Creating Supported Plasma Membrane Bilayers Using Acoustic Pressure

 ,
,  and
and 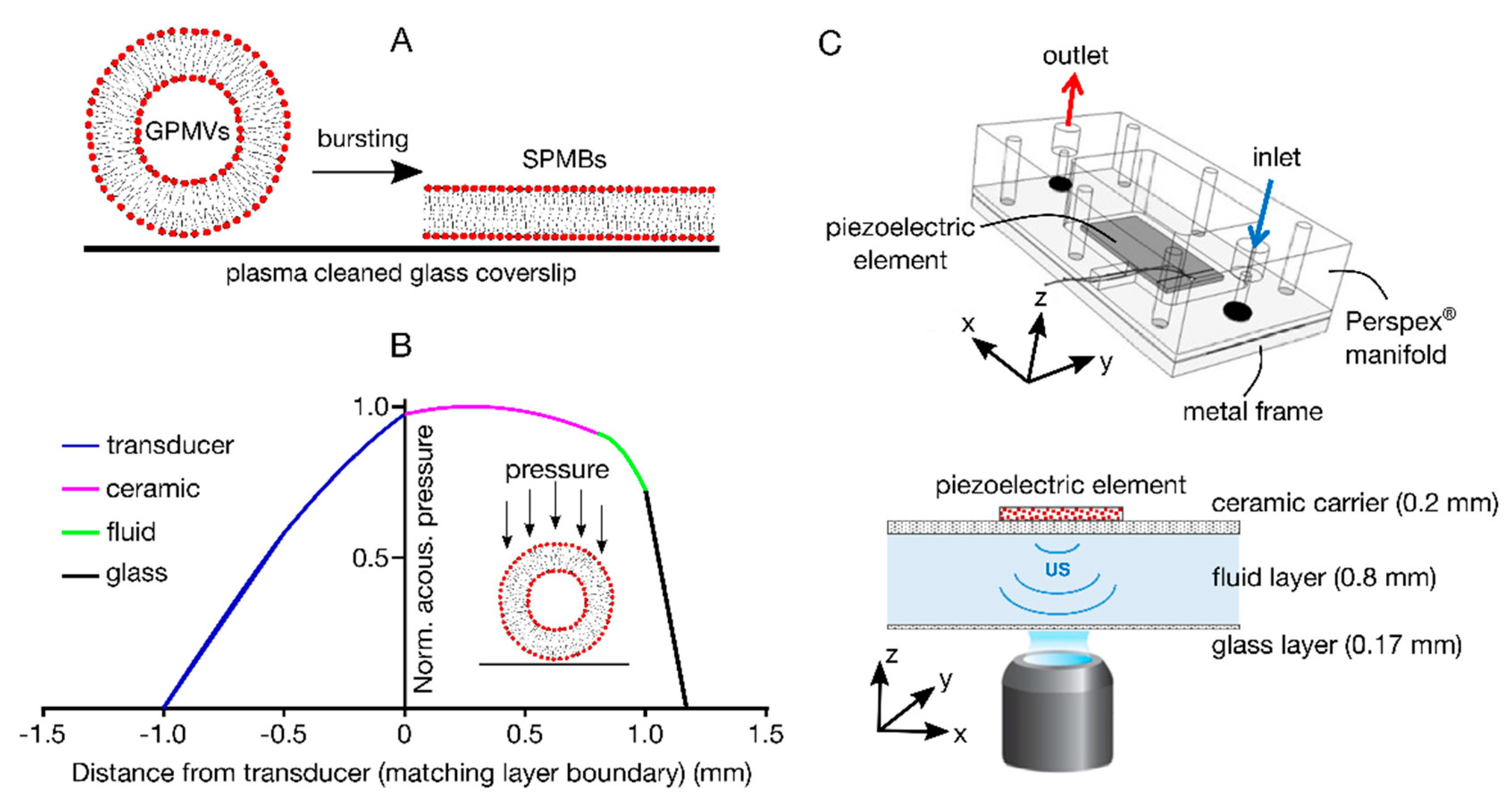{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Lipids and Proteins
2.2. Cleaning of Coverslips
2.3. Preparation of GPMVs
2.4. Acoustofluidic Device
2.5. Imaging and FCS
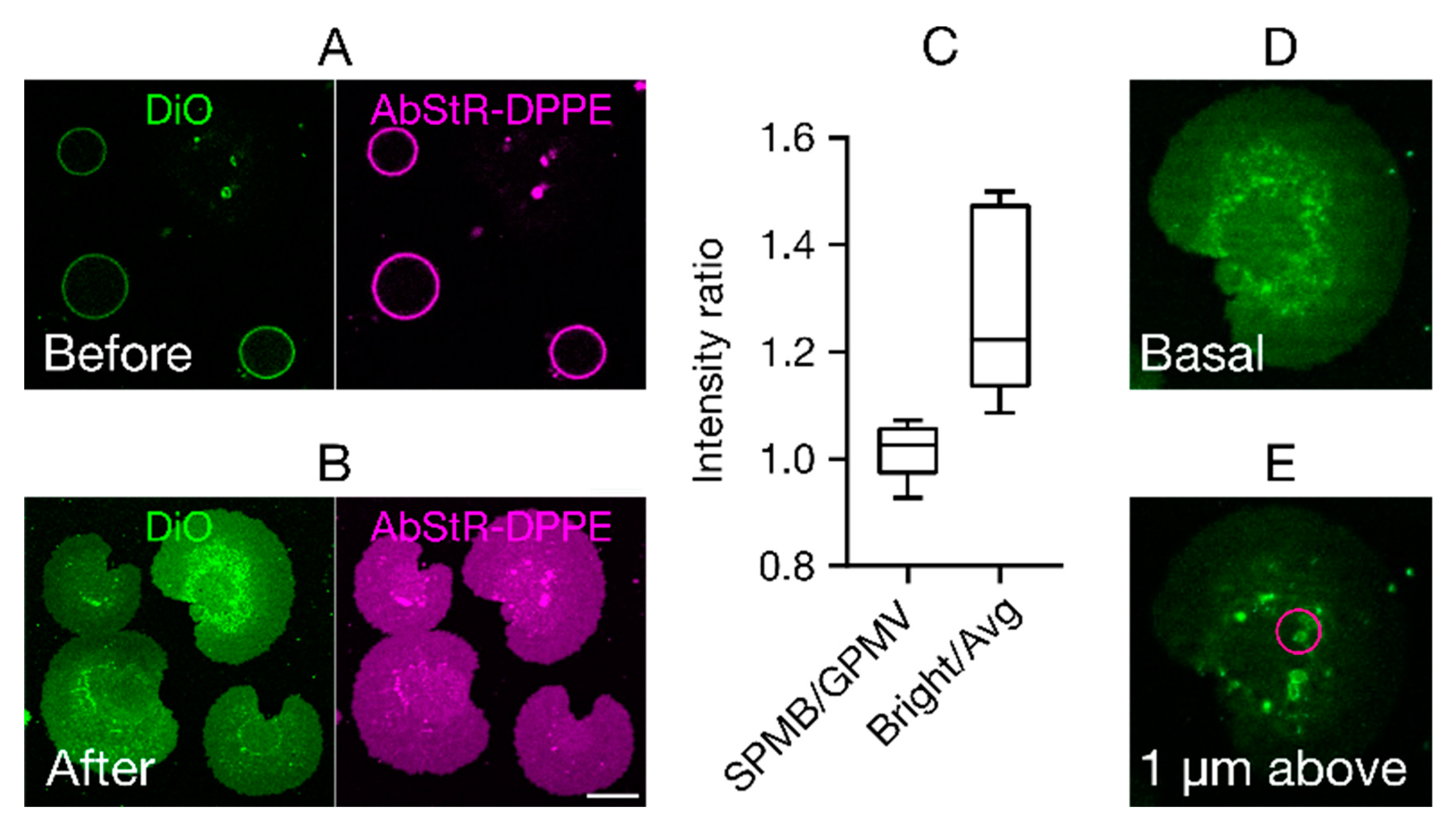
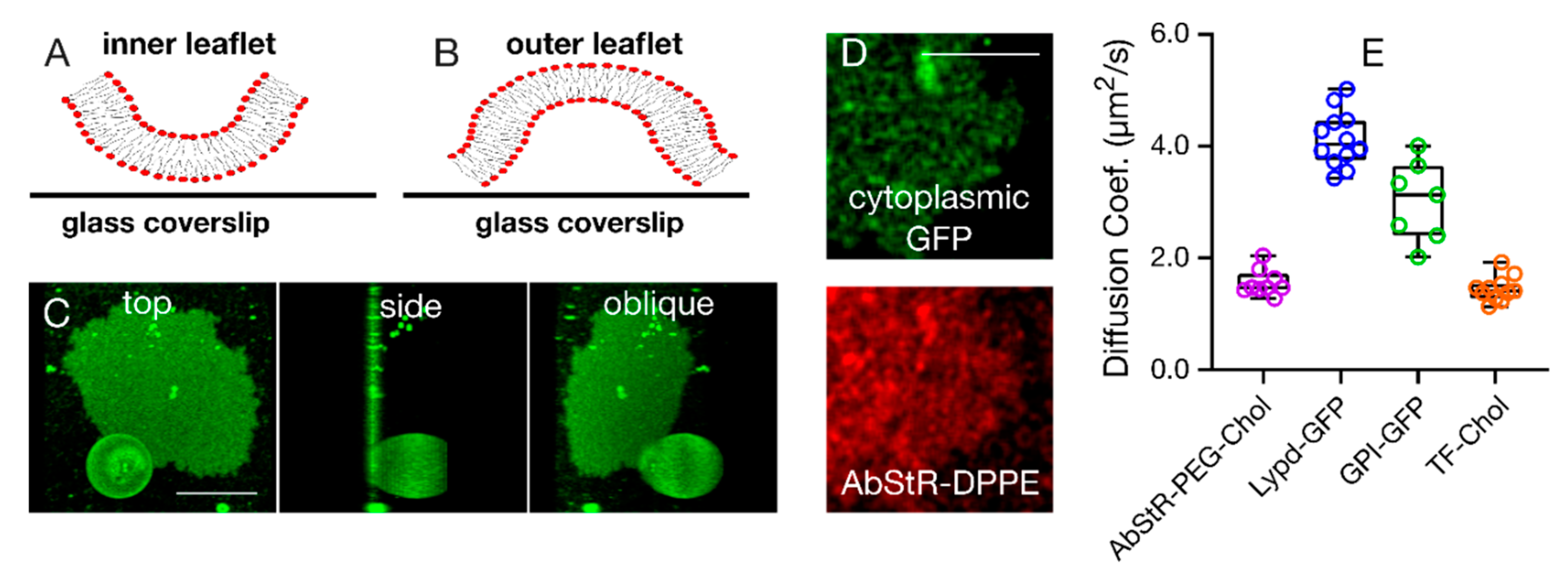
3. Results
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sezgin, E.; Schwille, P. Model membrane platforms to study protein-membrane interactions. Mol. Membr. Biol. 2012, 29, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, O.G. Model answers to lipid membrane questions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004622. [Google Scholar] [CrossRef] [PubMed]
- Tamm, L.K.; McConnell, H.M. Supported phospholipid-bilayers. Biophys. J. 1985, 47, 105–113. [Google Scholar] [CrossRef] [Green Version]
- El Kirat, K.; Morandat, S.; Dufrene, Y.F. Nanoscale analysis of supported lipid bilayers using atomic force microscopy. Biochim. Biophys. Acta-Biomembr. 2010, 1798, 750–765. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Vardhana, S.; Cammer, M.; Curado, S.; Santos, L.; Sheetz, M.P.; Dustin, M.L. T Lymphocyte myosin IIA is required for maturation of the immunological synapse. Front. Immunol. 2012, 3, 230. [Google Scholar] [CrossRef] [Green Version]
- Sezgin, E.; Kaiser, H.-J.; Baumgart, T.; Schwille, P.; Simons, K.; Levental, I. Elucidating membrane structure and protein behavior using giant plasma membrane vesicles. Nat. Protoc. 2012, 7, 1042–1051. [Google Scholar] [CrossRef]
- Baumgart, T.; Hammond, A.T.; Sengupta, P.; Hess, S.T.; Holowka, D.A.; Baird, B.A.; Webb, W.W. Large-scale fluid/fluid phase separation of proteins and lipids in giant plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2007, 104, 3165–3170. [Google Scholar] [CrossRef] [Green Version]
- Gray, E.; Karslake, J.; Machta, B.B.; Veatch, S. Liquid General Anesthetics Lower Critical Temperatures in Plasma Membrane Vesicles. Biophys. J. 2013, 105, 2751–2759. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Maxwell, K.; Sezgin, E.; Lu, M.; Liang, H.; Hancock, J.; Dial, E.J.; Lichtenberger, L.M.; Levental, I. Bile Acids Modulate Signaling by Functional Perturbation of Plasma Membrane Domains*. J. Biol. Chem. 2013, 288, 35660–35670. [Google Scholar] [CrossRef] [Green Version]
- Ozhan, G.; Sezgin, E.; Wehner, D.; Pfister, A.S.; Kuehl, S.J.; Kagermeier-Schenk, B.; Kuehl, M.; Schwille, P.; Weidinger, G. Lypd6 enhances Wnt/beta-catenin signaling by promoting Lrp6 phosphorylation in raft plasma membrane domains. Dev. Cell 2013, 26, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Pae, J.; Liivamagi, L.; Lubenets, D.; Arukuusk, P.; Langel, U.; Pooga, M. Glycosaminoglycans are required for translocation of amphipathic cell-penetrating peptides across membranes. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 1860–1867. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Levental, I.; Grzybek, M.; Schwarzmann, G.; Mueller, V.; Honigmann, A.; Belov, V.N.; Eggeling, C.; Coskun, U.; Simons, K.; et al. Partitioning, diffusion, and ligand binding of raft lipid analogs in model and cellular plasma membranes. Biochim. Biophys. Acta-Biomembr. 2012, 1818, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezgin, E.; Sadowski, T.; Simons, K. Measuring lipid packing of model and cellular membranes with environment sensitive probes. Langmuir 2014, 30, 8160–8166. [Google Scholar] [CrossRef] [PubMed]
- Honigmann, A.; Mueller, V.; Ta, H.; Schoenle, A.; Sezgin, E.; Hell, S.W.; Eggeling, C. Scanning STED-FCS reveals spatiotemporal heterogeneity of lipid interaction in the plasma membrane of living cells. Nat. Commun. 2014, 5, 5412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezgin, E.; Gutmann, T.; Buhl, T.; Dirkx, R.; Grzybek, M.; Coskun, U.; Solimena, M.; Simons, K.; Levental, I.; Schwille, P. Adaptive lipid packing and bioactivity in membrane domains. PLoS ONE 2015, 10, e0123930. [Google Scholar] [CrossRef]
- Levental, I.; Byfield, F.J.; Chowdhury, P.; Gai, F.; Baumgart, T.; Janmey, P.A. Cholesterol-dependent phase separation in cell-derived giant plasma-membrane vesicles. Biochem. J. 2009, 424, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, P.; Hammond, A.; Holowka, D.; Baird, B. Structural determinants for partitioning of lipids and proteins between coexisting fluid phases in giant plasma membrane vesicles. Biochim. Et Biophys. Acta-Biomembr. 2008, 1778, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Levental, K.R.; Levental, I. Isolation of giant plasma membrane vesicles for evaluation of plasma membrane structure and protein partitioning. Methods Mol. Biol. 2015, 1232, 65–77. [Google Scholar]
- Simonsson, L.; Gunnarsson, A.; Wallin, P.; Jönsson, P.; Höök, F. Continuous Lipid Bilayers Derived from Cell Membranes for Spatial Molecular Manipulation. J. Am. Chem. Soc. 2011, 133, 14027–14032. [Google Scholar] [CrossRef]
- Costello, D.A.; Hsia, C.Y.; Millet, J.K.; Porri, T.; Daniel, S. Membrane fusion-competent virus-like proteoliposomes and proteinaceous supported bilayers made directly from cell plasma membranes. Langmuir 2013, 29, 6409–6419. [Google Scholar] [CrossRef]
- Richards, M.J.; Hsia, C.Y.; Singh, R.R.; Haider, H.; Kumpf, J.; Kawate, T.; Daniel, S. Membrane protein mobility and orientation preserved in supported bilayers created directly from cell plasma membrane blebs. Langmuir 2016, 32, 2963–2974. [Google Scholar] [CrossRef] [PubMed]
- Chiang, P.C.; Tanady, K.; Huang, L.T.; Chao, L. Rupturing giant plasma membrane vesicles to form micron-sized supported cell plasma membranes with native transmembrane proteins. Sci. Rep. 2017, 7, 15139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.T.; Kreutzberger AJ, B.; Kiessling, V.; Ganser-Pornillos, B.K.; White, J.M.; Tamm, L.K. HIV virions sense plasma membrane heterogeneity for cell entry. Sci. Adv. 2017, 3, e1700338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carugo, D.; Octon, T.; Messaoudi, W.; Fisher, A.L.; Carboni, M.; Harris, N.R.; Hill, M.; Glynne-Jones, P. A thin-reflector microfluidic resonator for continuous-flow concentration of microorganisms: A new approach to water quality analysis using acoustofluidics. Lab Chip 2014, 14, 3830–3842. [Google Scholar] [CrossRef] [Green Version]
- Schneider, F.; Waithe, D.; Clausen, M.P.; Galiani, S.; Koller, T.; Ozhan, G.; Eggeling, C.; Sezgin, E. Diffusion of lipids and GPI-anchored proteins in actin-free plasma membrane vesicles measured by STED-FCS. Mol. Biol. Cell 2017, 28, 1507–1518. [Google Scholar] [CrossRef] [Green Version]
- Carugo, D.; Aron, M.; Sezgin, E.; Bernardino de la Serna, J.; Kuimova, M.K.; Eggeling, C.; Stride, E. Modulation of the molecular arrangement in artificial and biological membranes by phospholipid-shelled microbubbles. Biomaterials 2017, 113, 105–117. [Google Scholar] [CrossRef]
- Hill, M.; Wood, R.J. Modelling in the design of a flow-through ultrasonic separator. Ultrasonics 2000, 38, 662–665. [Google Scholar] [CrossRef]
- Waithe, D.; Clausen, M.P.; Sezgin, E.; Eggeling, C. FoCuS-point: Software for STED fluorescence correlation and time-gated single photon counting. Bioinformatics 2015, 32, 958–960. [Google Scholar] [CrossRef] [Green Version]
- Glynne-Jones, P.; Boltryk, R.J.; Hill, M.; Harris, N.R.; Baclet, P. Robust acoustic particle manipulation: A thin-reflector design for moving particles to a surface. JASA 2009, 126, El75–El79. [Google Scholar] [CrossRef] [Green Version]
- Honigmann, A.; Mueller, V.; Hell, S.W.; Eggeling, C. STED microscopy detects and quantifies liquid phase separation in lipid membranes using a new far-red emitting fluorescent phosphoglycerolipid analogue. Faraday Discuss. 2013, 161, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Pinkwart, K.; Schneider, F.; Lukoseviciute, M.; Sauka-Spengler, T.; Lyman, E.; Eggeling, C.; Sezgin, E. Nanoscale dynamics of cholesterol in the cell membrane. J. Biol. Chem. 2019, 294, 12599–12609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, B.J.; Kenworthy, A.K.; Polishchuk, R.S.; Lodge, R.; Roberts, T.H.; Hirschberg, K.; Phair, R.D.; Lippincott-Schwartz, J. Rapid Cycling of Lipid Raft Markers between the Cell Surface and Golgi Complex. J. Cell Biol. 2001, 153, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Keller, H.; Lorizate, M.; Schwille, P. PI(4,5)P-2 Degradation promotes the formation of cytoskeleton-free model membrane systems. ChemPhysChem 2009, 10, 2805–2812. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sezgin, E.; Carugo, D.; Levental, I.; Stride, E.; Eggeling, C. Creating Supported Plasma Membrane Bilayers Using Acoustic Pressure. Membranes 2020, 10, 30. https://doi.org/10.3390/membranes10020030
Sezgin E, Carugo D, Levental I, Stride E, Eggeling C. Creating Supported Plasma Membrane Bilayers Using Acoustic Pressure. Membranes. 2020; 10(2):30. https://doi.org/10.3390/membranes10020030
Chicago/Turabian StyleSezgin, Erdinc, Dario Carugo, Ilya Levental, Eleanor Stride, and Christian Eggeling. 2020. "Creating Supported Plasma Membrane Bilayers Using Acoustic Pressure" Membranes 10, no. 2: 30. https://doi.org/10.3390/membranes10020030