

Expression of Mutant Glycine Receptors in Xenopus Oocytes Using Canonical and Non-Canonical Amino Acids Reveals Distinct Roles of Conserved Proline Residues

Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Biology
2.2. Oocyte Preparation and RNA Injection
2.3. Electrophysiology
2.4. Structures
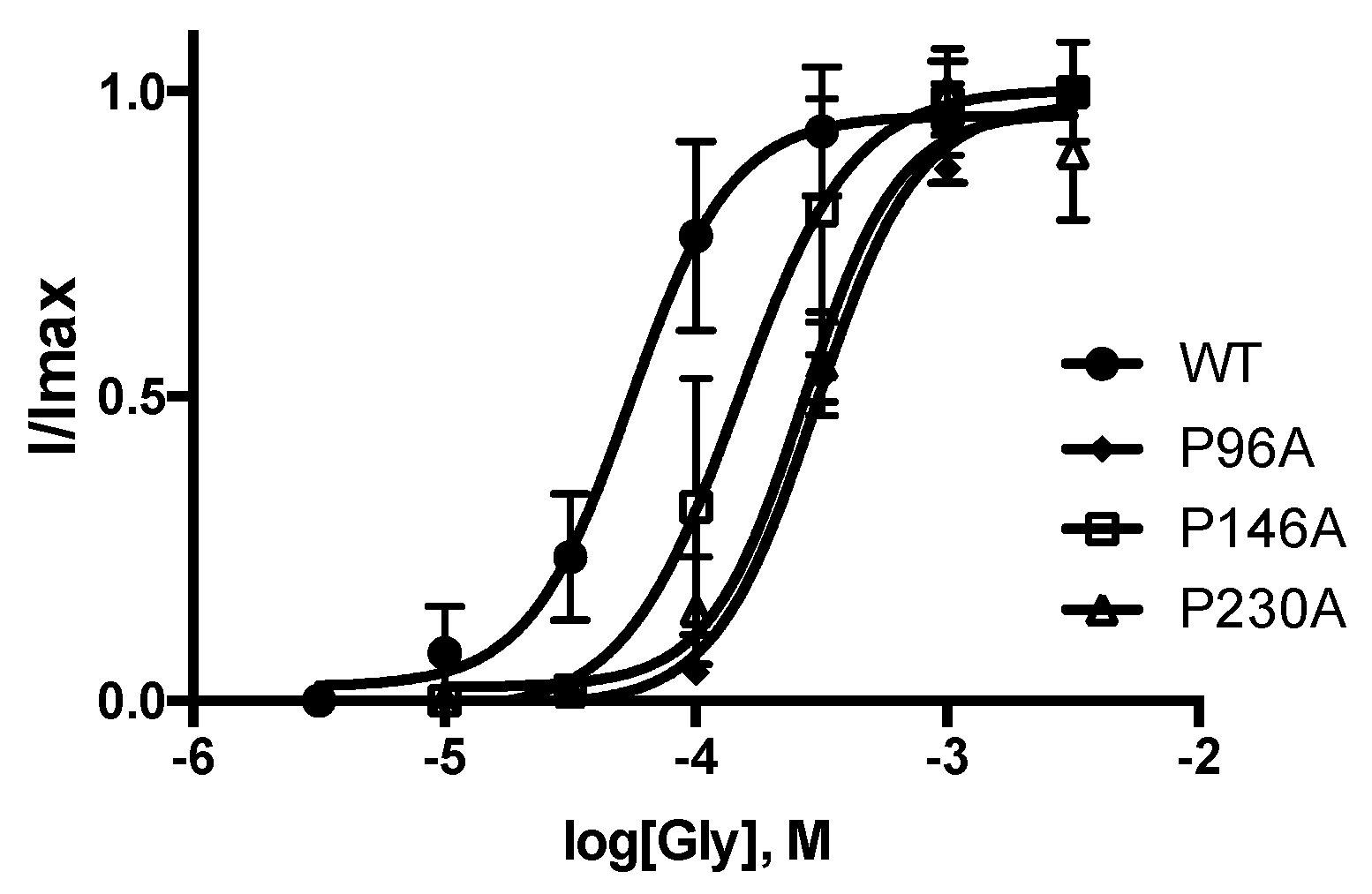
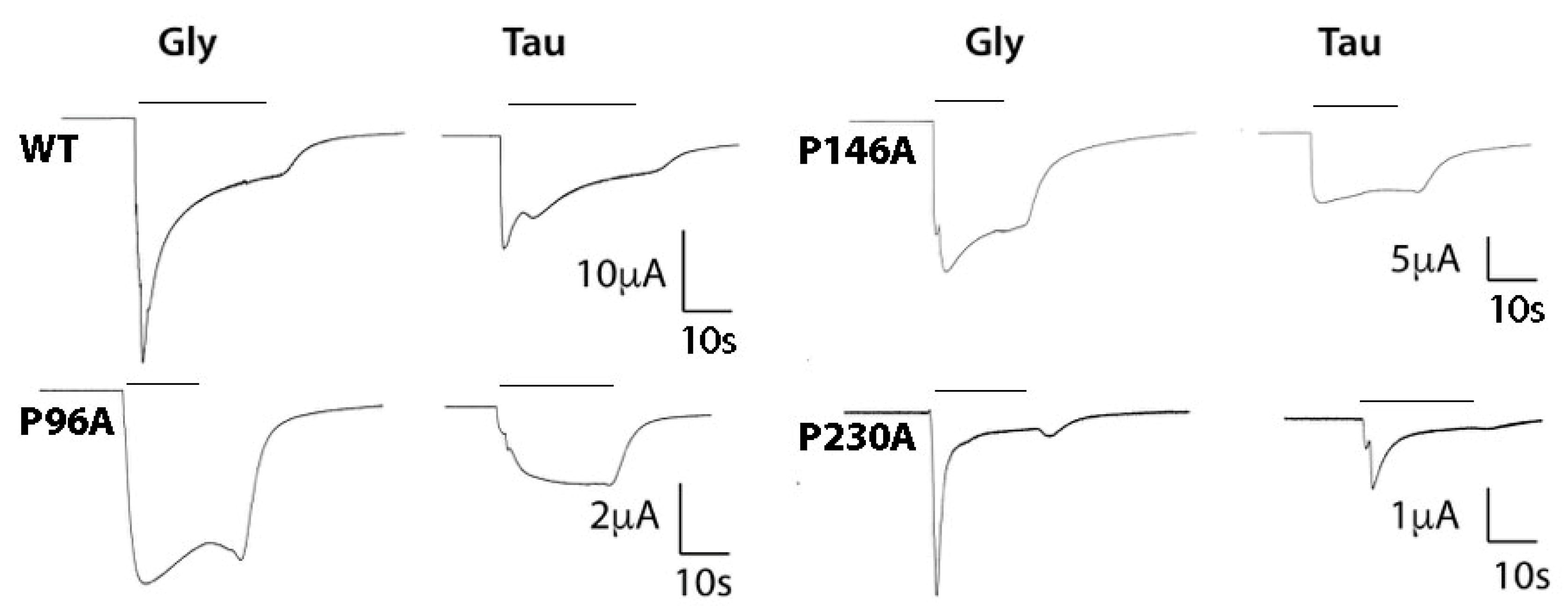
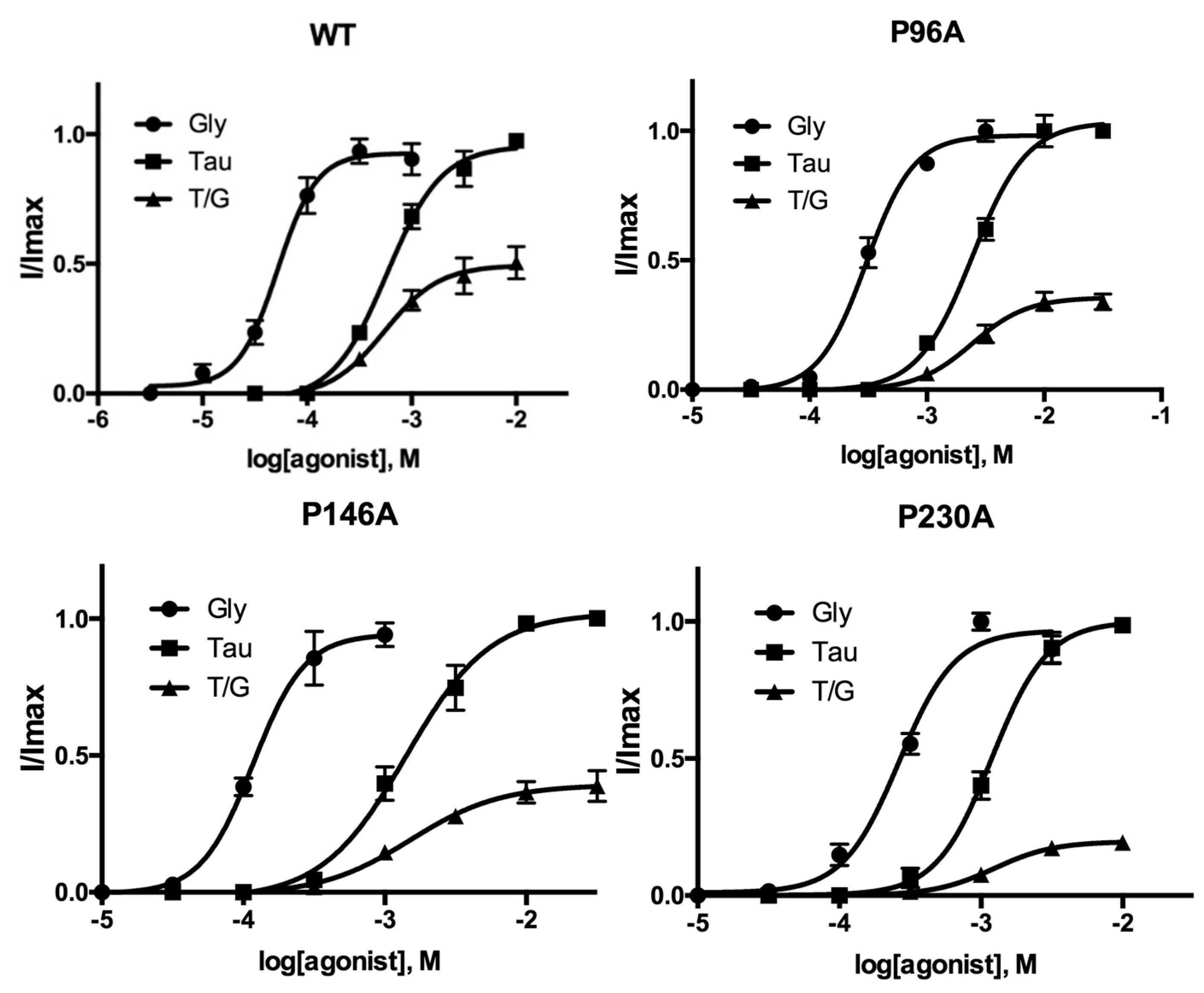
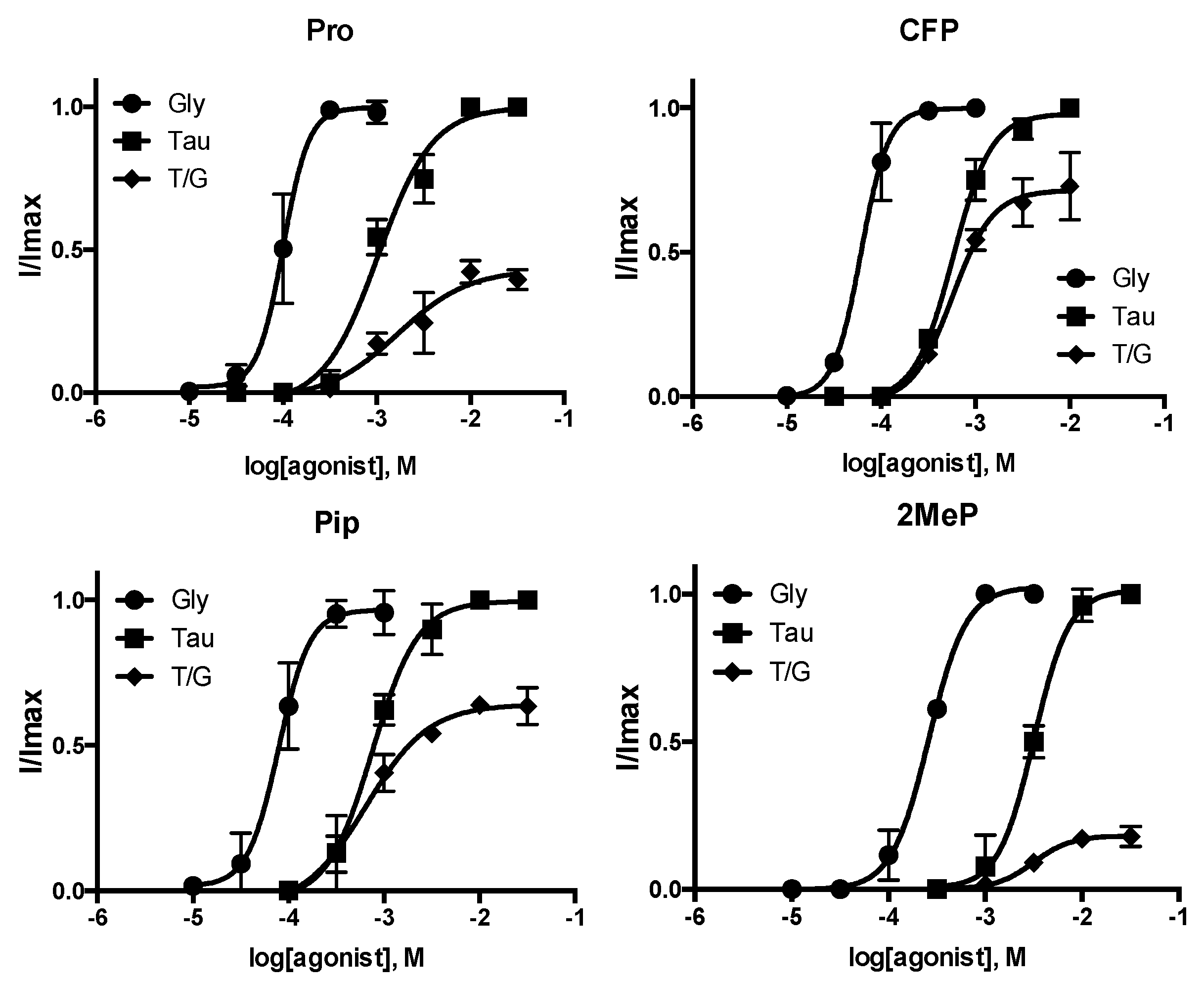
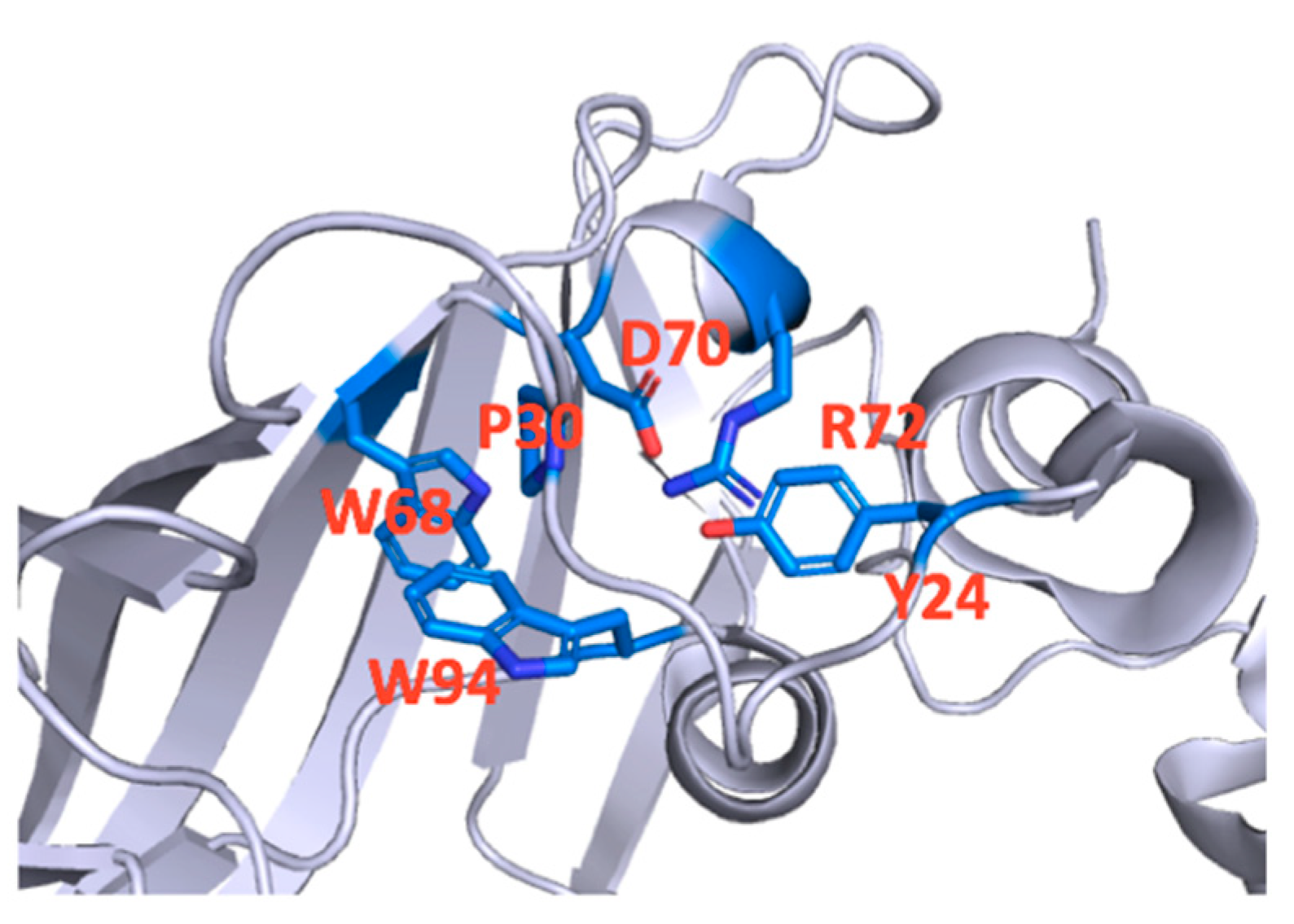
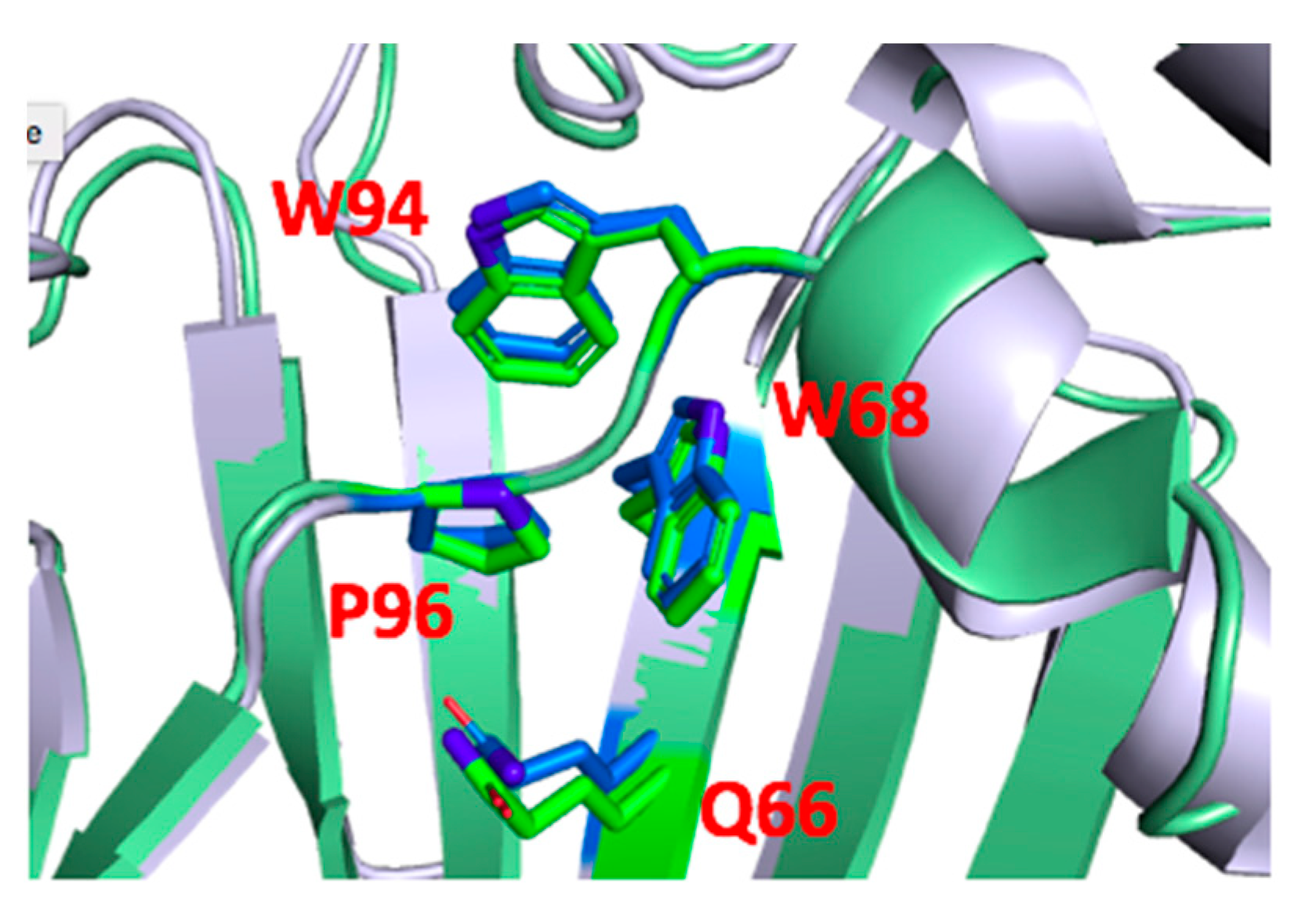
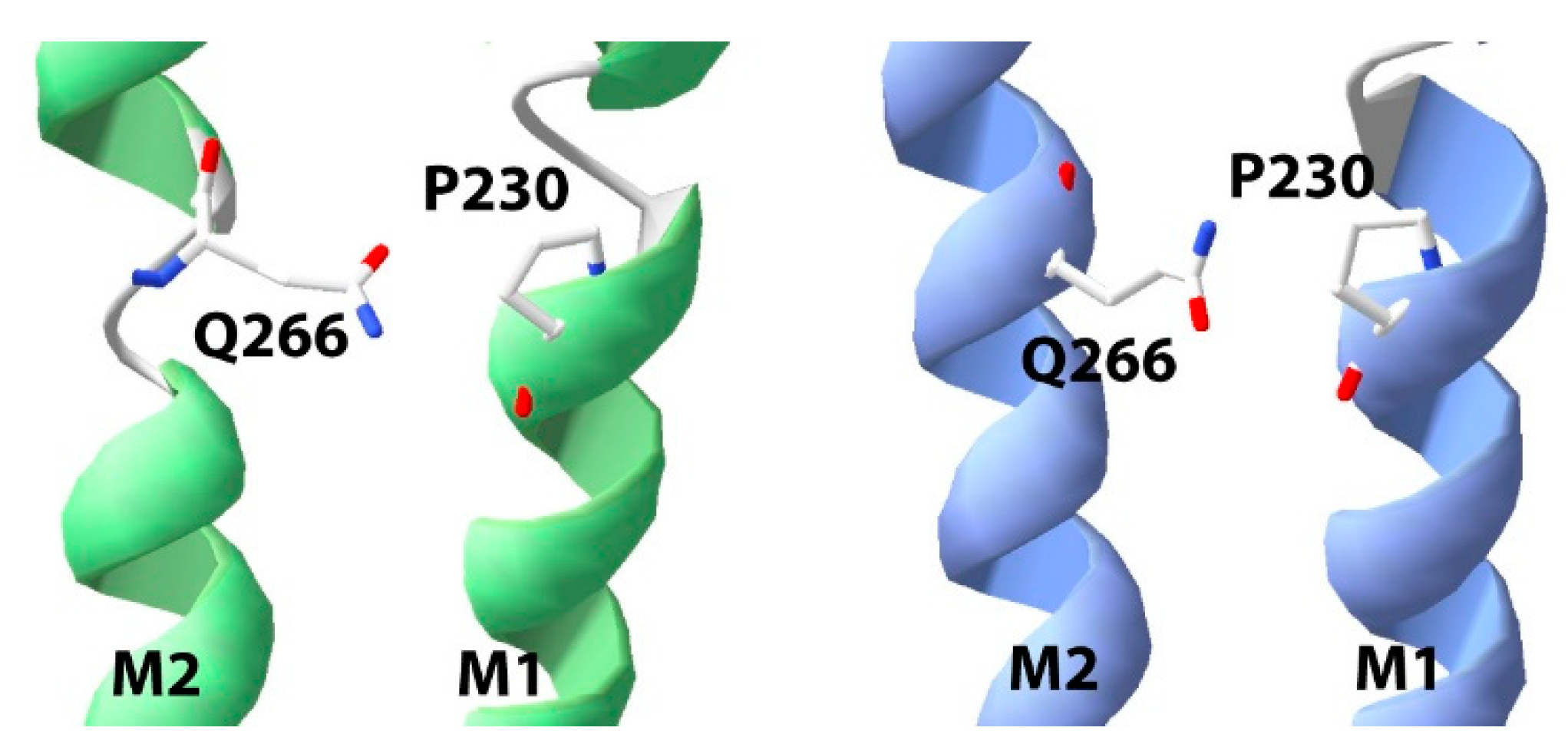
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burgos, C.F.; Yevenes, G.E.; Aguayo, L.G. Structure and Pharmacologic Modulation of Inhibitory Glycine Receptors. Mol. Pharmacol. 2016, 90, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.W. Molecular structure and function of the glycine receptor chloride channel. Physiol. Rev. 2004, 84, 1051–1095. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.W. Native glycine receptor subtypes and their physiological roles. Neuropharmacology 2009, 56, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [Green Version]
- Bocquet, N.; Nury, H.; Baaden, M.; Le Poupon, C.; Changeux, J.P.; Delarue, M.; Corringer, P.J. X-ray structure of a pentameric ligand-gated ion channel in an apparently open conformation. Nature 2009, 457, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Hassaine, G.; Deluz, C.; Grasso, L.; Wyss, R.; Tol, M.B.; Hovius, R.; Graff, A.; Stahlberg, H.; Tomizaki, T.; Desmyter, A.; et al. X-ray structure of the mouse serotonin 5-HT3 receptor. Nature 2014, 512, 276–281. [Google Scholar] [CrossRef]
- Hibbs, R.E.; Gouaux, E. Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature 2011, 474, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Hilf, R.J.; Dutzler, R. X-ray structure of a prokaryotic pentameric ligand-gated ion channel. Nature 2008, 452, 375–379. [Google Scholar] [CrossRef]
- Hilf, R.J.; Dutzler, R. Structure of a potentially open state of a proton-activated pentameric ligand-gated ion channel. Nature 2009, 457, 115–118. [Google Scholar] [CrossRef]
- Huang, X.; Chen, H.; Michelsen, K.; Schneider, S.; Shaffer, P.L. Crystal structure of human glycine receptor-alpha3 bound to antagonist strychnine. Nature 2015, 526, 277–280. [Google Scholar] [CrossRef]
- Huang, X.; Shaffer, P.L.; Ayube, S.; Bregman, H.; Chen, H.; Lehto, S.G.; Luther, J.A.; Matson, D.J.; McDonough, S.I.; Michelsen, K.; et al. Crystal structures of human glycine receptor alpha3 bound to a novel class of analgesic potentiators. Nat. Struct. Mol. Biol. 2017, 24, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Overington, J.P. Derivation of rules for comparative protein modeling from a database of protein structure alignments. Protein Sci. 1994, 3, 1582–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiteh, M.; Taly, A.; Henin, J. Evolution of Pentameric Ligand-Gated Ion Channels: Pro-Loop Receptors. PLoS ONE 2016, 11, e0151934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, M.W.; Gallivan, J.P.; Silverman, S.K.; Labarca, C.G.; Dougherty, D.A.; Lester, H.A. In vivo incorporation of unnatural amino acids into ion channels in Xenopus oocyte expression system. Methods Enzymol. 1998, 293, 504–529. [Google Scholar] [PubMed]
- Reeves, D.C.; Goren, E.N.; Akabas, M.H.; Lummis, S.C. Structural and electrostatic properties of the 5-HT3 receptor pore revealed by substituted cysteine accessibility mutagenesis. J. Biol. Chem. 2001, 276, 42035–42042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Saint Jan, D.; David-Watine, B.; Korn, H.; Bregestovski, P. Activation of human alpha1 and alpha2 homomeric glycine receptors by taurine and GABA. J. Physiol. 2001, 535 Pt 3, 741–755. [Google Scholar]
- Baker, B.W.; Dougherty, D.A.; Lummis, S.C.R. Proline Residues Contribute to Efficient GABAp Receptor Function. ACS Chem. Neurosci. 2020, 11, 4215–4222. [Google Scholar] [CrossRef]
- Deane, C.M.; Lummis, S.C. The role and predicted propensity of conserved proline residues in the 5-HT3 receptor. J. Biol. Chem. 2001, 276, 37962–37966. [Google Scholar] [CrossRef]
- Mosesso, R.; Dougherty, D.A.; Lummis, S.C.R. Probing Proline Residues in the Prokaryotic Ligand-Gated Ion Channel, ELIC. Biochemistry 2018, 57, 4036–4043. [Google Scholar] [CrossRef] [Green Version]
- Rienzo, M.; Rocchi, A.R.; Threatt, S.D.; Dougherty, D.A.; Lummis, S.C. Perturbation of Critical Prolines in Gloeobacter violaceus Ligand-gated Ion Channel (GLIC) Supports Conserved Gating Motions among Cys-loop Receptors. J. Biol. Chem. 2016, 291, 6272–6280. [Google Scholar] [CrossRef] [Green Version]
- Braun, N.; Lynagh, T.; Yu, R.; Biggin, P.C.; Pless, S.A. Role of an Absolutely Conserved Tryptophan Pair in the Extracellular Domain of Cys-Loop Receptors. ACS Chem. Neurosci. 2016, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, N.; Kluck, C.J.; Price, K.L.; Meiselbach, H.; Vornberger, N.; Schwarzinger, S.; Hartmann, S.; Langlhofer, G.; Schulz, S.; Schlegel, N.; et al. Disturbed neuronal ER-Golgi sorting of unassembled glycine receptors suggests altered subcellular processing is a cause of human hyperekplexia. J. Neurosci. 2015, 35, 422–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Devenish, S.O.; Lummis, S.C.R. Identification of Novel Functionally Important Aromatic Residue Interactions in the Extracellular Domain of the Glycine Receptor. Biochemistry 2018, 57, 4029–4035. [Google Scholar] [CrossRef] [PubMed]
- Limapichat, W.; Lester, H.A.; Dougherty, D.A. Chemical scale studies of the Phe-Pro conserved motif in the cys loop of Cys loop receptors. J. Biol. Chem. 2010, 285, 8976–8984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Naduthambi, D.; Thomas, K.M.; Zondlo, N.J. Proline editing: A general and practical approach to the synthesis of functionally and structurally diverse peptides. Analysis of steric versus stereoelectronic effects of 4-substituted prolines on conformation within peptides. J. Am. Chem. Soc. 2013, 135, 4333–4363. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.K.; Park, H.S. Conformational preferences of the 2-methylproline residue and its role in stabilizing β-turn and polyproline II structures of peptides. New J. Chem. 2014, 38, 2831–2840. [Google Scholar] [CrossRef]
- Jabs, A.; Weiss, M.S.; Hilgenfeld, R. Non-proline cis peptide bonds in proteins. J. Mol. Biol. 1999, 286, 291–304. [Google Scholar] [CrossRef]
- Valley, C.C.; Cembran, A.; Perlmutter, J.D.; Lewis, A.K.; Labello, N.P.; Gao, J.; Sachs, J.N. The methionine-aromatic motif plays a unique role in stabilizing protein structure. J. Biol. Chem. 2012, 287, 34979–34991. [Google Scholar] [CrossRef] [Green Version]
- Viguera, A.R.; Serrano, L. Stable proline box motif at the N-terminal end of alpha-helices. Protein Sci. 1999, 8, 1733–1742. [Google Scholar] [CrossRef]
- Dang, H.; England, P.M.; Farivar, S.S.; Dougherty, D.A.; Lester, H.A. Probing the role of a conserved M1 proline residue in 5-hydroxytryptamine(3) receptor gating. Mol. Pharmacol. 2000, 57, 1114–1122. [Google Scholar]
- England, P.M.; Zhang, Y.; Dougherty, D.A.; Lester, H.A. Backbone mutations in transmembrane domains of a ligand-gated ion channel: Implications for the mechanism of gating. Cell 1999, 96, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.E.; Mesoy, S.M.; Capes, E.; Lummis, S.C.R. Many Proline Residues in the Extracellular Domain Contribute to Glycine Receptor Function. ACS Chem. Neurosci. 2020, 11, 2658–2665. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.; Wood, S.E.; Mullins, J.G.; Keramidas, A.; Cushion, T.D.; Thomas, R.H.; Pickrell, W.O.; Drew, C.J.; Masri, A.; Jones, E.A.; et al. New hyperekplexia mutations provide insight into glycine receptor assembly, trafficking, and activation mechanisms. J. Biol. Chem. 2013, 288, 33745–33759. [Google Scholar] [CrossRef] [Green Version]
- Scott, S.; Lynch, J.W.; Keramidas, A. Correlating structural and energetic changes in glycine receptor activation. J. Biol. Chem. 2015, 290, 5621–5634. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gly | Tau | |||||||
|---|---|---|---|---|---|---|---|---|
| pEC50, M | EC50 μM | nH | pEC50, M | EC50 μM | nH | % ImaxTau /ImaxGly | n | |
| WT | 4.282 ± 0.05 | 52 | 2.3 ± 0.5 | 3.235 ± 0.05 | 583 | 1.7 ± 0.3 | 50 ± 4 | 5 |
| P96A | 3.518 ± 0.03 * | 304 | 2.2 ± 0.3 | 2.612 ± 0.03 * | 2440 | 1.9 ± 0.2 | 36 ± 2 * | 5 |
| P146A | 3.932 ± 0.05 * | 117 | 2.4 ± 0.7 | 2.850 ± 0.06 * | 1410 | 1.5 ± 0.3 | 39 ± 3 * | 5 |
| P230A | 3.579 ± 0.03 * | 264 | 2.2 ± 0.4 | 2.925 ± 0.03 * | 1190 | 2.2 ± 0.4 | 20 ± 1 * | 5 |
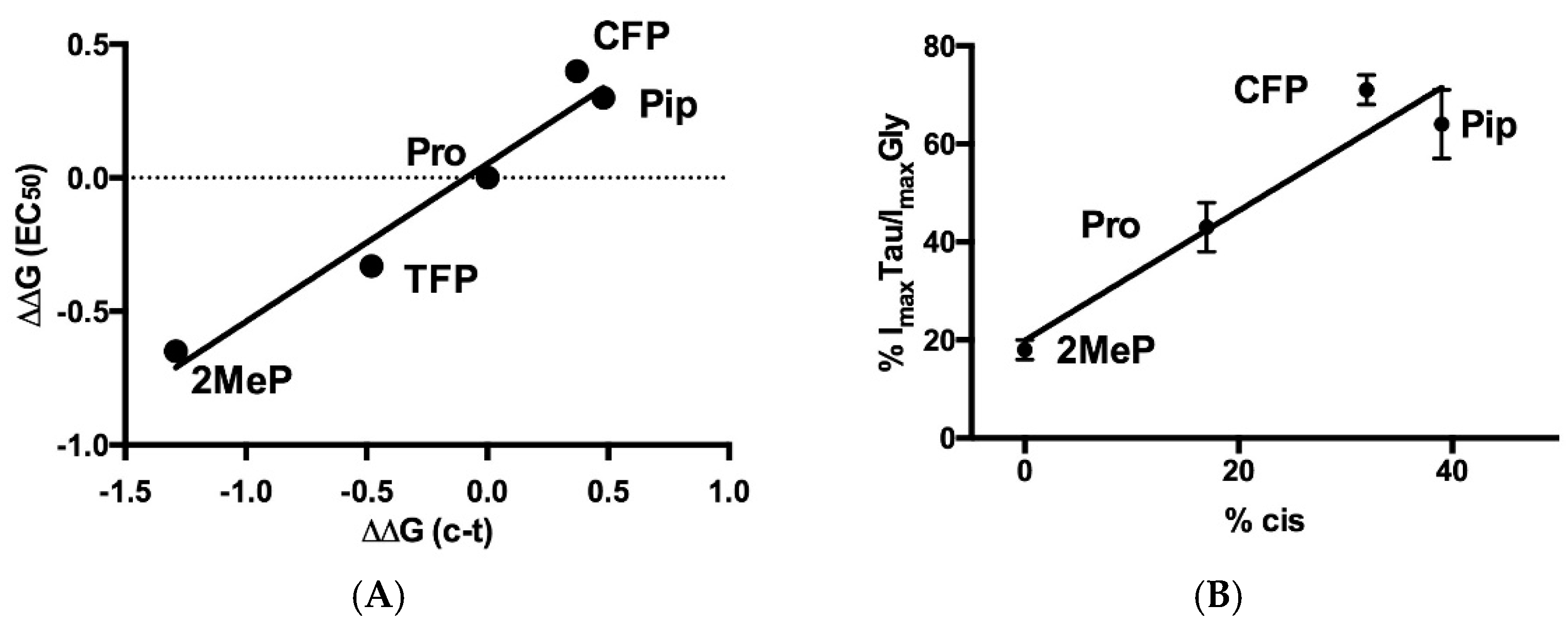
| Site | Substitution | pEC50, M | EC50, μM | nH | n |
|---|---|---|---|---|---|
| P30(α1β1L) | Pro | 3.948 ± 0.04 | 113 | 2.9 ± 1.2 | 6 |
| CFP | 3.825 ± 0.06 | 150 | 2.8 ± 0.7 | 6 | |
| TFP | 4.013 ± 0.05 | 97 | 2.7 ± 1.3 | 5 | |
| Pip | 4.439 ± 0.03 * | 36 | 2.7 ± 0.7 | 4 | |
| 2MeP | 3.434 ± 0.06 * | 368 | 1.6 ± 0.3 | 5 | |
| 3MeP | 5.046 ± 0.05 * | 8.9 | 3.0 ± 2.7 | 4 | |
| Vah | 4.684 ± 0.03 * | 21 | 3.8 ± 0.5 | 4 | |
| P96 (loopA) | Pro | 3.951 ± 0.04 | 112 | 2.7 ± 1.1 | 5 |
| CFP | 3.840 ± 0.05 | 145 | 3.2 ± 0.7 | 4 | |
| TFP | 3.898 ± 0.03 | 127 | 2.9 ± 0.7 | 4 | |
| Pip | 3.667 ± 0.02 * | 215 | 2.9 ± 0.3 | 4 | |
| 2MeP | 3.596 ± 0.03 * | 254 | 2.8 ± 0.5 | 4 | |
| 3MeP | 3.289 ± 0.02 * | 514 | 3.0 ± 0.2 | 4 | |
| Vah | 4.029 ± 0.05 | 94 | 1.5 ± 0.3 | 4 | |
| P146 (CysL) | Pro | 4.052 ± 0.02 | 89 | 2.5 ± 0.4 | 6 |
| CFP | 4.350 ± 0.03 * | 45 | 3.7 ± 0.6 | 6 | |
| TFP | 3.806 ± 0.05 * | 156 | 2.3 ± 0.4 | 5 | |
| Pip | 4.279 ± 0.11 * | 53 | 1.9 ± 0.7 | 5 | |
| 2MeP | 3.568 ± 0.04 * | 270 | 2.2 ± 0.5 | 6 | |
| 3MeP | ND | ||||
| Vah | NR | 6 | |||
| P230 (M1) | Pro | 3.922 ± 0.06 | 119 | 2.5 ± 0.9 | 4 |
| CFP | 4.131 ± 0.04 | 74 | 1.4 ± 0.2 | 4 | |
| TFP | 3.904 ± 0.06 | 125 | 3.0 ± 1.4 | 4 | |
| Pip | 4.166 ± 0.08 | 68 | 1.1 ± 0.2 | 4 | |
| 2MeP | 3.751 ± 0.08 | 177 | 1.6 ± 0.5 | 4 | |
| 3MeP | 4.471 ± 0.08 * | 34 | 1.1 ± 0.4 | 4 | |
| Vah | 4.015 ± 0.02 | 97 | 2.6 ± 0.4 | 4 |
| Gly | Tau | |||||||
|---|---|---|---|---|---|---|---|---|
| pEC50, M | EC50 μM | nH | pEC50, M | EC50 μM | nH | % ImaxTau /ImaxGly | n | |
| Pro | 3.999 ± 0.03 | 100 | 3.2 ± 1.6 | 2.984 ± 0.07 | 1030 | 1.5 ± 0.3 | 43 ± 5 | 4 |
| CFP | 4.213 ± 0.04 * | 61 | 3.0 ± 0.5 | 2.139 ± 0.03 * | 579 | 2.1 ± 0.2 | 71 ± 3 * | 3 |
| Pip | 4.103 ± 0.05 * | 79 | 2.7 ± 0.9 | 3.121 ± 0.05 * | 757 | 1.8 ± 0.3 | 64 ± 7 * | 3 |
| 2MeP | 3.582 ± 0.02 * | 261 | 3.6 ± 0.2 | 2.497 ± 0.04 * | 3184 | 2.4 ± 0.6 | 18 ± 2 * | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lummis, S.C.R.; Dougherty, D.A. Expression of Mutant Glycine Receptors in Xenopus Oocytes Using Canonical and Non-Canonical Amino Acids Reveals Distinct Roles of Conserved Proline Residues. Membranes 2022, 12, 1012. https://doi.org/10.3390/membranes12101012
Lummis SCR, Dougherty DA. Expression of Mutant Glycine Receptors in Xenopus Oocytes Using Canonical and Non-Canonical Amino Acids Reveals Distinct Roles of Conserved Proline Residues. Membranes. 2022; 12(10):1012. https://doi.org/10.3390/membranes12101012
Chicago/Turabian StyleLummis, Sarah C. R., and Dennis A. Dougherty. 2022. "Expression of Mutant Glycine Receptors in Xenopus Oocytes Using Canonical and Non-Canonical Amino Acids Reveals Distinct Roles of Conserved Proline Residues" Membranes 12, no. 10: 1012. https://doi.org/10.3390/membranes12101012