
Staphylococcus aureus Carotenoids Modulate the Thermotropic Phase Behavior of Model Systems That Mimic Its Membrane Composition

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Bacterial Cultures
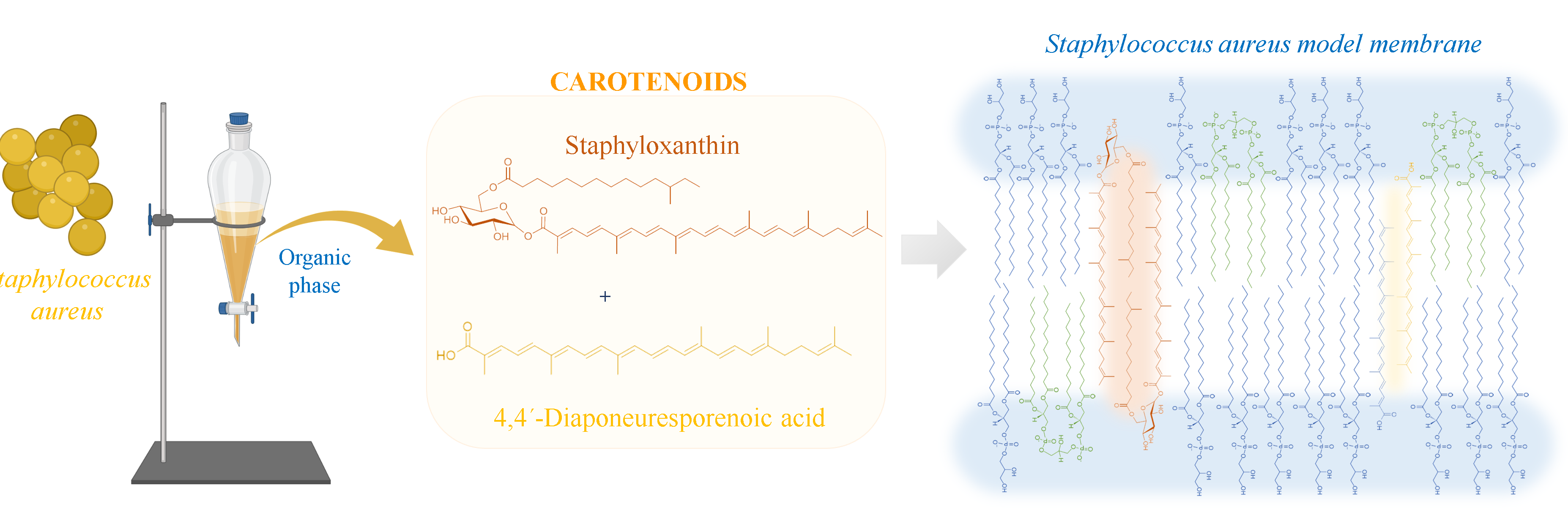
2.3. Carotenoid Extraction
2.4. Differential Scanning Calorimetry (DSC)
2.5. Infrared Spectroscopy Experiments
3. Results
3.1. Differential Scanning Calorimetry (DSC) of DMPG:CL Mixtures in the Presence of Carotenoids from S. aureus
3.2. Phase-Transition Measurements by Infrared Spectroscopy
4. Discussion
4.1. Effect of Carotenoid Content in Pure DMPG and CL Lipid Systems
4.2. Effect of Carotenoid Content in DMPG/CL Mixtures
4.3. Comparison with Live-Cell Systems and Other Model Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Vozza, E.G.; Mulcahy, M.E.; McLoughlin, R.M. Making the Most of the Host; Targeting the Autophagy Pathway Facilitates Staphylococcus aureus Intracellular Survival in Neutrophils. Front. Immunol. 2021, 12, 667387. [Google Scholar] [CrossRef] [PubMed]
- Braungardt, H.; Singh, V.K. Impact of deficiencies in branched-chain fatty acids and staphyloxanthin in Staphylococcus aureus. BioMed Res. Int. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus aureus Infection. Front. Immunol. 2021, 11, 620339. [Google Scholar] [CrossRef]
- Hernández-Villa, L.; Manrique-Moreno, M.; Leidy, C.; Jemioła-Rzemińska, M.; Ortíz, C.; Strzałka, K. Biophysical evaluation of cardiolipin content as a regulator of the membrane lytic effect of antimicrobial peptides. Biophys. Chem. 2018, 238, 8–15. [Google Scholar] [CrossRef]
- Romantsov, T.; Guan, Z.; Wood, J.M. Cardiolipin and the osmotic stress responses of bacteria. Biochim. Biophys. Acta 2009, 1788, 2092–2100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Muraih, J.K.; Tishbi, N.; Herskowitz, J.; Victor, R.L.; Silverman, J.; Uwumarenogie, S.; Taylor, S.D.; Palmer, M.; Mintzer, E. Cardiolipin prevents membrane translocation and permeabilization by daptomycin. J. Biol. Chem. 2014, 289, 11584–11591. [Google Scholar] [CrossRef] [PubMed]
- López, G.-D.; Álvarez-Rivera, G.; Carazzone, C.; Ibáñez, E.; Leidy, C.; Cifuentes, A. Bacterial Carotenoids: Extraction, Characterization, and Applications. Crit. Rev. Anal. Chem. 2021, 1–24. [Google Scholar]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef]
- Clauditz, A.; Resch, A.; Wieland, K.-P.; Peschel, A.; Götz, F. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef]
- Perez-Lopez, M.I.; Mendez-Reina, R.; Trier, S.; Herrfurth, C.; Feussner, I.; Bernal, A.; Forero-Shelton, M.; Leidy, C. Variations in carotenoid content and acyl chain composition in exponential, stationary and biofilm states of Staphylococcus aureus, and their influence on membrane biophysical properties. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2019, 1861, 978–987. [Google Scholar] [CrossRef] [PubMed]
- López, G.-D.; Suesca, E.; Álvarez-Rivera, G.; Rosato, A.E.; Ibáñez, E.; Cifuentes, A.; Leidy, C.; Carazzone, C. Carotenogenesis of Staphylococcus aureus: New insights and impact on membrane biophysical properties. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2021, 1866, 158941. [Google Scholar] [CrossRef] [PubMed]
- Coker, M.S.; Forbes, L.V.; Plowman-Holmes, M.; Murdoch, D.R.; Winterbourn, C.C.; Kettle, A.J. Interactions of staphyloxanthin and enterobactin with myeloperoxidase and reactive chlorine species. Arch. Biochem. Biophys. 2018, 646, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Chen, Y.Y.; Yan, Z.; Lu, W.; Wan, D.; Zhu, H. Staphyloxanthin: A potential target for antivirulence therapy. Infect. Drug Resist. 2019, 12, 2151. [Google Scholar] [CrossRef] [PubMed]
- Cueno, M.E.; Imai, K. Network analytics approach towards identifying potential antivirulence drug targets within the Staphylococcus aureus staphyloxanthin biosynthetic network. Arch. Biochem. Biophys. 2018, 645, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Crass, R.L.; Powell, K.L.; Huang, A.M. Daptomycin for the treatment of Staphylococcus aureus infections complicated by septic pulmonary emboli. Diagn. Microbiol. Infect. Dis. 2019, 93, 131–135. [Google Scholar] [CrossRef]
- Dhand, A.; Sakoulas, G. Daptomycin in combination with other antibiotics for the treatment of complicated methicillin-resistant Staphylococcus aureus bacteremia. Clin. Ther. 2014, 36, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Heidary, M.; Khosravi, A.D.; Khoshnood, S.; Nasiri, M.J.; Soleimani, S.; Goudarzi, M. Daptomycin. J. Antimicrob. Chemother. 2018, 73, 1–11. [Google Scholar] [CrossRef]
- Chamberlain, N.R.; Mehrtens, B.; Xiong, Z.; Kapral, F.; Boardman, J.; Rearick, J. Correlation of carotenoid production, decreased membrane fluidity, and resistance to oleic acid killing in Staphylococcus aureus 18Z. Infect. Immun. 1991, 59, 4332–4337. [Google Scholar] [CrossRef]
- Mishra, N.N.; Liu, G.Y.; Yeaman, M.R.; Nast, C.C.; Proctor, R.A.; McKinnell, J.; Bayer, A.S. Carotenoid-related alteration of cell membrane fluidity impacts Staphylococcus aureus susceptibility to host defense peptides. Antimicrob. Agents Chemother. 2011, 55, 526–531. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, E.; Koch, G.; Wagner, R.M.; Fekete, A.; Stengel, S.T.; Schneider, J.; Mielich-Süss, B.; Geibel, S.; Markert, S.M.; Stigloher, C. Membrane microdomain disassembly inhibits MRSA antibiotic resistance. Cell 2017, 171, 1354–1367.e20. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, G.; Sagarzazu, N.; Pagán, R.; Condón, S.; Mañas, P. Heat and pulsed electric field resistance of pigmented and non-pigmented enterotoxigenic strains of Staphylococcus aureus in exponential and stationary phase of growth. Int. J. Food Microbiol. 2007, 118, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.I.; Trier, S.M.; Bernal, A.; Vargas, J.C.; Herrfurth, C.; Feussner, I.; Gonzalez, J.M.; Leidy, C.S. Aureus Adapt to Growth Conditions by Changing Membrane Order. Biophys. J. 2014, 106 (Suppl. S1), 580a. [Google Scholar] [CrossRef]
- White, D.C.; Frerman, F.E. Extraction, characterization, and cellular localization of the lipids of Staphylococcus aureus. J. Bacteriol. 1967, 94, 1854–1867. [Google Scholar] [CrossRef]
- Tristram-Nagle, S.; Liu, Y.; Legleiter, J.; Nagle, J.F. Structure of Gel Phase DMPC Determined by X-Ray Diffraction. Biophys. J. 2002, 83, 3324–3335. [Google Scholar] [CrossRef]
- Lewis, R.N.; Zweytick, D.; Pabst, G.; Lohner, K.; McElhaney, R.N. Calorimetric, X-ray diffraction, and spectroscopic studies of the thermotropic phase behavior and organization of tetramyristoyl cardiolipin membranes. Biophys. J. 2007, 92, 3166–3177. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.N.; McElhaney, R.N. Membrane lipid phase transitions and phase organization studied by Fourier transform infrared spectroscopy. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2013, 1828, 2347–2358. [Google Scholar] [CrossRef]
- Mantsch, H.H.; McElhaney, R.N. Phospholipid phase transitions in model and biological membranes as studied by infrared spectroscopy. Chem. Phys. Lipids 1991, 57, 213–226. [Google Scholar] [CrossRef]
- Mantsch, H.H.; McElhaney, R.N. Applications of infrared spectroscopy to biology and medicine. J. Mol. Struct. 1990, 217, 347–362. [Google Scholar] [CrossRef]
- Gauger, D.R.; Selle, C.; Fritzsche, H.; Pohle, W. Chain-length dependence of the hydration properties of saturated phosphatidylcholines as revealed by FTIR spectroscopy. J. Mol. Struct. 2001, 565–566, 25–29. [Google Scholar] [CrossRef]
- Domingo, J.C.; Mora, M.; Africa de Madariaga, M. Role of headgroup structure in the phase behaviour of N-acylethanolamine phospholipids: Hydrogen-bonding ability and headgroup size. Chem. Phys. Lipids 1994, 69, 229–240. [Google Scholar] [CrossRef]
- Marshall, J.H.; Wilmoth, G.J. Pigments of Staphylococcus aureus, a series of triterpenoid carotenoids. J. Bacteriol. 1981, 147, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Pelz, A.; Wieland, K.-P.; Putzbach, K.; Hentschel, P.; Albert, K.; Götz, F. Structure and Biosynthesis of Staphyloxanthin from Staphylococcus aureus*. J. Biol. Chem. 2005, 280, 32493–32498. [Google Scholar] [CrossRef] [PubMed]
- Eliasz, A.W.; Chapman, D.; Ewing, D.F. Phospholipid phase transitions. Effects of n-alcohols, n-monocarboxylic acids, phenylalkyl alcohols and quatenary ammonium compounds. Biochim. Biophys. Acta 1976, 448, 220–233. [Google Scholar] [CrossRef]
- Fa, N.; Ronkart, S.; Schanck, A.; Deleu, M.; Gaigneaux, A.; Goormaghtigh, E.; Mingeot-Leclercq, M.P. Effect of the antibiotic azithromycin on thermotropic behavior of DOPC or DPPC bilayers. Chem. Phys. Lipids 2006, 144, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Yellin, N.; Levin, I.W. Cooperative unit size in the gel-liquid crytalline phase transition of dipalmitoyl phosphatidylcholine-water multilayers: An estimate from raman spectroscopy. Biochim. Biophys. Acta 1977, 468, 490–494. [Google Scholar] [CrossRef]
- Tiwari, K.B.; Gatto, C.; Wilkinson, B.J. Interrelationships among Fatty Acid Composition, Staphyloxanthin Content, Fluidity, and Carbon Flow in the Staphylococcus aureus Membrane. Molecules 2018, 23, 1201. [Google Scholar] [CrossRef] [PubMed]
- Valliammai, A.; Selvaraj, A.; Muthuramalingam, P.; Priya, A.; Ramesh, M.; Pandian, S.K. Staphyloxanthin inhibitory potential of thymol impairs antioxidant fitness, enhances neutrophil mediated killing and alters membrane fluidity of methicillin resistant Staphylococcus aureus. Biomed. Pharm. 2021, 141, 111933. [Google Scholar] [CrossRef]
- Kossakowska-Zwierucho, M.; Kaźmierkiewicz, R.; Bielawski, K.P.; Nakonieczna, J. Factors determining Staphylococcus aureus susceptibility to photoantimicrobial chemotherapy: RsbU activity, staphyloxanthin level, and membrane fluidity. Front. Microbiol. 2016, 7, 1141. [Google Scholar] [CrossRef] [PubMed]
- Seel, W.; Baust, D.; Sons, D.; Albers, M.; Etzbach, L.; Fuss, J.; Lipski, A. Carotenoids are used as regulators for membrane fluidity by Staphylococcus xylosus. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McMullen, T.P.W.; McElhaney, R.N. New aspects of the interaction of cholesterol with dipalmitoylphosphatidylcholine bilayers as revealed by high-sensitivity differential scanning calorimetry. Biochim. Biophys. Acta 1995, 1234, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Mills, T.T.; Huang, J.; Feigenson, G.W.; Nagle, J.F. Effects of cholesterol and unsaturated DOPC lipid on chain packing of saturated gel-phase DPPC bilayers. Gen. Physiol. Biophys. 2009, 28, 126. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.; Schuster, H.; Brauner, J.; Mendelsohn, R. Effects of cholesterol on conformational disorder in dipalmitoylphosphatidylcholine bilayers. A quantitative IR study of the depth dependence. Biochemistry 1990, 29, 4368–4373. [Google Scholar] [CrossRef] [PubMed]
- Drobnis, E.Z.; Crowe, L.M.; Berger, T.; Anchordoguy, T.J.; Overstreet, J.W.; Crowe, J.H. Cold shock damage is due to lipid phase transitions in cell membranes: A demonstration using sperm as a model. J. Exp. Zool. 1993, 265, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, P.S.; Cronan Jr, J.E.; De Mendoza, D. A Bacillus subtilis gene induced by cold shock encodes a membrane phospholipid desaturase. J. Bacteriol. 1998, 180, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.H.; Marahiel, M.A. Coping with the cold: The cold shock response in the Gram-positive soil bacterium Bacillus subtilis. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 357, 895–907. [Google Scholar] [CrossRef]
- Beranová, J.; Jemioła-Rzemińska, M.; Elhottová, D.; Strzałka, K.; Konopásek, I. Metabolic control of the membrane fluidity in Bacillus subtilis during cold adaptation. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2008, 1778, 445–453. [Google Scholar] [CrossRef]
- Mileykovskaya, E.; Dowhan, W. Cardiolipin membrane domains in prokaryotes and eukaryotes. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2009, 1788, 2084–2091. [Google Scholar] [CrossRef]
- Koprivnjak, T.; Zhang, D.; Ernst, C.; Peschel, A.; Nauseef, W.; Weiss, J. Characterization of Staphylococcus aureus cardiolipin synthases 1 and 2 and their contribution to accumulation of cardiolipin in stationary phase and within phagocytes. J. Bacteriol. 2011, 193, 4134–4142. [Google Scholar] [CrossRef]
- Kumar, S.V.; Taylor, G.; Hasim, S.; Collier, C.P.; Farmer, A.T.; Campagna, S.R.; Bible, A.N.; Doktycz, M.J.; Morrell-Falvey, J. Loss of carotenoids from membranes of Pantoea sp. YR343 results in altered lipid composition and changes in membrane biophysical properties. Biochim. Biophys. Acta 2019, 1861, 1338–1345. [Google Scholar] [CrossRef]
- Silva, T.R.e.; Silva, L.C.F.; de Queiroz, A.C.; Alexandre Moreira, M.S.; de Carvalho Fraga, C.A.; de Menezes, G.C.A.; Rosa, L.H.; Bicas, J.; de Oliveira, V.M.; Duarte, A.W.F. Pigments from Antarctic bacteria and their biotechnological applications. Crit. Rev. Biotechnol. 2021, 41, 809–826. [Google Scholar] [CrossRef] [PubMed]
- Gabrielska, J.; Gruszecki, W.I. Zeaxanthin (dihydroxy-β-carotene) but not β-carotene rigidifies lipid membranes: A 1H-NMR study of carotenoid-egg phosphatidylcholine liposomes. Biochim. Biophys. Acta 1996, 1285, 167–174. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Markowska, E.; Gruszecki, W.I.; Sielewiesiuk, J. Effects of polar carotenoids on dimyristoylphosphatidylcholine membranes: A spin-label study. Biochim. Biophys. Acta 1992, 1105, 97–108. [Google Scholar] [CrossRef]
- Augustynska, D.; Jemioła-Rzemińska, M.; Burda, K.; Strzałka, K. Influence of polar and nonpolar carotenoids on structural and adhesive properties of model membranes. Chem. Biol. Interact. 2015, 239, 19–25. [Google Scholar] [CrossRef]
- Gruszecki, W.I.; Strzałka, K. Carotenoids as modulators of lipid membrane physical properties. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2005, 1740, 108–115. [Google Scholar] [CrossRef]
- Kostecka-Gugała, A.; Latowski, D.; Strzałka, K. Thermotropic phase behaviour of α-dipalmitoylphosphatidylcholine multibilayers is influenced to various extents by carotenoids containing different structural features- evidence from differential scanning calorimetry. Biochim. Biophys. Acta 2003, 1609, 193–202. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak #1 | Peak #2 | |||||||
|---|---|---|---|---|---|---|---|---|
| T [°C] | ΔT1/2 [°C] | Cpmax [kJ mol−1 K−1] | ΔH [kJ mol−1] | T [°C] | ΔT1/2 [°C] | Cpmax [kJ mol−1 K−1] | ΔH [kJ mol−1] | |
| DMPG:CL | 28.10 | 3.33 | 4.41 | 16.88 | 31.27 | 2.38 | 4.00 | 11.12 |
| +1 mol% Car | 28.05 | 3.81 | 4.18 | 18.02 | 31.36 | 2.48 | 4.05 | 10.94 |
| +3 mol% Car | 27.61 | 4.38 | 3.95 | 19.45 | 31.10 | 2.76 | 3.77 | 11.15 |
| +5 mol% Car | 27.55 | 5.14 | 3.41 | 18.25 | 31.18 | 2.76 | 3.36 | 10.12 |
| +10 mol% Car | 27.18 | 5.71 | 3.05 | 18.85 | 31.13 | 2.86 | 3.23 | 10.98 |
| +20 mol% Car | 26.06 | 6.29 | 2.27 | 18.12 | 30.50 | 3.24 | 2.82 | 10.61 |
| Lipids System/ Carotenoids (mol%) | DMPG | CL | DMPG:CL |
|---|---|---|---|
| 0 | 22.9 | 43.4 | 28.7 |
| 1 | 22.6 | 42.0 | 30.1 |
| 3 | 22.5 | 41.1 | 29.3 |
| 5 | 22.4 | 40.9 | 29.4 |
| 10 | 21.7 | 40.9 | 28.3 |
| 20 | 21.7 | 39.4 | 28.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manrique-Moreno, M.; Jemioła-Rzemińska, M.; Múnera-Jaramillo, J.; López, G.-D.; Suesca, E.; Leidy, C.; Strzałka, K. Staphylococcus aureus Carotenoids Modulate the Thermotropic Phase Behavior of Model Systems That Mimic Its Membrane Composition. Membranes 2022, 12, 945. https://doi.org/10.3390/membranes12100945
Manrique-Moreno M, Jemioła-Rzemińska M, Múnera-Jaramillo J, López G-D, Suesca E, Leidy C, Strzałka K. Staphylococcus aureus Carotenoids Modulate the Thermotropic Phase Behavior of Model Systems That Mimic Its Membrane Composition. Membranes. 2022; 12(10):945. https://doi.org/10.3390/membranes12100945
Chicago/Turabian StyleManrique-Moreno, Marcela, Małgorzata Jemioła-Rzemińska, Jessica Múnera-Jaramillo, Gerson-Dirceu López, Elizabeth Suesca, Chad Leidy, and Kazimierz Strzałka. 2022. "Staphylococcus aureus Carotenoids Modulate the Thermotropic Phase Behavior of Model Systems That Mimic Its Membrane Composition" Membranes 12, no. 10: 945. https://doi.org/10.3390/membranes12100945