
Evaluation of Four Commercial Vaccines for the Protection of Piglets against the Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (hp-PRRSV) QH-08 Strain

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. PRRSV Strains Sequence Profiles from GenBank and Analysis
2.2. Cells, Virus Isolation, and Vaccines
2.3. Animal Vaccine/Challenge Design
2.4. Clinical Examinations
2.5. Serology
2.6. RT-PCR for Antigen Detection
2.7. Statistical Analysis
3. Results
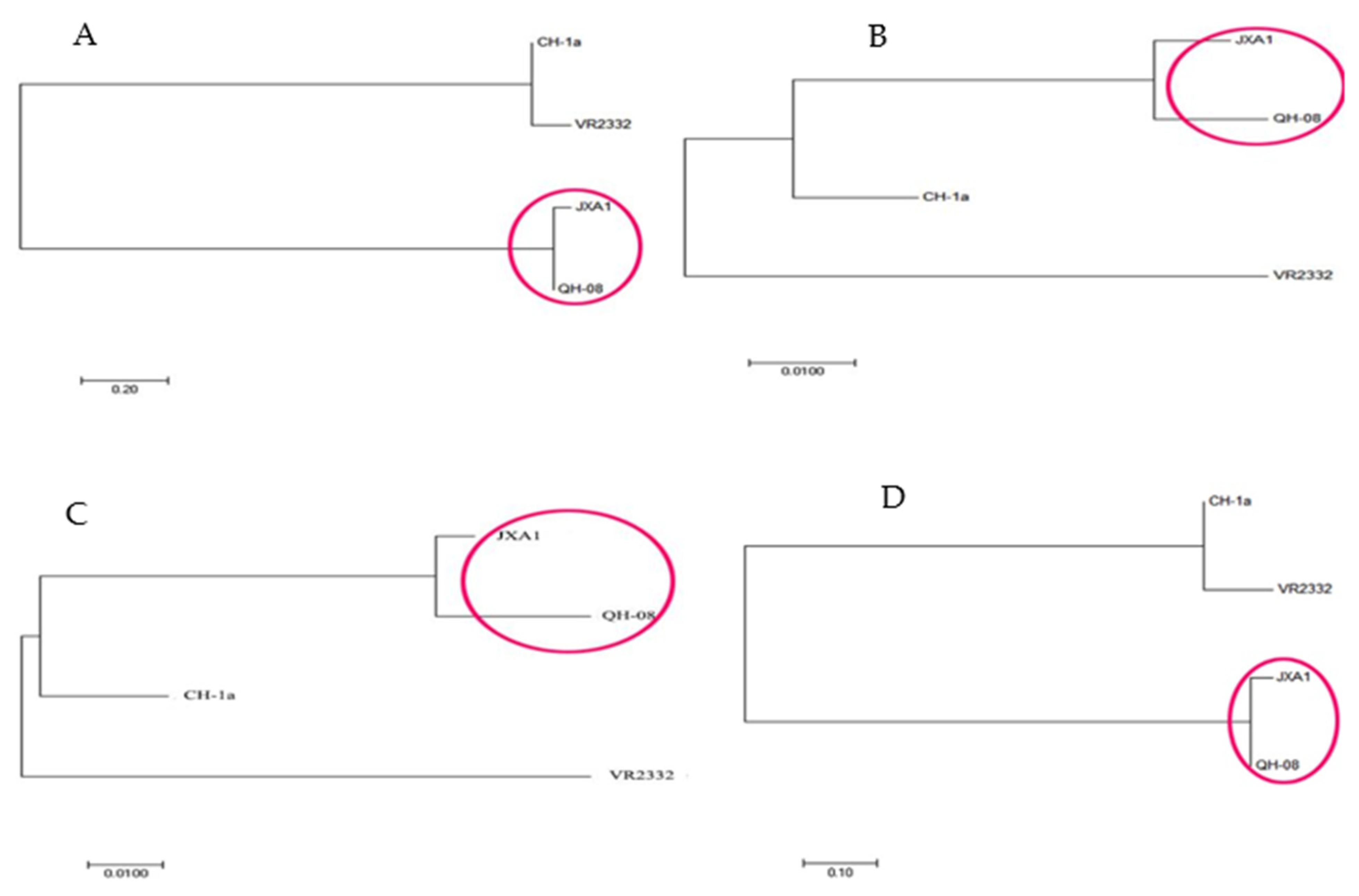
3.1. Gene Identity and Phylogenetic Analysis
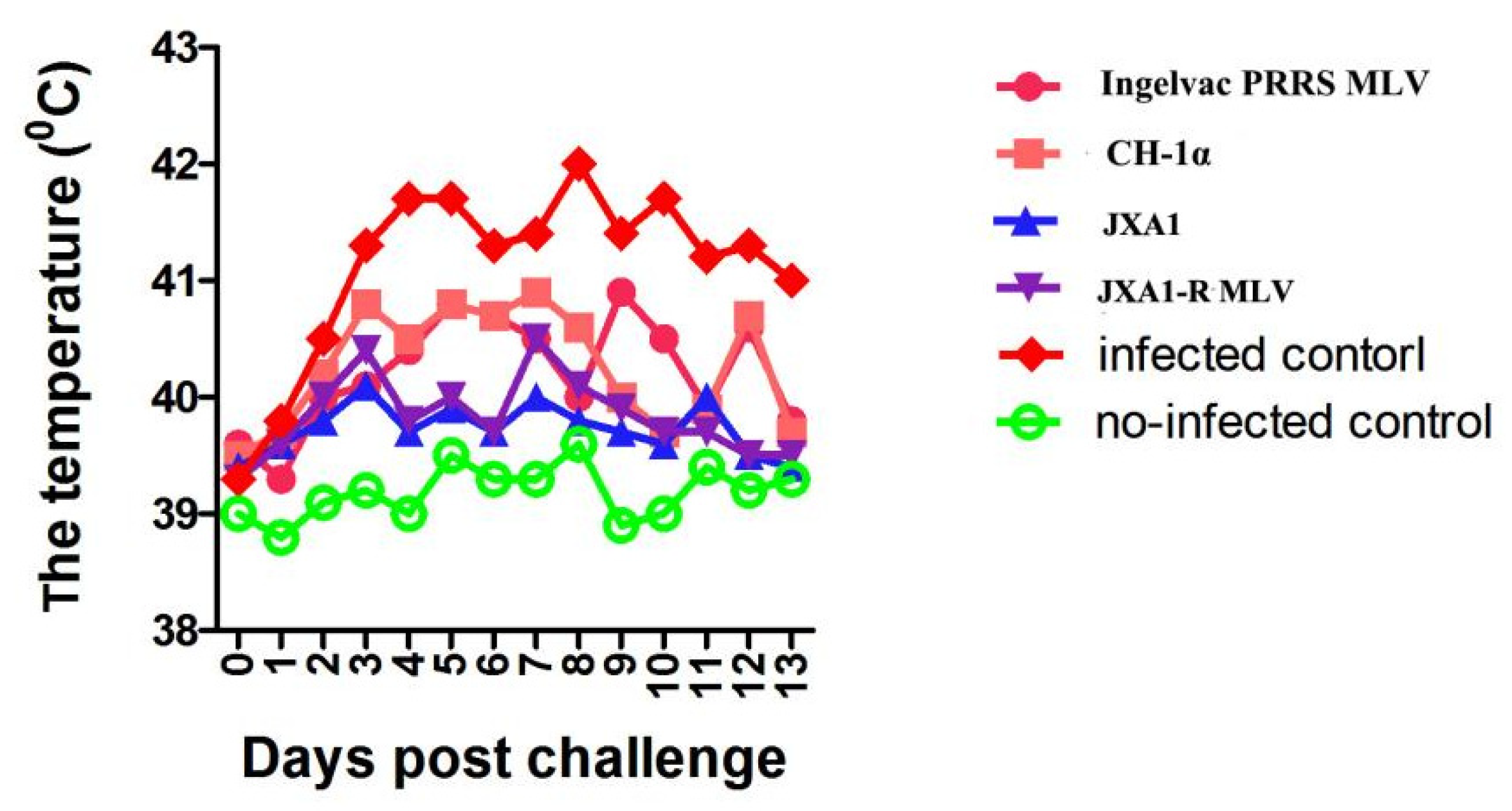
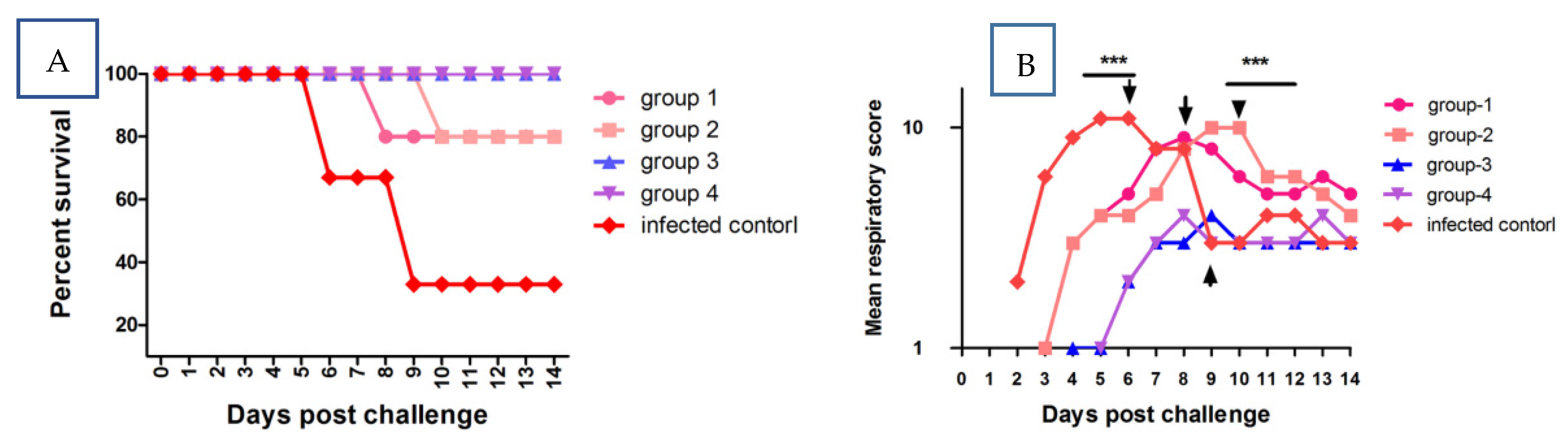
3.2. Clinical Observation
3.3. Detection of Virus RNA in Sera and Lung Tissue
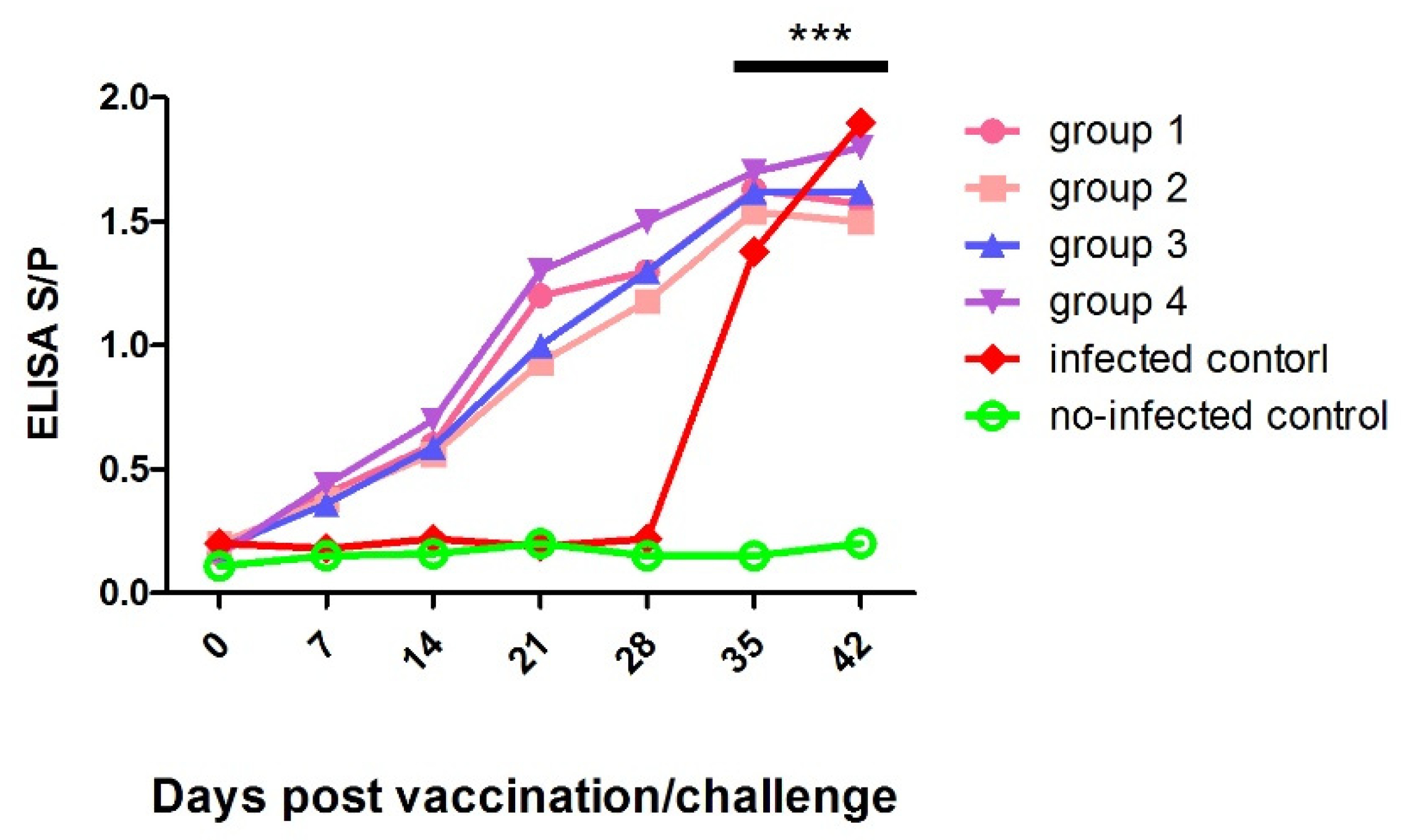
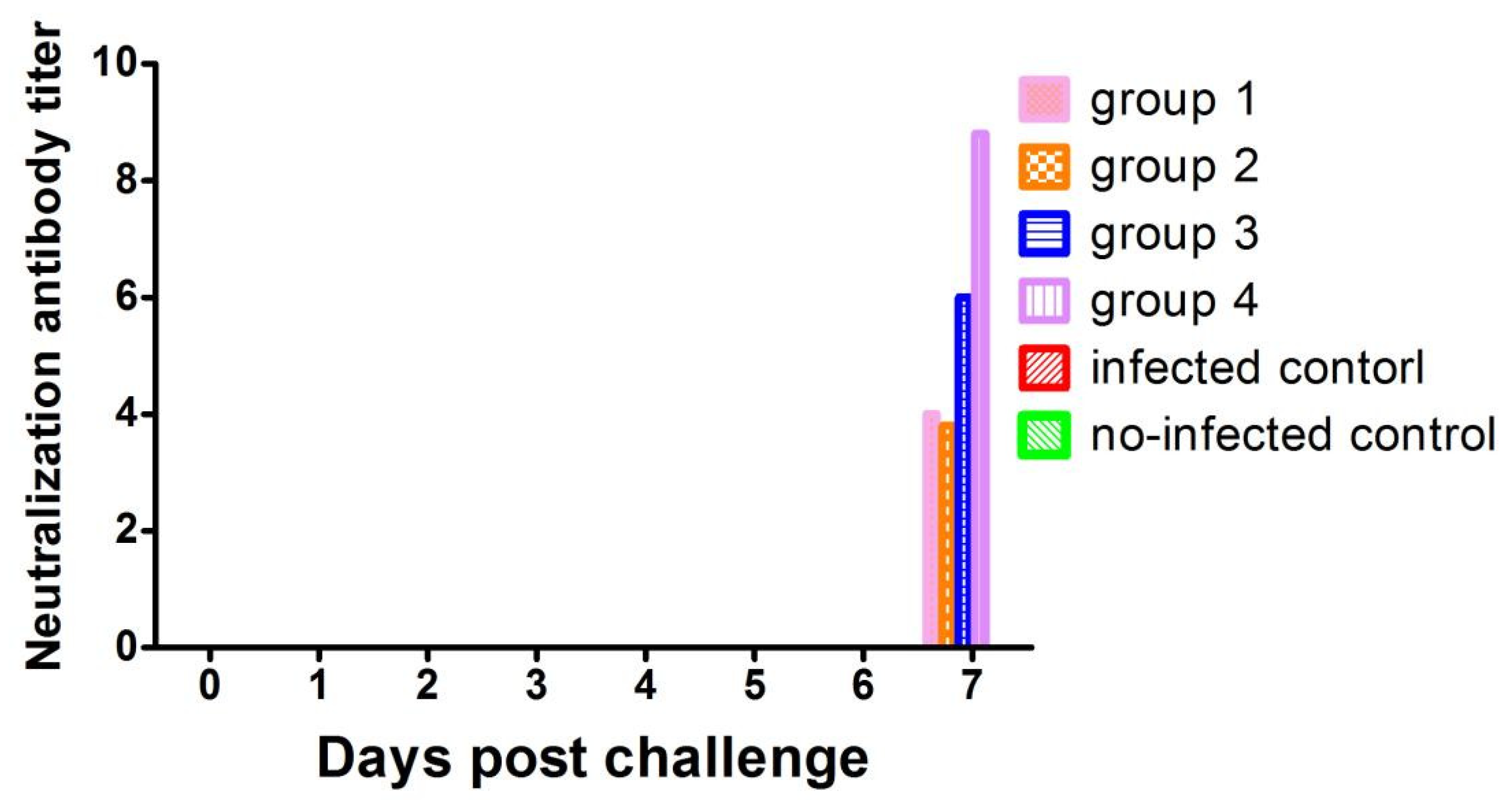
3.4. Detection of Serum Antibodies against PRRSV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, L.; Yang, H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010, 154, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.S.; Zhou, J.H.; Chen, H.T.; Ma, L.N.; Ding, Y.Z.; Wang, M.; Zhang, J. Analysis of synonymous codon usage in porcine reproductive and respiratory syndrome virus. Infect. Genet. Evol. 2010, 10, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.Z.; You, Y.N.; Sun, D.J.; Chen, H.T.; Wang, Y.L.; Chang, H.Y.; Pan, L.; Fang, Y.Z.; Zhang, Z.W.; Zhou, P.; et al. The Effects of the Context-Dependent Codon Usage Bias on the Structure of the nsp1 alpha of Porcine Reproductive and Respiratory Syndrome Virus. Biomed. Res. Int. 2014, 2014, 765320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.Z.; Li, G.X.; Cheng, F.Z.; Dai, J.F.; Hu, Y.H.; Zhang, J. Yansuanmalingua inhibits replication of type 2 porcine reproductive and respiratory syndrome virus via activating the caspase-8 apoptosis pathway. J. Basic Microb. 2020, 60, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Kikkert, M.; Fang, Y. Arterivirus molecular biology and pathogenesis. J. Gen. Virol. 2013, 94, 2141–2163. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; McCullough, S.; Morrison, R.B.; Joo, H.S.; et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Investig. 1992, 4, 117–126. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef]
- Anping, W.; Qi, C.; Leyi, W.; Darin, M.; Karen, H.; Phillip, G.; Jianqiang, Z.; Ganwu, L. Recombination between Vaccine and Field Strains of Porcine Reproductive and Respiratory Syndrome Virus. Emerg. Infect. Dis. 2019, 25, 2335–2337. [Google Scholar]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bào, Y.; Ng, T.F.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Darwich, L.; Gimeno, M.; Sibila, M.; Diaz, I.; de la Torre, E.; Dotti, S.; Kuzemtseva, L.; Martin, M.; Pujols, J.; Mateu, E. Genetic and immunobiological diversities of porcine reproductive and respiratory syndrome genotype I strains. Vet. Microbiol. 2011, 150, 49–62. [Google Scholar] [CrossRef]
- Do, D.T.; Park, C.; Choi, K.; Jeong, J.; Nguyen, T.T.; Nguyen, K.D.; Vo, D.T.; Chae, C. Comparison of two genetically distant type 2 porcine reproductive and respiratory syndrome virus (PRRSV) modified live vaccines against Vietnamese highly pathogenic PRRSV. Vet. Microbiol. 2015, 179, 233–241. [Google Scholar] [CrossRef]
- Charoenchanikran, P.; Kedkovid, R.; Sirisereewan, C.; Woonwong, Y.; Arunorat, J.; Sitthichareonchai, P.; Sopipan, N.; Jittimanee, S.; Kesdangsakonwut, S.; Thanawongnuwech, R. Efficacy of Fostera(R) PRRS modified live virus (MLV) vaccination strategy against a Thai highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection. Trop. Anim. Health Prod. 2016, 48, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Sirisereewan, C.; Woonwong, Y.; Arunorat, J.; Kedkovid, R.; Nedumpun, T.; Kesdangsakonwut, S.; Suradhat, S.; Thanawongnuwech, R.; Teankum, K. Efficacy of a type 2 PRRSV modified live vaccine (PrimePac PRRS) against a Thai HP-PRRSV challenge. Trop. Anim. Health Prod. 2018, 50, 1509–1518. [Google Scholar] [CrossRef]
- An, T.Q.; Zhou, Y.J.; Liu, G.Q.; Tian, Z.J.; Li, J.; Qiu, H.J.; Tong, G.Z. Genetic diversity and phylogenetic analysis of glycoprotein 5 of PRRSV isolates in mainland China from 1996 to 2006: Coexistence of two NA-subgenotypes with great diversity. Vet. Microbiol. 2007, 123, 43–52. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, J.; Zhuang, J.; Sun, Z.; Gao, F.; Yuan, S. Immunization of pigs with a type 2 modified live PRRSV vaccine prevents the development of a deadly long lasting hyperpyrexia in a challenge study with highly pathogenic PRRSV JX143. Vaccine 2013, 31, 2062–2066. [Google Scholar] [CrossRef]
- Thanawongnuwech, R.; Halbur, P.G.; Ackermann, M.R.; Thacker, E.L.; Royer, R.L. Effects of low (modified-live virus vaccine) and high (VR-2385)-virulence strains of porcine reproductive and respiratory syndrome virus on pulmonary clearance of copper particles in pigs. Vet. Pathol. 1998, 35, 398–406. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Li, G.; Yu, L.; Li, L.; Zhang, Y.; Zhou, Y.; Tong, W.; Liu, C.; Gao, F.; Tong, G. Genetic Diversity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) From 1996 to 2017 in China. Front. Microbiol. 2020, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhou, Z.; Cao, Z.; Wu, J.; Zhang, Z.; Xu, B.; Wang, C.; Hu, D.; Deng, X.; Han, W.; et al. Assessment of the safety and efficacy of an attenuated live vaccine based on highly pathogenic porcine reproductive and respiratory syndrome virus. Clin. Vaccine Immunol. CVI 2015, 22, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarque, G.; Reeth, K.V.; Nauwynck, H.; Drexler, C.; Van Gucht, S.; Pensaert, M. Impact of genetic diversity of European-type porcine reproductive and respiratory syndrome virus strains on vaccine efficacy. Vaccine 2004, 22, 4183–4190. [Google Scholar] [CrossRef]
- Li, X.; Galliher-Beckley, A.; Pappan, L.; Trible, B.; Kerrigan, M.; Beck, A.; Hesse, R.; Blecha, F.; Nietfeld, J.C.; Rowland, R.R.; et al. Comparison of host immune responses to homologous and heterologous type II porcine reproductive and respiratory syndrome virus (PRRSV) challenge in vaccinated and unvaccinated pigs. BioMed Res. Int. 2014, 2014, 416727. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.I.; Kim, J.J.; Cha, S.H.; Wu, W.H.; Cooper, V.; Evans, R.; Choi, E.J.; Yoon, K.J. Significance of genetic variation of Significance of genetic variation of PRRSV ORF5 in virus neutralization and molecular determinants corresponding to cross neutralization among PRRS viruses. Vet. Microbiol. 2013, 162, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Mengeling, W.L.; Vorwald, A.C.; Lager, K.M.; Clouser, D.F.; Wesley, R.D. Identification and clinical assessment of suspected vaccine-related field strains of porcine reproductive and respiratory syndrome virus. Am. J. Vet. Res. 1999, 60, 334–340. [Google Scholar]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhuang, J.; Wang, J.; Han, L.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; et al. Outbreak Investigation of NADC30-Like PRRSV in South-East China. Transbound. Emerg. Dis. 2016, 63, 474–479. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Isolate | Genebank | Y | Whole | ORF1a-1b | ORF2 | ORF3 | ORF4 | ORF5 | ORF6 | ORF7 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | CH-1a | AY032626 | 1998 | 40% | 28.2% | 96.1% | 92.9% | 97.2% | 91.5% | 96.8% | 92.7% |
| 2 | JXA1 | EF112445 | 2007 | 97.2% | 96.3% | 99.6% | 98.4% | 95.5% | 97.5% | 98.9% | 99.2 |
| 3 | QH-08 | KU201579 | 2008 | - | - | - | - | - | - | - | - |
| 4 | VR2332 | EF536003 | 2007 | 37.9% | 25.3% | 91% | 85.8% | 89.9% | 86.5% | 96.6% | 93.5 |
| Groups | Vaccines Injected | Dose | Challenge Virus | Route of Injection | |
|---|---|---|---|---|---|
| G1 | 5 pigs/each | Ingelvac PRRS MLV | 2 mL/piglet | (QH-08 at (2 × 106 TCID50) 2 mL/piglet | Intramuscularly |
| G2 | CH-1α | 2 mL/piglet | |||
| G3 | JXA1 | 2 mL/piglet | |||
| G4 | JXA1-R MLV | 2 mL/piglet | |||
| infected control | 3 pigs/each | No vaccine | PBS-2 mL/piglet | ||
| non-infected control | PBS-2 mL/piglet | ||||
| Experimental Groups | ||||||
|---|---|---|---|---|---|---|
| G1 | G2 | G3 | G4 | Infected Control | Non-Infected Control | |
| Vaccines | Ingelvac PRRS MLV | CH-1α | JXA1 | JXA1-R MLV | No | No |
| challenge | QH-08 | no | ||||
| Macroscopic lung lesion score | 55 ± 5.4 * | 54 ± 4.8 * | 32 ± 3.1 | 30 ± 2.8 | 82 ± 5 * | 0/3 |
| Microscopic lung lesion score | 2.4 ± 0.3 | 2.3 ± 0.24 | 1.3 ± 0.2 | 1.1 ± 0.1 | 3.4 ± 5.4 * | 0/3 |
| Virus isolation | 4/5 a* | 3/5 a* | 1/5 | 1/5 | 3/3 a* | 0/3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Wubshet, A.K.; Ding, X.; Zhang, Z.; Li, Q.; Dai, J.; Hou, Q.; Hu, Y.; Zhang, J. Evaluation of Four Commercial Vaccines for the Protection of Piglets against the Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (hp-PRRSV) QH-08 Strain. Vaccines 2021, 9, 1020. https://doi.org/10.3390/vaccines9091020
Ding Y, Wubshet AK, Ding X, Zhang Z, Li Q, Dai J, Hou Q, Hu Y, Zhang J. Evaluation of Four Commercial Vaccines for the Protection of Piglets against the Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (hp-PRRSV) QH-08 Strain. Vaccines. 2021; 9(9):1020. https://doi.org/10.3390/vaccines9091020
Chicago/Turabian StyleDing, Yaozhong, Ashenafi Kiros Wubshet, Xiaolong Ding, Zhongwang Zhang, Qian Li, Junfei Dai, Qian Hou, Yonghao Hu, and Jie Zhang. 2021. "Evaluation of Four Commercial Vaccines for the Protection of Piglets against the Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (hp-PRRSV) QH-08 Strain" Vaccines 9, no. 9: 1020. https://doi.org/10.3390/vaccines9091020