
| AcdA(G8A) | >32 | 2 | 4 | 8 | 8 | 2 | 16 | 8 |
| AcdA(d9) | >32 | 2 | 8 | 8 | 8 | 4 | 16 | 16 |
| AcdA(x4) | >32 | 2 | 4 | 8 | 4 | 4 | 8 | 16 |
| OR-7(VI) | >64 | 4 | >32 | 16 | 8 | 32 | >32 | >32 |
| OR-7(VI) (DTT reduced) | >32 | 4 | >32 | 16 | 8 | 32 | >32 | >32 |
| Avicin A ** | <0.125 | >32 | >32 | >32 | >32 | >32 | >32 | >32 |
| AviA(T) | >32 | >32 | >32 | >32 | >32 | >32 | >32 | >32 |
| Bacteriocins | Minimum Inhibitory Concentration (µM) * | |||||
|---|---|---|---|---|---|---|
| E. coli ML-35p | E. coli ATCC 25922 | E. coli SQ110 | E. coli XDR CI 1057 | P. aeruginosa MDR CI 1995 | A. baumannii XDR CI 2675 | |
| AcdA | 2 | 4 | 2 | 8 | 16 | 4 |
| AcdA(dT) | 2 | 4 | 1 | 8 | 16 | 4 |
| AcdA (DTT reduced) | 4 | 8 | 4 | 16 | >32 | 8 |
| AcdA(dT) (DTT reduced) | 4 | 8 | 4 | 16 | >32 | 8 |
| AcdA(dT,C11S) | 4 | 8 | 4 | 16 | >32 | 8 |
| AcdA(dT,C31S) | 8 | 8 | 8 | 16 | >32 | 16 |
| AcdA(dT,C11S,C31S) | >64 | >64 | 16 | >64 | >32 | >32 |
| AcdA(N7A) | 8 | 16 | 8 | 8 | 16 | 4 |
| AcdA(G8A) | 8 | 16 | 4 | 8 | 16 | 8 |
| AcdA(d9) | 4 | 16 | 8 | 8 | 16 | 4 |
| AcdA(x4) | 8 | 16 | 8 | 8 | 16 | 4 |
| OR-7(VI) | >32 | >32 | >64 | >64 | >32 | >32 |
| OR-7(VI) (DTT reduced) | >32 | >32 | >32 | >32 | >32 | >32 |
| AviA | >32 | >32 | >32 | >32 | >32 | >32 |
| AviA(T) | >32 | >32 | >32 | >32 | >32 | >32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoshina, D.V.; Balandin, S.V.; Bogdanov, I.V.; Vershinina, M.A.; Sheremeteva, E.V.; Toropygin, I.Y.; Finkina, E.I.; Ovchinnikova, T.V. Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure. Membranes 2022, 12, 1253. https://doi.org/10.3390/membranes12121253
Antoshina DV, Balandin SV, Bogdanov IV, Vershinina MA, Sheremeteva EV, Toropygin IY, Finkina EI, Ovchinnikova TV. Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure. Membranes. 2022; 12(12):1253. https://doi.org/10.3390/membranes12121253
Chicago/Turabian StyleAntoshina, Daria V., Sergey V. Balandin, Ivan V. Bogdanov, Maria A. Vershinina, Elvira V. Sheremeteva, Ilia Yu. Toropygin, Ekaterina I. Finkina, and Tatiana V. Ovchinnikova. 2022. "Antimicrobial Activity and Immunomodulatory Properties of Acidocin A, the Pediocin-like Bacteriocin with the Non-Canonical Structure" Membranes 12, no. 12: 1253. https://doi.org/10.3390/membranes12121253