
Ameliorative Effects of Camel Milk and Its Exosomes on Diabetic Nephropathy in Rats
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Milk-Derived Exosomes
2.2. Characterization of Milk-Derived Exosomes
2.3. Chemical Induction of Diabetic Nephropathy
2.4. Experimental Design
2.5. Sampling
2.6. Biochemical Parameters
2.7. Real-Time PCR
2.8. Histopathology
2.9. Statistical Analysis
3. Results
3.1. Characterization of CM-Derived EXOs
3.2. Effects of CM and/or Its EXOs on Kidney Damage Parameters
3.3. Effects of CM and/or Its EXOs on Oxidative Stress and Antioxidants Parameters
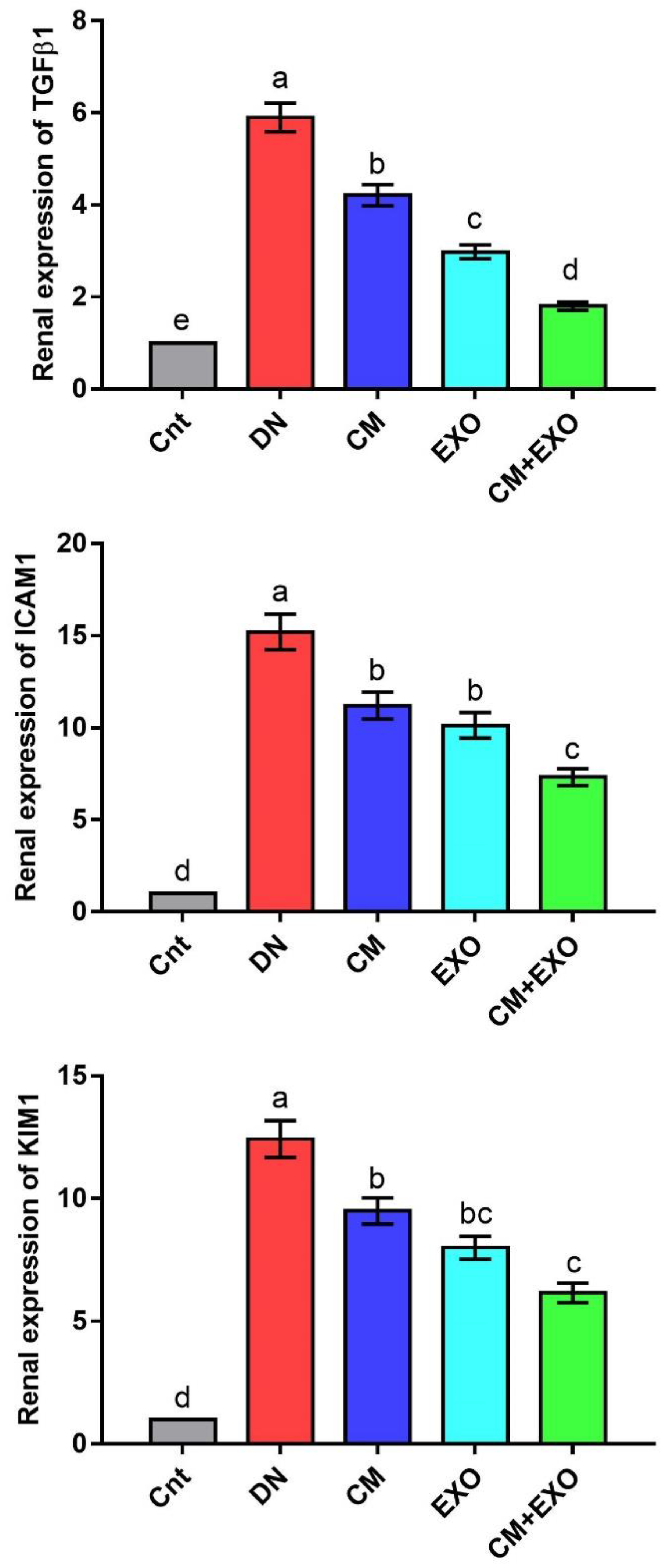
3.4. Effect of CM and/or Its EXOs on Fibrosis and Kidney Damage-Related Genes
3.5. Effect of CM and/or Its EXOs on Matrix Remodeling-Related Genes
3.6. Effects of CM and/or Its EXOs on Renal Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Woodhams, L.; Sim, T.F.; Chalmers, L.; Yeap, B.; Green, D.; Schlaich, M.; Schultz, C.; Hillis, G. Diabetic kidney disease in type 2 diabetes: A review of pathogenic mechanisms, patient-related factors and therapeutic options. PeerJ 2021, 9, e11070. [Google Scholar] [CrossRef]
- Shao, B.Y.; Zhang, S.F.; Li, H.D.; Meng, X.M.; Chen, H.Y. Epigenetics and Inflammation in Diabetic Nephropathy. Front. Physiol. 2021, 12, 649587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, R.; Shi, W.; Liang, X.; Liu, S.; Ye, Z.; Yu, C.; Chen, Y.; Zhang, B.; Wang, W. NFAT2 inhibitor ameliorates diabetic nephropathy and podocyte injury in db/db mice. Br. J. Pharmacol. 2013, 170, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Ajay, A.K.; Chang, J.H.; Mou, S.; Zhao, H.; Kishi, S.; Li, J.; Brooks, C.R.; Xiao, S.; Woo, H.M.; et al. KIM-1 mediates fatty acid uptake by renal tubular cells to promote progressive diabetic kidney disease. Cell Metab. 2021, 33, 1042–1061.e1047. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.D.; Wang, W.W.; Feng, Z.; Liu, R.; Cheng, X.L.; Shen, W.J.; Dong, Z.Y.; Cai, G.Y.; Chen, X.M.; Hong, Q.; et al. Identification of key genes and pathways in diabetic nephropathy by bioinformatics analysis. J. Diabetes Investig. 2019, 10, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-x.; Liu, X.-m.; Su, Y.; Zhang, X.-J. Renal expression of proto-oncogene Ets-1 on matrix remodeling in experimental diabetic nephropathy. Acta Histochem. 2011, 113, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Rayego-Mateos, S.; Morgado-Pascual, J.L.; Opazo-Rios, L.; Guerrero-Hue, M.; Garcia-Caballero, C.; Vazquez-Carballo, C.; Mas, S.; Sanz, A.B.; Herencia, C.; Mezzano, S.; et al. Pathogenic Pathways and Therapeutic Approaches Targeting Inflammation in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 3798. [Google Scholar] [CrossRef]
- Yoshimura, E.H.; Santos, N.W.; Machado, E.; Agustinho, B.C.; Pereira, L.M.; de Aguiar, S.C.; Sá-Nakanishi, A.B.; Mareze-da-Costa, C.E.; Zeoula, L.M. Functionality of cow milk naturally enriched with polyunsaturated fatty acids and polyphenols in diets for diabetic rats. PLoS ONE 2018, 13, e0195839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alu’datt, M.H.; Al-u’datt, D.a.G.F.; Alhamad, M.N.; Tranchant, C.C.; Rababah, T.; Gammoh, S.; Althnaibat, R.M.; Daradkeh, M.G.; Kubow, S. Characterization and biological properties of peptides isolated from dried fermented cow milk products by RP-HPLC: Amino acid composition, antioxidant, antihypertensive, and antidiabetic properties. J. Food Sci. 2021, 86, 3046–3060. [Google Scholar] [CrossRef] [PubMed]
- Shori, A.B. Camel milk as a potential therapy for controlling diabetes and its complications: A review of in vivo studies. J. Food Drug Anal. 2015, 23, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Jain, S.; Shah, S.; Chopra, A.; Agarwal, V. Effect of camel milk on glycemic control and insulin requirement in patients with type 1 diabetes: 2-years randomized controlled trial. Eur. J. Clin. Nutr. 2011, 65, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Sboui, A.; Khorchani, T.; Djegham, M.; Agrebi, A.; Elhatmi, H.; Belhadj, O. Anti-diabetic effect of camel milk in alloxan-induced diabetic dogs: A dose–response experiment. J. Anim. Physiol. Anim. Nutr. 2010, 94, 540–546. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Budania, S.; Sharma, P.; Gupta, R.; Kochar, D.K.; Panwar, R.B.; Sahani, M.S. Zero prevalence of diabetes in camel milk consuming Raica community of north-west Rajasthan, India. Diabetes Res. Clin. Pract. 2007, 76, 290–296. [Google Scholar] [CrossRef]
- Mohamad, R.H.; Zekry, Z.K.; Al-Mehdar, H.A.; Salama, O.; El-Shaieb, S.E.; El-Basmy, A.A.; Al-said, M.G.; Sharawy, S.M. Camel milk as an adjuvant therapy for the treatment of type 1 diabetes: Verification of a traditional ethnomedical practice. J. Med. Food 2009, 12, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Alzohairy, M.A.; Mohieldein, A.H. Antidiabetic effects of camel milk in streptozotocin-induced diabetic rats. Am. J. Biochem. Mol. Biol. 2013, 3, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Korish, A. The antidiabetic action of camel milk in experimental type 2 diabetes mellitus: An overview on the changes in incretin hormones, insulin resistance, and inflammatory cytokines. Horm. Metab. Res. 2014, 46, 404–411. [Google Scholar] [CrossRef]
- Hull, S. Camel’s milk to treat diabetes. Nature 2004, 363, 446–448. [Google Scholar]
- Malik, A.; Al-Senaidy, A.; Skrzypczak-Jankun, E.; Jankun, J. A study of the anti-diabetic agents of camel milk. Int. J. Mol. Med. 2012, 30, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Mal, G.; Kumar, D.; Patil, N.V.; Pathak, K.M.L. Camel Milk: An Important Natural Adjuvant. Agric. Res. 2017, 6, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Gader, A.G.M.A.; Alhaider, A.A. The unique medicinal properties of camel products: A review of the scientific evidence. J. Taibah Univ. Med. Sci. 2016, 11, 98–103. [Google Scholar]
- Alzahrani, F.A.; El-Magd, M.A.; Abdelfattah-Hassan, A.; Saleh, A.A.; Saadeldin, I.M.; El-Shetry, E.S.; Badawy, A.A.; Alkarim, S. Potential Effect of Exosomes Derived from Cancer Stem Cells and MSCs on Progression of DEN-Induced HCC in Rats. Stem Cells Int. 2018, 2018, 8058979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzahrani, F.A.; Shait Mohammed, M.R.; Alkarim, S.; Azhar, E.I.; El-Magd, M.A.; Hawsawi, Y.; Abdulaal, W.H.; Yusuf, A.; Alhatmi, A.; Albiheyri, R.; et al. Untargeted Metabolic Profiling of Extracellular Vesicles of SARS-CoV-2-Infected Patients Shows Presence of Potent Anti-Inflammatory Metabolites. Int. J. Mol. Sci. 2021, 22, 10467. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, I.; Thakur, A.; Iyaswamy, A.; Wang, X.; Chen, X.; Yang, Z. Factors Affecting Extracellular Vesicles Based Drug Delivery Systems. Molecules 2021, 26, 1544. [Google Scholar] [CrossRef]
- Badawy, A.A.; Othman, R.Q.A.; El-Magd, M.A. Effect of combined therapy with camel milk-derived exosomes, tamoxifen, and hesperidin on breast cancer. Mol. Cell. Toxicol. 2021, 1–10. [Google Scholar] [CrossRef]
- Othman, R.; Badawy, A.; Alruwaili, M.; El-Magd, M. Camel Milk Exosomes Potentiate The Anticancer Effect of Doxorubicin on Multidrug-Resistant Human Leukemia Hl60 Cells in Vitro and in Vivo. Pak. J. Med. Health Sci. 2021, 15, 3313–3320. [Google Scholar] [CrossRef]
- Thakur, A.; Johnson, A.; Jacobs, E.; Zhang, K.; Chen, J.; Wei, Z.; Lian, Q.; Chen, H.J. Energy Sources for Exosome Communication in a Cancer Microenvironment. Cancers 2022, 14, 1698. [Google Scholar] [CrossRef]
- Qu, X.; Leung, T.C.N.; Ngai, S.-M.; Tsai, S.-N.; Thakur, A.; Li, W.-K.; Lee, Y.; Leung, L.; Ng, T.-H.; Yam, J.; et al. Proteomic Analysis of Circulating Extracellular Vesicles Identifies Potential Biomarkers for Lymph Node Metastasis in Oral Tongue Squamous Cell Carcinoma. Cells 2021, 10, 2179. [Google Scholar] [CrossRef]
- Van Hese, I.; Goossens, K.; Vandaele, L.; Opsomer, G. Invited review: MicroRNAs in bovine colostrum—Focus on their origin and potential health benefits for the calf. J. Dairy Sci. 2020, 103, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J. Dairy Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef] [Green Version]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Pieters, B.C.; Arntz, O.J.; Bennink, M.B.; Broeren, M.G.; van Caam, A.P.; Koenders, M.I.; van Lent, P.L.; van den Berg, W.B.; de Vries, M.; van der Kraan, P.M. Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-β. PLoS ONE 2015, 10, e0121123. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Mohammed-Geba, K.; Tawfic, A.A.; El-Magd, M.A. Camel milk exosomes modulate cyclophosphamide-induced oxidative stress and immuno-toxicity in rats. Food Funct. 2019, 10, 7523–7532. [Google Scholar] [CrossRef] [PubMed]
- Cianciaruso, C.; Phelps, E.A.; Pasquier, M.; Hamelin, R.; Demurtas, D.; Alibashe Ahmed, M.; Piemonti, L.; Hirosue, S.; Swartz, M.A.; De Palma, M. Primary human and rat β-cells release the intracellular autoantigens GAD65, IA-2, and proinsulin in exosomes together with cytokine-induced enhancers of immunity. Diabetes 2017, 66, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.J.; Regn, D.; Bashratyan, R.; Dai, Y.D. Exosomes released by islet-derived mesenchymal stem cells trigger autoimmune responses in NOD mice. Diabetes 2014, 63, 1008–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Zhu, H.; Yang, S.; Fei, X. Let-7a-5p may participate in the pathogenesis of diabetic nephropathy through targeting HMGA2. Mol. Med. Rep. 2019, 19, 4229–4237. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Bae, Y.-U.; Jeon, J.S.; Noh, H.; Park, H.K.; Byun, D.W.; Han, D.C.; Ryu, S.; Kwon, S.H. The circulating exosomal microRNAs related to albuminuria in patients with diabetic nephropathy. J. Transl. Med. 2019, 17, 236. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Choi, M.E. Autophagy in diabetic nephropathy. J. Endocrinol. 2015, 224, R15. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Wang, W.W.; Zhang, M.Z.; Ma, Z.X.; Qiu, X.R.; Shen, M.; Yin, X.X. ROS induces epithelial-mesenchymal transition via the TGF-β1/PI3K/Akt/mTOR pathway in diabetic nephropathy. Exp. Ther. Med. 2019, 17, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Zahran, R.; Ghozy, A.; Elkholy, S.S.; El-Taweel, F.; El-Magd, M.A. Combination therapy with melatonin, stem cells and extracellular vesicles is effective in limiting renal ischemia–reperfusion injury in a rat model. Int. J. Urol. 2020, 27, 1039–1049. [Google Scholar] [CrossRef]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; Gazzar, W.B.E.; Sorour, S.M.; Seleem, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Ameliorated Diabetic Nephropathy by Autophagy Induction through the mTOR Signaling Pathway. Cells 2018, 7, 226. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, S.; Yamada, T.; Takahashi, K.; Munakata, Y.; Hosaka, S.; Takahashi, H.; Gao, J.; Shirai, Y.; Kodama, S.; Asai, Y. MicroRNAs 106b and 222 improve hyperglycemia in a mouse model of insulin-deficient diabetes via pancreatic β-cell proliferation. EBioMedicine 2017, 15, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Elsayed, G.R.; Salama, M.F.; El-Magd, M.A. Camel milk exosomes had a selective anticancer effect on PANC1 cells and a proliferative effect on H6c7 cells. Kafrelsheikh Vet. Med. J. 2022, 20, 1–5. [Google Scholar] [CrossRef]
- Wu, L.; Wang, L.; Liu, X.; Bai, Y.; Wu, R.; Li, X.; Mao, Y.; Zhang, L.; Zheng, Y.; Gong, T.; et al. Milk-derived exosomes exhibit versatile effects for improved oral drug delivery. Acta Pharm. Sin. B 2022, 12, 2029–2042. [Google Scholar] [CrossRef] [PubMed]
- Korish, A.A.; Abdel Gader, A.G.; Korashy, H.M.; Al-Drees, A.M.; Alhaider, A.A.; Arafah, M.M. Camel milk attenuates the biochemical and morphological features of diabetic nephropathy: Inhibition of Smad1 and collagen type IV synthesis. Chem.-Biol. Interact. 2015, 229, 100–108. [Google Scholar] [CrossRef]
- Badawy, A.A.; El-Magd, M.A.; AlSadrah, S.A. Therapeutic Effect of Camel Milk and Its Exosomes on MCF7 Cells In Vitro and In Vivo. Integr. Cancer Ther. 2018, 7, 1235–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-kattawy, A.M.; Algezawy, O.; Alfaifi, M.Y.; Noseer, E.A.; Hawsawi, Y.M.; Alzahrani, O.R.; Algarni, A.; Kahilo, K.A.; El-Magd, M.A. Therapeutic potential of camel milk exosomes against HepaRG cells with potent apoptotic, anti-inflammatory, and anti-angiogenesis effects for colostrum exosomes. Biomed. Pharmacother. 2021, 143, 112220. [Google Scholar] [CrossRef]
- Tesch, G.H.; Allen, T.J. Rodent models of streptozotocin-induced diabetic nephropathy (Methods in Renal Research). Nephrology 2007, 12, 261–266. [Google Scholar] [CrossRef]
- El-Magd, M.A.; Kahilo, K.A.; Nasr, N.E.; Kamal, T.; Shukry, M.; Saleh, A.A. A potential mechanism associated with lead-induced testicular toxicity in rats. Andrologia 2016, 49, e12750. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.A.; Ahmed, M.M.; El-Magd, M.A.; Magdy, A.; Ghamry, H.I.; Alshahrani, M.Y.; Abou El-Fotoh, M.F. Quercetin-Ameliorated, Multi-Walled Carbon Nanotubes-Induced Immunotoxic, Inflammatory, and Oxidative Effects in Mice. Molecules 2022, 27, 2117. [Google Scholar] [CrossRef]
- Zedan, A.M.G.; Sakran, M.I.; Bahattab, O.; Hawsawi, Y.M.; Al-Amer, O.; Oyouni, A.A.A.; Nasr Eldeen, S.K.; El-Magd, M.A. Oriental Hornet (Vespa orientalis) Larval Extracts Induce Antiproliferative, Antioxidant, Anti-Inflammatory, and Anti-Migratory Effects on MCF7 Cells. Molecules 2021, 26, 3303. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.L.; Sung, P.H.; Chen, K.H.; Shao, P.L.; Yang, C.C.; Cheng, B.C.; Lin, K.C.; Chen, C.H.; Chai, H.T.; Chang, H.W.; et al. Adipose-derived mesenchymal stem cell-derived exosomes alleviate overwhelming systemic inflammatory reaction and organ damage and improve outcome in rat sepsis syndrome. Am. J. Transl. Res. 2018, 10, 1053–1070. [Google Scholar]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Perkins, B.A.; Ficociello, L.H.; Ostrander, B.E.; Silva, K.H.; Weinberg, J.; Warram, J.H.; Krolewski, A.S. Microalbuminuria and the risk for early progressive renal function decline in type 1 diabetes. J. Am. Soc. Nephrol. 2007, 18, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.M.; Cordeiro, P.M.; Watanabe, M.; Fonseca, C.D.d.; Vattimo, M.d.F.F. The role of oxidative stress in streptozotocin-induced diabetic nephropathy in rats. Arch. Endocrinol. Metab. 2016, 60, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.-W.; Shen, C.-J.; Tung, Y.-T.; Chen, H.-L.; Chen, Y.-H.; Chang, W.-H.; Cheng, K.-C.; Yang, S.-H.; Chen, C.-M. Extracellular superoxide dismutase ameliorates streptozotocin-induced rat diabetic nephropathy via inhibiting the ROS/ERK1/2 signaling. Life Sci. 2015, 135, 77–86. [Google Scholar] [CrossRef]
- Mohamed, A.E.; El-Magd, M.A.; El-Said, K.S.; El-Sharnouby, M.; Tousson, E.M.; Salama, A.F. Potential therapeutic effect of thymoquinone and/or bee pollen on fluvastatin-induced hepatitis in rats. Sci. Rep. 2021, 11, 15688. [Google Scholar] [CrossRef]
- Elmoslemany, A.M.; El-Magd, M.A.; Ghamry, H.I.; Alshahrani, M.Y.; Zidan, N.S.; Zedan, A.M.G. Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats. Antioxidants 2021, 10, 1194. [Google Scholar] [CrossRef] [PubMed]
- Abu Gazia, M.; El-Magd, M.A. Effect of pristine and functionalized multiwalled carbon nanotubes on rat renal cortex. Acta Histochem. 2018, 121, 207–217. [Google Scholar] [CrossRef]
- Mohamed, Y.; Basyony, M.A.; El-Desouki, N.I.; Abdo, W.S.; El-Magd, M.A. The potential therapeutic effect for melatonin and mesenchymal stem cells on hepatocellular carcinoma. BioMedicine 2019, 9, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhady, D.; El-Abasy, M.; Abou-Asa, S.; Elbialy, Z.; Shukry, M.; Hussein, A.; Saleh, A.; El-Magd, M. The ameliorative effect of Aspergillus awamori on aflatoxin B1-induced hepatic damage in rabbits. World Mycotoxin J. 2017, 10, 363–373. [Google Scholar] [CrossRef]
- Navarro-Gonzalez, J.F.; Mora-Fernandez, C.; Muros de Fuentes, M.; Garcia-Perez, J. Inflammatory molecules and pathways in the pathogenesis of diabetic nephropathy. Nat. Rev. Nephrol. 2011, 7, 327–340. [Google Scholar] [CrossRef]
- Han, H.; Cao, A.; Wang, L.; Guo, H.; Zang, Y.; Li, Z.; Zhang, X.; Peng, W. Huangqi decoction ameliorates streptozotocin-induced rat diabetic nephropathy through antioxidant and regulation of the TGF-β/MAPK/PPAR-γ signaling. Cell. Physiol. Biochem. 2017, 42, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, A.; Kettritz, R.; Schreiber, A. Monocytes promote crescent formation in anti-myeloperoxidase antibody–induced glomerulonephritis. Am. J. Pathol. 2017, 187, 1908–1915. [Google Scholar] [CrossRef]
- Lin, J.; Glynn, R.J.; Rifai, N.; Manson, J.E.; Ridker, P.M.; Nathan, D.M.; Schaumberg, D.A. Inflammation and progressive nephropathy in type 1 diabetes in the diabetes control and complications trial. Diabetes Care 2008, 31, 2338–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liang, Y.; Huang, H.; Wang, L.; Li, Y.; Li, J.; Li, X.; Wang, H. ERK-dependent signaling pathway and transcriptional factor Ets-1 regulate matrix metalloproteinase-9 production in transforming growth factor-β1 stimulated glomerular podocytes. Cell. Physiol. Biochem. 2005, 16, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.; Naito, T.; Taguchi, T. Proto-oncogene Ets-1 and the kidney. Nephron 2001, 89, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-Y.; Miaw, C.-L.; Hsieh, C.-C.; Tseng, H.-C.; Yang, Y.-H.; Yen, C.-H. Effects of chronic 4-n-nonylphenol treatment on aortic vasoconstriction and vasorelaxation in rats. Arch. Toxicol. 2009, 83, 941–946. [Google Scholar] [CrossRef]
- Malik, A.; Mehmood, M.H.; Channa, H.; Akhtar, M.S.; Gilani, A.-H. Pharmacological basis for the medicinal use of polyherbal formulation and its ingredients in cardiovascular disorders using rodents. BMC Complement. Altern. Med. 2017, 17, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenoir, O.; Jasiek, M.; Hénique, C.; Guyonnet, L.; Hartleben, B.; Bork, T.; Chipont, A.; Flosseau, K.; Bensaada, I.; Schmitt, A. Endothelial cell and podocyte autophagy synergistically protect from diabetes-induced glomerulosclerosis. Autophagy 2015, 11, 1130–1145. [Google Scholar] [CrossRef] [Green Version]
- Tesch, G.H. Diabetic nephropathy–is this an immune disorder? Clin. Sci. 2017, 131, 2183–2199. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Isono, H.; Miyata, T. Potential antioxidant bioactive peptides from camel milk proteins. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2018, 4, 273–280. [Google Scholar] [CrossRef]
- Ebaid, H.; Ahmed, O.M.; Mahmoud, A.M.; Ahmed, R.R. Limiting prolonged inflammation during proliferation and remodeling phases of wound healing in streptozotocin-induced diabetic rats supplemented with camel undenatured whey protein. BMC Immunol. 2013, 14, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arab, H.H.; Salama, S.A.; Maghrabi, I.A. Camel Milk Ameliorates 5-Fluorouracil-Induced Renal Injury in Rats: Targeting MAPKs, NF-κB and PI3K/Akt/eNOS Pathways. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1628–1642. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Thakur, A.; Kar Li, W.; Qiu, G.; Yang, T.; He, B.; Lee, Y.; Lawrence Wu, C.-M. Site specific biotinylated antibody functionalized Ag@AuNIs LSPR biosensor for the ultrasensitive detection of exosomal MCT4, a glioblastoma progression biomarker. Chem. Eng. J. 2022, 446, 137383. [Google Scholar] [CrossRef]
- Thakur, A.; Qiu, G.; Xu, C.; Han, X.; Yang, T.; Ng, S.P.; Chan, K.W.Y.; Wu, C.M.L.; Lee, Y. Label-free sensing of exosomal MCT1 and CD147 for tracking metabolic reprogramming and malignant progression in glioma. Sci. Adv. 2020, 6, eaaz6119. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Xu, C.; Li, W.K.; Qiu, G.; He, B.; Ng, S.P.; Wu, C.L.; Lee, Y. In vivo liquid biopsy for glioblastoma malignancy by the AFM and LSPR based sensing of exosomal CD44 and CD133 in a mouse model. Biosens. Bioelectron. 2021, 191, 113476. [Google Scholar] [CrossRef]
- Thakur, A.; Mishra, A.P.; Panda, B.; Sweta, K.; Majhi, B. Detection of Disease-Specific Parent Cells Via Distinct Population of Nano-Vesicles by Machine Learning. Curr. Pharm. Des. 2020, 26, 3985–3996. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Thakur, A.; Xu, C.; Ng, S.-P.; Lee, Y.; Wu, C.-M.L. Detection of Glioma-Derived Exosomes with the Biotinylated Antibody-Functionalized Titanium Nitride Plasmonic Biosensor. Adv. Funct. Mater. 2019, 29, 1806761. [Google Scholar] [CrossRef]
- Thakur, A.; Qiu, G.; NG, S.-P.; Lawrence Wu, C.-M.; Lee, Y. Detection of membrane antigens of extracellular vesicles by surface plasmon resonance. J. Lab. Precis. Med. 2017, 2, 98. [Google Scholar] [CrossRef]
- Thakur, A.; Qiu, G.; Ng, S.P.; Guan, J.; Yue, J.; Lee, Y.; Wu, C.L. Direct detection of two different tumor-derived extracellular vesicles by SAM-AuNIs LSPR biosensor. Biosens. Bioelectron. 2017, 94, 400–407. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-----3′) | Reverse Primer (5′-----3′) |
|---|---|---|
| TGFβ1 | ACGTCAGACATTCGGGAAGCAGTG | GCAAGGACCTTGCTGTACTGTGTG |
| ICAM1 | GCAACCCCATCAAGAGGATTC | GGGGCCGTGTAGATAAACTCG |
| KIM1 | TGGCACTGTGACATCCTCAGA | GCAACGGACATGCCAACATA |
| ETS1 | CCTGCAGATTGTTCCGGAGT | CTGGGGCCAC CTTTACTGAG |
| ITGβ2 | GTTTCAGACAGAGGTCGGCA | AATTTCCTCCGGACAGGCAG |
| TIMP2 | GCAACCCCATCAAGAGGATTC | GGGGCCGTGTAGATAAACTCG |
| β-actin | AAGTCCCTCACCCTCCCAAAAG | AAGCAATGCTGTCACCTTCCC |
| Groups | Urea (mg/dL) | Creatinine (mg/dL) | Retinol Binding Protein (ng/mL) | Urinary Proteins (mg/mL) | Fasting Blood Glucose (mg/dL) |
|---|---|---|---|---|---|
| Cnt | 16.11 ± 0.85 e | 0.77 ± 0.04 d | 36.22 ± 1.49 e | 0.32 ± 0.01 d | 109.38 ± 5.66 d |
| DN | 72.32 ± 3.58 a | 2.10 ± 0.10 a | 215.54 ± 10.15 a | 1.90 ± 0.08 a | 348.60 ± 17.90 a |
| CM | 56.49 ± 2.17 b | 1.55 ± 0.08 b | 159.06 ± 7.42 b | 1.52 ± 0.06 b | 236.09 ± 12.00 b |
| EXO | 42.85 ± 2.03 c | 1.39 ± 0.07 b | 119.37 ± 5.38 c | 1.43 ± 0.06 b | 219.27 ± 11.62 b |
| CM + EXO | 34.27 ± 1.42 d | 1.04 ± 0.06 c | 88.73 ± 4.40 d | 0.95 ± 0.03 c | 177.46 ± 8.11 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaban, A.M.; Raslan, M.; Qahl, S.H.; Elsayed, K.; Abdelhameed, M.S.; Oyouni, A.A.A.; Al-Amer, O.M.; Hammouda, O.; El-Magd, M.A. Ameliorative Effects of Camel Milk and Its Exosomes on Diabetic Nephropathy in Rats. Membranes 2022, 12, 1060. https://doi.org/10.3390/membranes12111060
Shaban AM, Raslan M, Qahl SH, Elsayed K, Abdelhameed MS, Oyouni AAA, Al-Amer OM, Hammouda O, El-Magd MA. Ameliorative Effects of Camel Milk and Its Exosomes on Diabetic Nephropathy in Rats. Membranes. 2022; 12(11):1060. https://doi.org/10.3390/membranes12111060
Chicago/Turabian StyleShaban, Amira M., Mai Raslan, Safa H. Qahl, Khaled Elsayed, Mohamed Sayed Abdelhameed, Atif Abdulwahab A. Oyouni, Osama M. Al-Amer, Ola Hammouda, and Mohammed A. El-Magd. 2022. "Ameliorative Effects of Camel Milk and Its Exosomes on Diabetic Nephropathy in Rats" Membranes 12, no. 11: 1060. https://doi.org/10.3390/membranes12111060