
The Promise and Challenges of Cyclic Dinucleotides as Molecular Adjuvants for Vaccine Development


Abstract
:1. Introduction
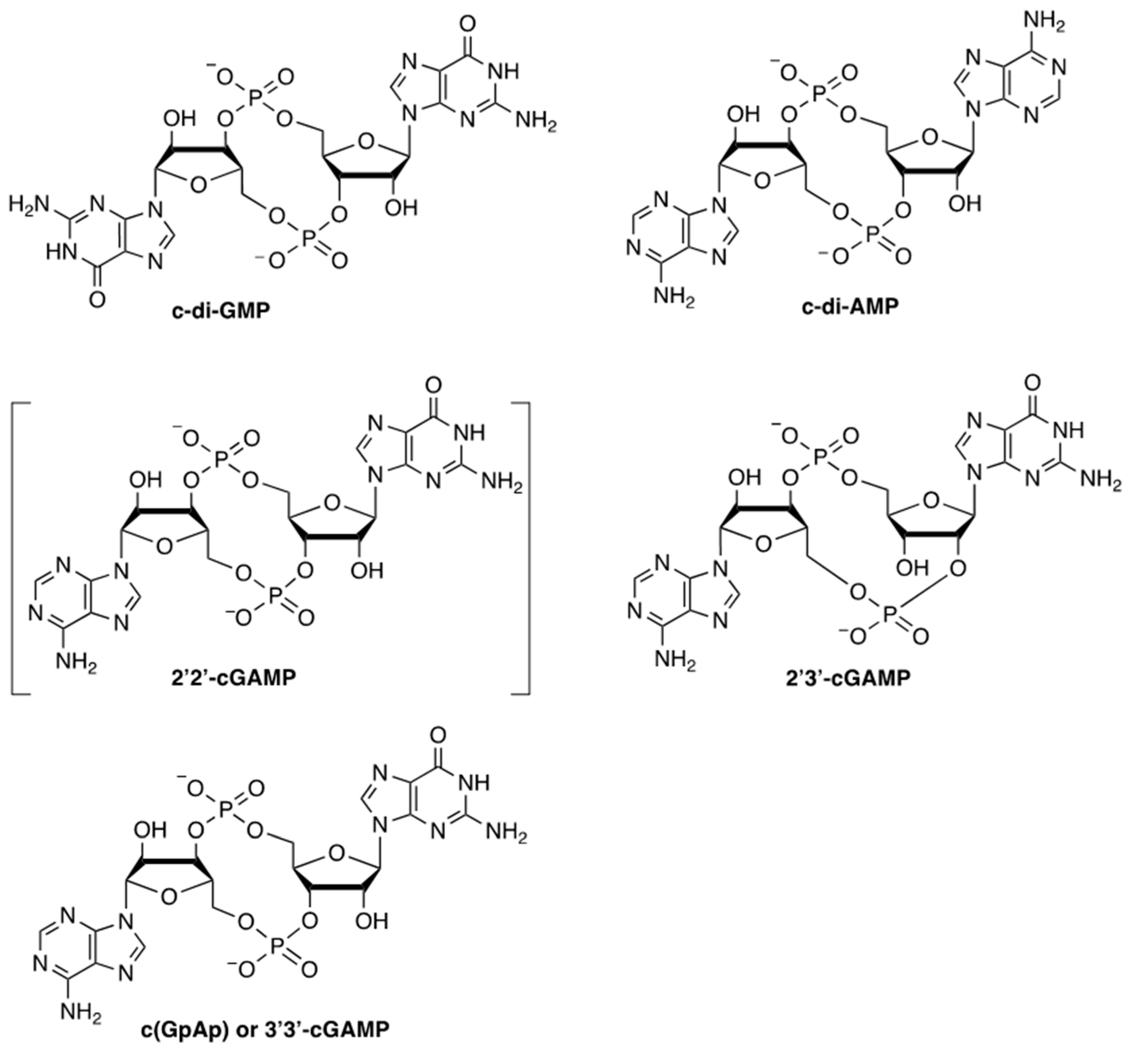
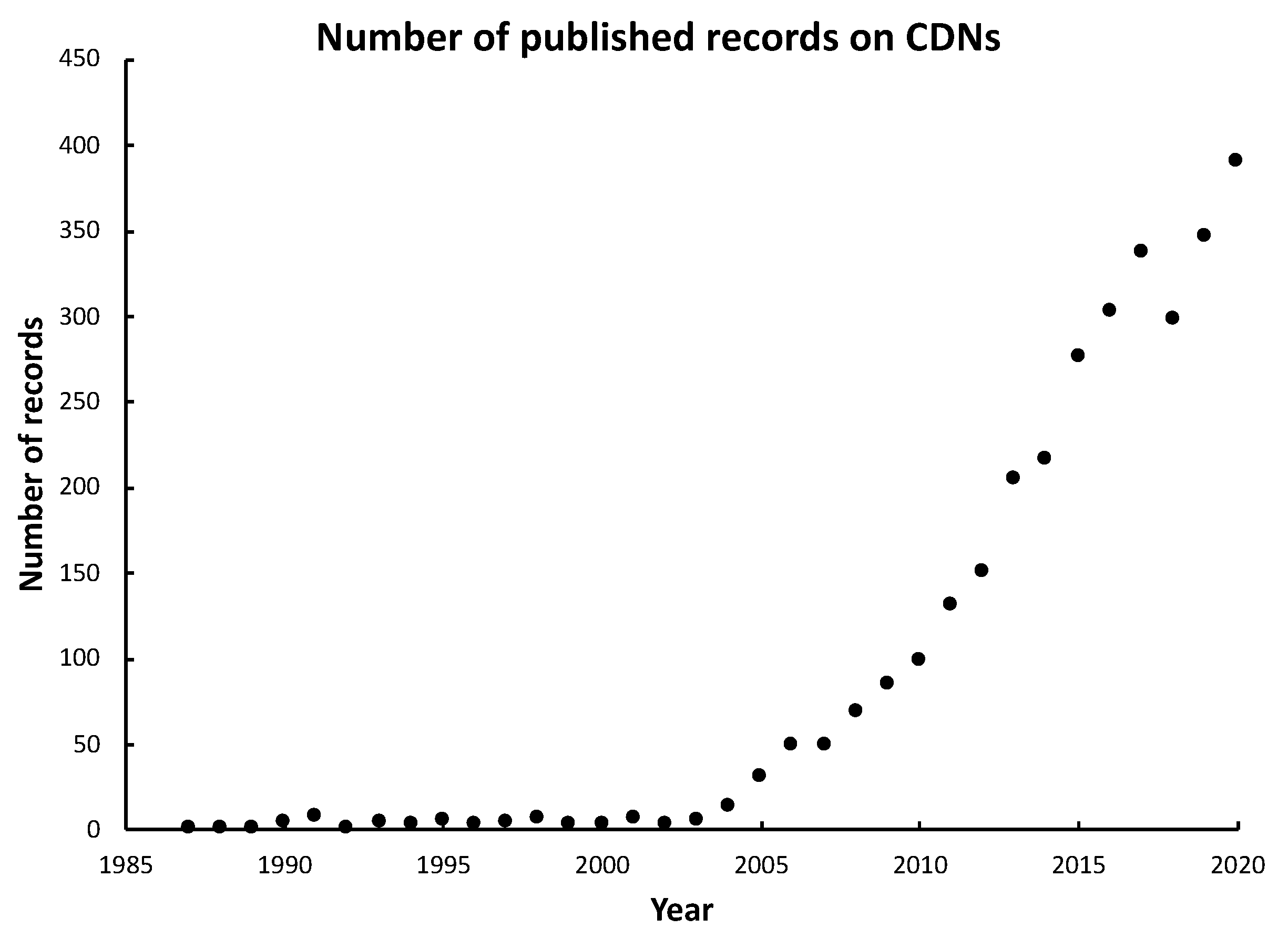
2. Discovery and Chemical Structure of CDNs
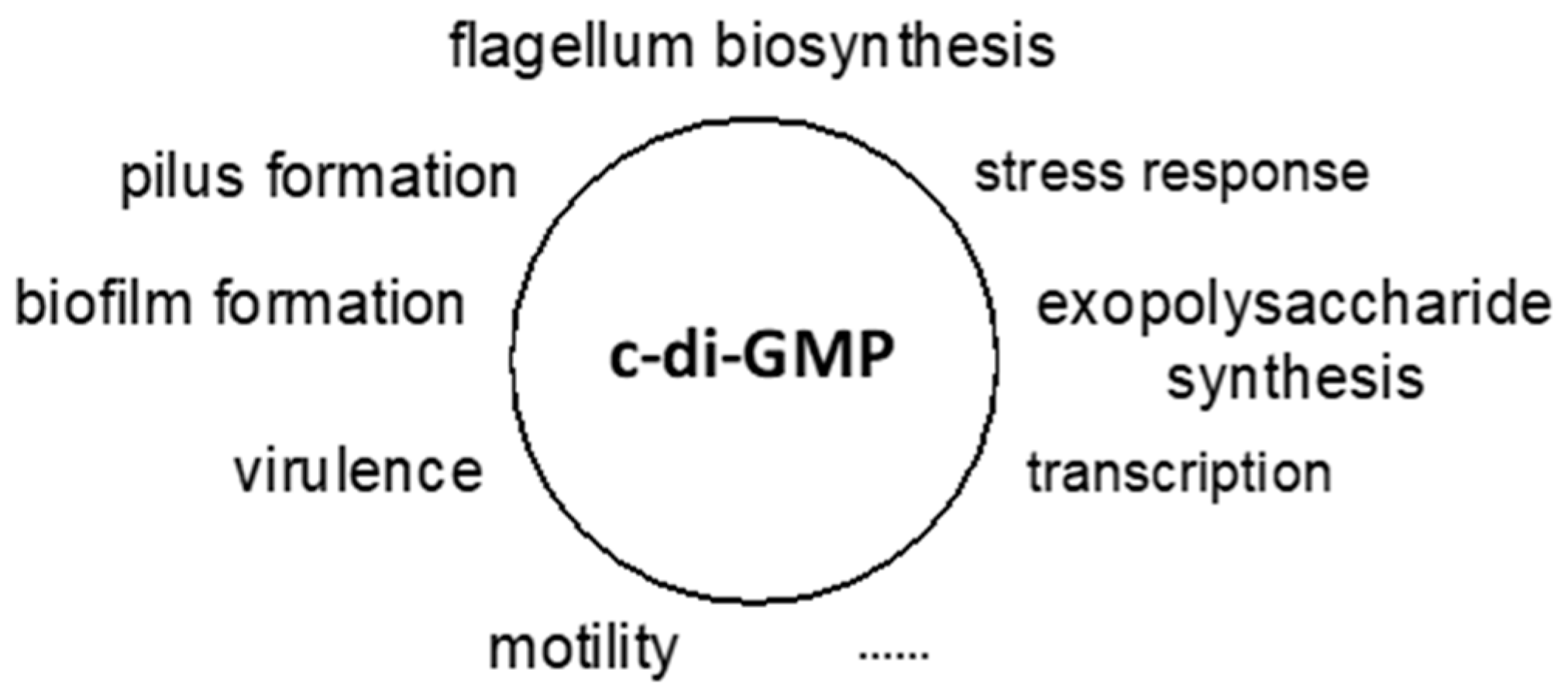
3. c-di-GMP as a Universal Bacterial Second Messenger
4. c-di-GMP and Analogs Generated to Date
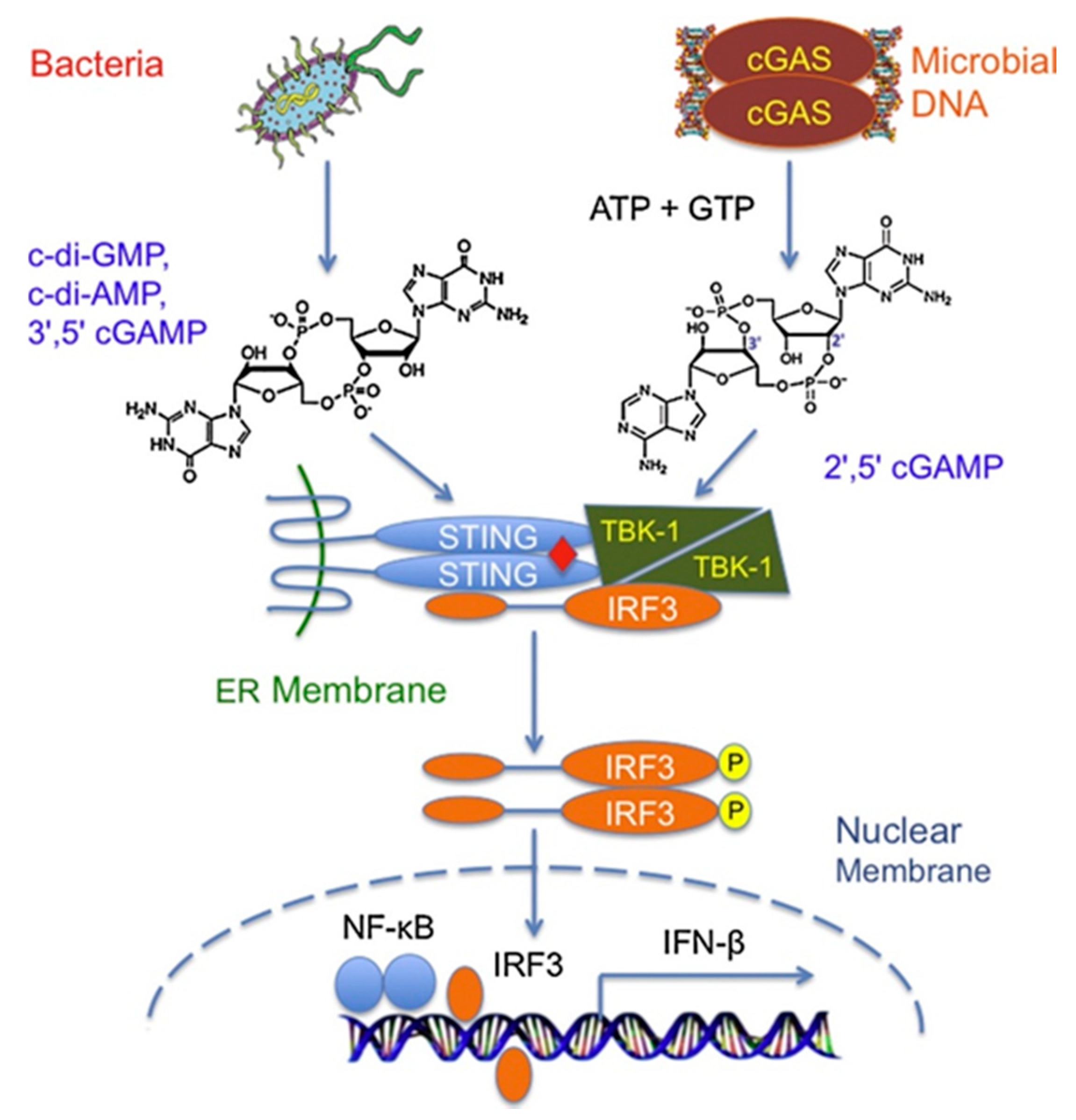
5. Immunostimulatory Function of CDNs in Host Innate Immune Responses
5.1. In Vitro Immunostimulatory Functions of CDNs
5.2. In Vivo Immunostimulatory Functions of CDNs
6. CDNs as Potent Vaccine Adjuvants for Systemic and Mucosal Immunization
6.1. CDNs as Adjuvants for the Development of Vaccines against Infectious Diseases
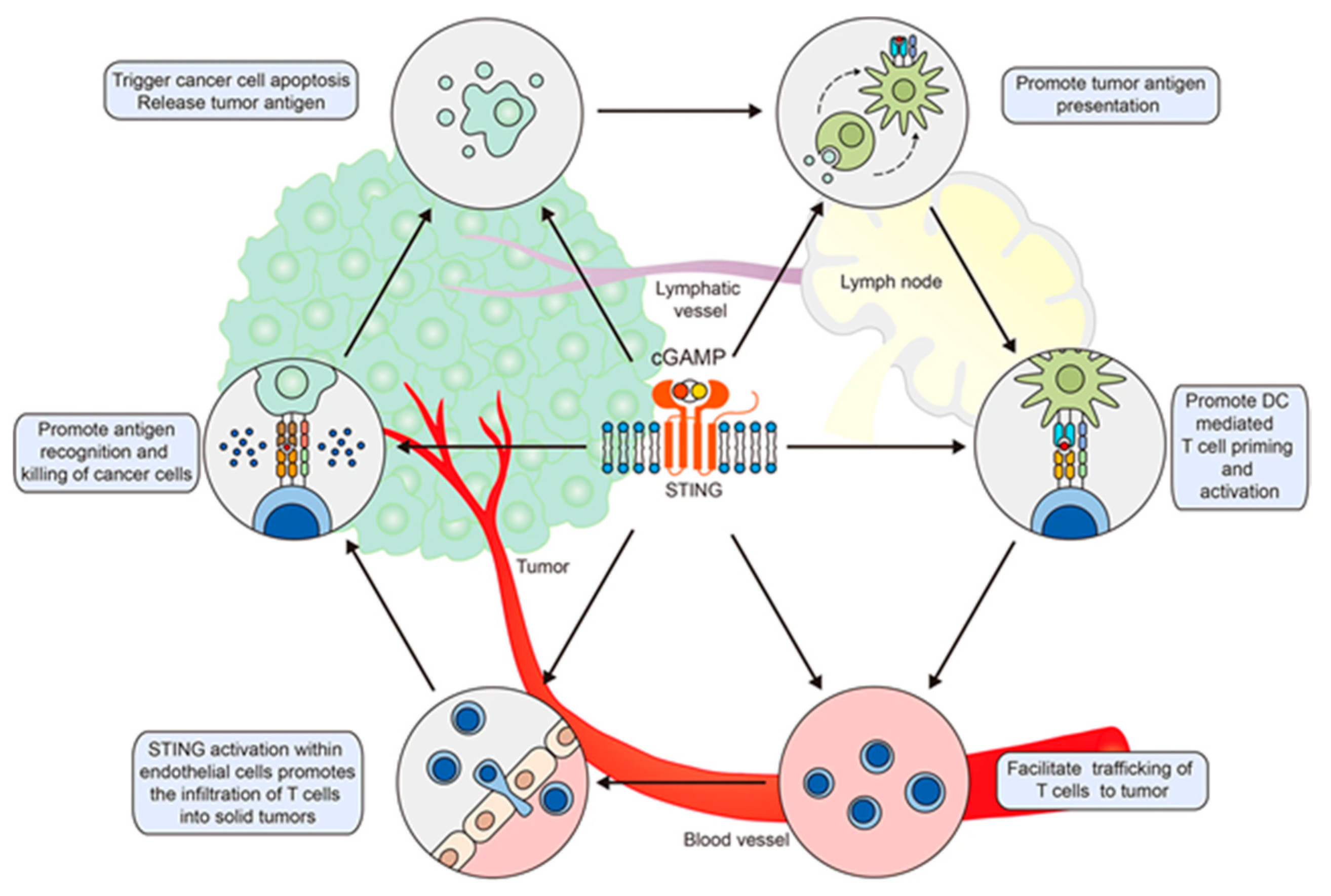
6.2. CDNs as Adjuvants for the Development of Cancer Vaccines
7. Novel Delivery Strategies to Enhance the Immunostimulatory and Adjuvant Function of CDNs
8. Conclusions and Future Direction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahn, J.; Barber, G.N. STING signaling and host defense against microbial infection. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Burdette, D.L.; Monroe, K.M.; Sotelo-Troha, K.; Iwig, J.S.; Eckert, B.; Hyodo, M.; Hayakawa, Y.; Vance, R.E. STING is a direct innate immune sensor of cyclic di-GMP. Nature 2011, 478, 515–518. [Google Scholar] [CrossRef]
- Ouyang, S.; Song, X.; Wang, Y.; Ru, H.; Shaw, N.; Jiang, Y.; Niu, F.; Zhu, Y.; Qiu, W.; Parvatiyar, K.; et al. Structural Analysis of the STING Adaptor Protein Reveals a Hydrophobic Dimer Interface and Mode of Cyclic di-GMP Binding. Immunity 2012, 36, 1073–1086. [Google Scholar] [CrossRef] [Green Version]
- Barber, G.N. STING-dependent cytosolic DNA sensing pathways. Trends Immunol. 2014, 35, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-J.; Zhao, L.; Hu, H.; Li, W.; Li, Y. Agonists and inhibitors of the STING pathway: Potential agents for immunotherapy. Med. Res. Rev. 2020, 40, 1117–1141. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.; Aloni, Y.; Weinhouse, C.; Michaeli, D.; Weinberger-Ohana, P.; Meyer, R.; Benziman, M. An unusual guanyl oligonucleotide regulates cellulose synthesis in Acetobacter xylinum. FEBS Lett. 1985, 186, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.; Aloni, Y.; Weinhouse, H.; Michaeli, D.; Weinberger-Ohana, P.; Mayer, R.; Benziman, M. Control of cellulose synthesis Acetobacter xylinum. A unique guanyl oligonucleotide is the immediate activator of the cellulose synthase. Carbohydr. Res. 1986, 149, 101–117. [Google Scholar] [CrossRef]
- Ross, P.; Weinhouse, H.; Aloni, Y.; Michaeli, D.; Weinberger-Ohana, P.; Mayer, R.; Braun, S.; de Vroom, E.; van der Marel, G.A.; van Boom, J.H.; et al. Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature 1987, 325, 279–281. [Google Scholar] [CrossRef]
- Egli, M.; Gessner, R.V.; Williams, L.D.; Quigley, G.J.; van der Marel, G.A.; van Boom, J.H.; Rich, A.; Frederick, C.A. Atomic-resolution structure of the cellulose synthase regulator cyclic diguanylic acid. Proc. Natl. Acad. Sci. USA 1990, 87, 3235–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, G.; Hartung, S.; Büttner, K.; Hopfner, K.-P. Structural Biochemistry of a Bacterial Checkpoint Protein Reveals Diadenylate Cyclase Activity Regulated by DNA Recombination Intermediates. Mol. Cell 2008, 30, 167–178. [Google Scholar] [CrossRef]
- Stülke, J.; Krüger, L. Cyclic di-AMP Signaling in Bacteria. Annu. Rev. Microbiol. 2020, 74, 159–179. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Cai, X.; Ma, H.; Zhu, L.; Zhang, Y.; Chou, S.-H.; Galperin, M.Y.; He, J. A decade of research on the second messenger c-di-AMP. FEMS Microbiol. Rev. 2020, 44, 701–724. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Bogard, R.W.; Young, T.S.; Mekalanos, J.J. Coordinated Regulation of Accessory Genetic Elements Produces Cyclic Di-Nucleotides for V. cholerae Virulence. Cell 2012, 149, 358–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP Synthase Is a Cytosolic DNA Sensor That Activates the Type I Interferon Pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Sun, L.; Chen, X.; Du, F.; Shi, H.; Chen, C.; Chen, Z.J. Cyclic GMP-AMP Is an Endogenous Second Messenger in Innate Immune Signaling by Cytosolic DNA. Science 2013, 339, 826–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Shi, H.; Wu, J.; Zhang, X.; Sun, L.; Chen, C.; Chen, Z.J. Cyclic GMP-AMP containing mixed phosphodiester linkages is an endogenous high-affinity ligand for STING. Mol. Cell. 2013, 51, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Dahlstrom, K.M.; O’Toole, G.A. A Symphony of Cyclases: Specificity in Diguanylate Cyclase Signaling. Annu. Rev. Microbiol. 2017, 71, 179–195. [Google Scholar] [CrossRef]
- Ross, P.; Mayer, R.; Weinhouse, H.; Amikam, D.; Huggirat, Y.; Benziman, M.; de Vroom, E.; Fidder, A.; de Paus, P.; Sliedregt, L.A. The cyclic diguanylic acid regulatory system of cellulose synthesis in Acetobacter xylinum. Chemical synthesis and biological activity of cyclic nucleotide dimer, trimer, and phosphothioate derivatives. J. Biol. Chem. 1990, 265, 18933–18943. [Google Scholar] [CrossRef]
- Amikam, D.; Galperin, M. PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics 2005, 22, 3–6. [Google Scholar] [CrossRef]
- Ryjenkov, D.A.; Simm, R.; Romling, U.; Gomelsky, M. The PilZ domain is a receptor for the second messenger c-di-GMP: The PilZ domain protein YcgR controls motility in enterobacteria. J. Biol. Chem. 2006, 281, 30310–30314. [Google Scholar] [CrossRef] [Green Version]
- Alm, R.A.; Bodero, A.J.; Free, P.D.; Mattick, J. Identification of a novel gene, pilZ, essential for type 4 fimbrial biogenesis in Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christen, M.; Christen, B.; Allan, M.G.; Folcher, M.; Jenö, P.; Grzesiek, S.; Jenal, U. DgrA is a member of a new family of cyclic diguanosine monophosphate receptors and controls flagellar motor function in Caulobacter crescentus. Proc. Natl. Acad. Sci. USA 2007, 104, 4112–4117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Granero, F.; Navazo, A.; Barahona, E.; Redondo-Nieto, M.; de Heredia, E.G.; Baena, I.; Martín-Martín, I.; Rivilla, R.; Martín, M. Identification of flgZ as a Flagellar Gene Encoding a PilZ Domain Protein That Regulates Swimming Motility and Biofilm Formation in Pseudomonas. PLoS ONE 2014, 9, e87608. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.T.; Tamayo, R.; Tischler, A.; Camilli, A. PilZ Domain Proteins Bind Cyclic Diguanylate and Regulate Diverse Processes in Vibrio cholerae. J. Biol. Chem. 2007, 282, 12860–12870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chen, J.-H.; Hao, Y.; Nair, S.K. Structures of the PelD Cyclic Diguanylate Effector Involved in Pellicle Formation in Pseudomonas aeruginosa PAO1. J. Biol. Chem. 2012, 287, 30191–30204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell, P.D.; Monds, R.D.; O’Toole, G.A. LapD is a bis-(3′,5′)-cyclic dimeric GMP-binding protein that regulates surface attachment by Pseudomonas fluorescens Pf0-1. Proc. Natl. Acad. Sci. USA 2009, 106, 3461–3466. [Google Scholar] [CrossRef] [Green Version]
- Hickman, J.W.; Harwood, C.S. Identification of FleQ from Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol. Microbiol. 2008, 69, 376–389. [Google Scholar] [CrossRef] [Green Version]
- Fazli, M.M.; O’Connell, A.; Nilsson, M.; Niehaus, K.; Dow, J.M.; Givskov, M.; Ryan, R.P.; Tolker-Nielsen, T. The CRP/FNR family protein Bcam1349 is a c-di-GMP effector that regulates biofilm formation in the respiratory pathogen Burkholderia cenocepacia. Mol. Microbiol. 2011, 82, 327–341. [Google Scholar] [CrossRef]
- Smith, K.D.; Shanahan, C.A.; Moore, E.L.; Simon, A.C.; Strobel, S.A. Structural basis of differential ligand recognition by two classes of bis-(3′-5′)-cyclic dimeric guanosine monophosphate-binding riboswitches. Proc. Natl. Acad. Sci. USA 2011, 108, 7757–7762. [Google Scholar] [CrossRef] [Green Version]
- Sudarsan, N.; Lee, E.R.; Weinberg, Z.; Moy, R.; Kim, J.N.; Link, K.H.; Breaker, R.R. Riboswitches in Eubacteria Sense the Second Messenger Cyclic Di-GMP. Science 2008, 321, 411–413. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.R.; Baker, J.L.; Weinberg, Z.; Sudarsan, N.; Breaker, R.R. An Allosteric Self-Splicing Ribozyme Triggered by a Bacterial Second Messenger. Science 2010, 329, 845–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.B.; Petrova, O.E.; Sauer, K. Extraction and Quantification of Cyclic Di-GMP from P. aeruginosa. Bio Protoc. 2013, 3, e828. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Sinn, M.; Stifel, J.; Heiler, A.C.; Sommershof, A.; Hartig, J.S. Synthesis of All Possible Canonical (3′-5′-Linked) Cyclic Di-nucleotides and Evaluation of Riboswitch Interactions and Immune-Stimulatory Effects. J. Am. Chem. Soc. 2017, 139, 16154–16160. [Google Scholar] [CrossRef] [Green Version]
- Rao, F.; Pasunooti, S.; Ng, Y.; Zhuo, W.; Lim, L.; Liu, A.W.; Liang, Z.-X. Enzymatic synthesis of c-di-GMP using a thermophilic diguanylate cyclase. Anal. Biochem. 2009, 389, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Korovashkina, A.S.; Rymko, A.N.; Kvach, S.V.; Zinchenko, A. Enzymatic synthesis of c-di-GMP using inclusion bodies of Ther-motoga maritima full-length diguanylate cyclase. J. Biotechnol. 2012, 164, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Shchokolova, A.S.; Rymko, A.N.; Kvach, S.V.; Shabunya, P.S.; Fatykhava, S.A.; Zinchenko, A.I. Enzymatic synthesis of 2′-ara and 2′-deoxy analogues of c-di-GMP. Nucleosides Nucleotides Nucleic Acids 2015, 34, 416–423. [Google Scholar] [CrossRef]
- Launer-Felty, K.D.; Strobel, S.A. Enzymatic synthesis of cyclic dinucleotide analogs by a promiscuous cyclic-AMP-GMP syn-thetase and analysis of cyclic dinucleotide responsive riboswitches. Nucleic Acids Res. 2018, 46, 2765–2776. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Nikel, P.; Andexer, J.; Lütz, S.; Rosenthal, K. A Multi-Enzyme Cascade Reaction for the Production of 2′3′-cGAMP. Biomolecules 2021, 11, 590. [Google Scholar] [CrossRef]
- Li, L.; Yin, Q.; Kuss, P.; Maliga, Z.; Millan, J.L.; Wu, H.; Mitchison, T.J. Hydrolysis of 2′3′-cGAMP by ENPP1 and design of nonhydrolyzable analogs. Nat. Chem. Biol. 2014, 10, 1043–1048. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, K.; Becker, M.; Rolf, J.; Siedentop, R.; Hillen, M.; Nett, M.; Lütz, S. Catalytic promiscuity of cGAS: A facile enzymatic synthesis of 2′-3′-Linked cyclic dinucleotides. Chembiochem Eur. J. Chem. Biol. 2020, 21, 3225–3228. [Google Scholar] [CrossRef]
- Dubensky, J.T.W.; Kanne, D.B.; Leong, M.L. Rationale, progress and development of vaccines utilizing STING-activating cyclic dinucleotide adjuvants. Ther. Adv. Vaccines 2013, 1, 131–143. [Google Scholar] [CrossRef]
- Parvatiyar, K.; Zhang, Z.; Teles, R.; Ouyang, S.; Jiang, Y.; Iyer, S.S.; Zaver, S.A.; Schenk, M.; Zeng, S.; Zhong, W.; et al. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat. Immunol. 2012, 13, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- McFarland, A.P.; Luo, S.; Ahmed-Qadri, F.; Zuck, M.; Thayer, E.F.; Goo, Y.A.; Hybiske, K.; Tong, L.; Woodward, J.J. Sensing of Bacterial Cyclic Dinucleotides by the Oxidoreductase RECON Promotes NF-kappaB Activation and Shapes a Proinflammatory Antibacterial State. Immunity 2017, 46, 433–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaauboer, S.M.; Gabrielle, V.D.; Jin, L. MPYS/STING-Mediated TNF-α, Not Type I IFN, Is Essential for the Mucosal Adjuvant Activity of (3′–5′)-Cyclic-Di-Guanosine-Monophosphate In Vivo. J. Immunol. 2013, 192, 492–502. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.; Whiteley, A.T.; Gwilt, K.B.; Lencer, W.I.; Mekalanos, J.J.; Thiagarajah, J.R. Extracellular cyclic dinucleotides induce polarized responses in barrier epithelial cells by adenosine signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 27502–27508. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Li, X.; Li, P. The mechanism of double-stranded DNA sensing through the cGAS-STING pathway. Cytokine Growth Factor Rev. 2014, 25, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škrnjug, I.; Rueckert, C.; Libanova, R.; Lienenklaus, S.; Weiss, S.; Guzmán, C. The Mucosal Adjuvant Cyclic di-AMP Exerts Immune Stimulatory Effects on Dendritic Cells and Macrophages. PLoS ONE 2014, 9, e95728. [Google Scholar] [CrossRef]
- Karaolis, D.K.R.; Means, T.K.; Yang, D.; Takahashi, M.; Yoshimura, T.; Muraille, E.; Philpott, D.; Schroeder, J.T.; Hyodo, M.; Hayakawa, Y.; et al. Bacterial c-di-GMP Is an Immunostimulatory Molecule. J. Immunol. 2007, 178, 2171–2181. [Google Scholar] [CrossRef] [Green Version]
- Ebensen, T.; Libanova, R.; Schulze, K.; Yevsa, T.; Morr, M.; Guzman, C.A. Bis-(3′,5′)-cyclic dimeric adenosine monophosphate: Strong Th1/Th2/Th17 promoting mucosal adjuvant. Vaccine 2011, 29, 5210–5220. [Google Scholar] [CrossRef]
- Jin, L.; Hill, K.K.; Filak, H.; Mogan, J.; Knowles, H.; Zhang, B.; Perraud, A.-L.; Cambier, J.C.; Lenz, L.L. MPYS Is Required for IFN Response Factor 3 Activation and Type I IFN Production in the Response of Cultured Phagocytes to Bacterial Second Messengers Cyclic-di-AMP and Cyclic-di-GMP. J. Immunol. 2011, 187, 2595–2601. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.R.; Blomquist, C.M.; Henare, K.L.; Jirik, F.R. Stimulator of interferon genes (STING) activation exacerbates experimental colitis in mice. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Darling, R.; Senapati, S.; Kelly, S.M.; Kohut, M.L.; Narasimhan, B.; Wannemuehler, M.J. STING pathway stimulation results in a differentially activated innate immune phenotype associated with low nitric oxide and enhanced antibody titers in young and aged mice. Vaccine 2019, 37, 2721–2730. [Google Scholar] [CrossRef]
- Mansouri, S.; Katikaneni, D.S.; Gogoi, H.; Jin, L. Monocyte-Derived Dendritic Cells (moDCs) Differentiate into Bcl6+ Mature moDCs to Promote Cyclic di-GMP Vaccine Adjuvant–Induced Memory TH Cells in the Lung. J. Immunol. 2021, 206, 2233–2245. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Sater, A.A.; Tattoli, I.; Jin, L.; Grajkowski, A.; Levi, A.; Koller, B.H.; Allen, I.C.; Beaucage, S.L.; Fitzgerald, K.A.; Ting, J.P.Y.; et al. Cyclic-di-GMP and cyclic-di-AMP activate the NLRP3 inflammasome. EMBO Rep. 2013, 14, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.M.; Crute, B.W.; Cambier, J.C.; Getahun, A. B Cell–Intrinsic STING Signaling Triggers Cell Activation, Synergizes with B Cell Receptor Signals, and Promotes Antibody Responses. J. Immunol. 2018, 201, 2641–2653. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, T.; Unno, M.; Kobayashi, W.; Yoneda, N.; Matsuda, S.; Ikeda, K.; Hoshii, T.; Hirao, A.; Miyake, K.; Barber, G.N.; et al. Reciprocal regulation of STING and TCR signaling by mTORC1 for T-cell activation and function. Life Sci. Alliance 2019, 2, e201800282. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, A.; Papagno, L.; Nicoli, F.; Kanuma, T.; Kuse, N.; Cabral-Piccin, M.P.; Rochereau, N.; Gostick, E.; Lioux, T.; Perouzel, E.; et al. The STING ligand cGAMP potentiates the efficacy of vaccine-induced CD8+ T cells. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Kuse, N.; Sun, X.; Akahoshi, T.; Lissina, A.; Yamamoto, T.; Appay, V.; Takiguchi, M. Priming of HIV-1-specific CD8+ T cells with strong functional properties from naïve T cells. EBioMedicine 2019, 42, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Kanuma, T.; Takahama, S.; Okamura, T.; Moriishi, E.; Ishii, K.J.; Terahara, K.; Yasutomi, Y. STING agonists activate latently infected cells and enhance SIV-specific responses ex vivo in naturally SIV controlled cynomolgus macaques. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Elmanfi, S.; Zhou, J.; Sintim, H.O.; Könönen, E.; Gürsoy, M.; Gürsoy, U.K. Regulation of gingival epithelial cytokine response by bacterial cyclic dinucleotides. J. Oral Microbiol. 2018, 11, 1538927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; KuoLee, R.; Tram, K.; Qiu, H.; Zhang, J.; Patel, G.B.; Chen, W. 3′,5′-Cyclic diguanylic acid elicits mucosal immunity against bacterial infection. Biochem. Biophys. Res. Commun. 2009, 387, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Ogunniyi, A.D.; Paton, J.C.; Kirby, A.C.; McCullers, J.A.; Cook, J.; Hyodo, M.; Hayakawa, Y.; Karaolis, D.K. c-di-GMP is an effective immunomodulator and vaccine adjuvant against pneumococcal infection. Vaccine 2008, 26, 4676–4685. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; KuoLee, R.; Harris, G.; Tram, K.; Yan, H.; Chen, W. c-di-GMP protects against intranasal Acinetobacter baumannii infection in mice by chemokine induction and enhanced neutrophil recruitment. Int. Immunopharmacol. 2011, 11, 1378–1383. [Google Scholar] [CrossRef]
- Skouboe, M.K.; Knudsen, A.; Reinert, L.; Boularan, C.; Lioux, T.; Pérouzel, E.; Thomsen, M.K.; Paludan, S.R. STING agonists enable antiviral cross-talk between human cells and confer protection against genital herpes in mice. PLoS Pathog. 2018, 14, e1006976. [Google Scholar] [CrossRef] [PubMed]
- Blaauboer, S.M.; Mansouri, S.; Tucker, H.R.; Wang, H.L.; Gabrielle, V.D.; Jin, L. The mucosal adjuvant cyclic di-GMP enhances antigen uptake and selectively activates pinocytosis-efficient cells in vivo. eLife 2015, 4, e06670. [Google Scholar] [CrossRef]
- Elahi, S.; van Kessel, J.; Kiros, T.G.; Strom, S.; Hayakawa, Y.; Hyodo, M.; Babiuk, L.A.; Gerdts, V. c-di-GMP Enhances Protective Innate Immunity in a Murine Model of Pertussis. PLoS ONE 2014, 9, e109778. [Google Scholar] [CrossRef] [Green Version]
- Fatima, M.; Rempel, H.; Kuang, X.T.; Allen, K.J.; Cheng, K.M.; Malouin, F.; Diarra, M.S. Effect of 3’,5’-cyclic diguanylic acid in a broiler Clostridium perfringens infection model. Poult. Sci. 2013, 92, 2644–2650. [Google Scholar] [CrossRef]
- Karaolis, D.K.R.; Newstead, M.W.; Zeng, X.; Hyodo, M.; Hayakawa, Y.; Bhan, U.; Liang, H.; Standiford, T.J. Cyclic Di-GMP Stimulates Protective Innate Immunity in Bacterial Pneumonia. Infect. Immun. 2007, 75, 4942–4950. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.M.; Uchimura, T.; Gallovic, M.D.; Thamilarasan, M.; Chou, W.-C.; Gibson, S.A.; Deng, M.; Tam, J.W.; Batty, C.J.; Williams, J.; et al. STING Agonist Mitigates Experimental Autoimmune Encephalomyelitis by Stimulating Type I IFN–Dependent and –Independent Immune-Regulatory Pathways. J. Immunol. 2021, 206, 2015–2028. [Google Scholar] [CrossRef] [PubMed]
- Ohkuri, T.; Ghosh, A.; Kosaka, A.; Zhu, J.; Ikeura, M.; David, M.; Watkins, S.C.; Sarkar, S.N.; Okada, H. STING contributes to antiglioma immunity via triggering type I IFN signals in the tumor microenvironment. Cancer Immunol. Res. 2014, 2, 1199–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales, L.; Glickman, L.H.; McWhirter, S.M.; Kanne, D.B.; Sivick, K.E.; Katibah, G.E.; Woo, S.-R.; Lemmens, E.; Banda, T.; Leong, J.J.; et al. Direct Activation of STING in the Tumor Microenvironment Leads to Potent and Systemic Tumor Regression and Immunity. Cell Rep. 2015, 11, 1018–1030. [Google Scholar] [CrossRef] [Green Version]
- Gadkaree, S.K.; Fu, J.; Sen, R.; Korrer, M.J.; Allen, C.; Kim, Y.J. Induction of tumor regression by intratumoral STING agonists combined with anti-programmed death-L1 blocking antibody in a preclinical squamous cell carcinoma model. Head Neck 2017, 39, 1086–1094. [Google Scholar] [CrossRef]
- Moore, E.; Clavijo, P.E.; Davis, R.; Cash, H.; van Waes, C.; Kim, Y.; Allen, C. Established T Cell–Inflamed Tumors Rejected after Adaptive Resistance Was Reversed by Combination STING Activation and PD-1 Pathway Blockade. Cancer Immunol. Res. 2016, 4, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Wehbe, M.; Wang-Bishop, L.; Becker, K.W.; Shae, D.; Baljon, J.J.; He, X.; Christov, P.; Boyd, K.L.; Balko, J.M.; Wilson, J.T. Nanoparticle delivery improves the pharmacokinetic properties of cyclic dinucleotide STING agonists to open a therapeutic window for intravenous administration. J. Control. Release 2021, 330, 1118–1129. [Google Scholar] [CrossRef]
- Wang-Bishop, L.; Wehbe, M.; Shae, D.; James, J.; Hacker, B.C.; Garland, K.; Chistov, P.P.; Rafat, M.; Balko, J.M.; Wilson, J.T. Potent STING activation stimulates immunogenic cell death to enhance antitumor immunity in neuroblastoma. J. Immunother. Cancer 2019, 8, e000282. [Google Scholar] [CrossRef] [Green Version]
- Foote, J.B.; Kok, M.; Leatherman, J.M.; Armstrong, T.D.; Marcinkowski, B.; Ojalvo, L.S.; Kanne, D.B.; Jaffee, E.; Dubensky, T.W.; Emens, L.A. A STING Agonist Given with OX40 Receptor and PD-L1 Modulators Primes Immunity and Reduces Tumor Growth in Tolerized Mice. Cancer Immunol. Res. 2017, 5, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielecki, P.A.; Lorkowski, M.E.; Becicka, W.M.; Atukorale, P.U.; Moon, T.J.; Zhang, Y.; Wiese, M.; Covarrubias, G.; Ravichandran, S.; Karathanasis, E. Immunostimulatory silica nanoparticle boosts innate immunity in brain tumors. Nanoscale Horiz. 2021, 6, 156–167. [Google Scholar] [CrossRef]
- Chen, Y.P.; Xu, L.; Tang, T.W.; Chen, C.H.; Zheng, Q.H.; Liu, T.P.; Mou, C.Y.; Wu, C.H.; Wu, S.H. STING Activator c-di-GMP-Loaded Mesoporous Silica Nano-particles Enhance Immunotherapy Against Breast Cancer. ACS Appl. Mater. Interfaces 2020, 12, 56741–56752. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Yu, C.; Xi, J.; Reyes, J.; Mao, G.; Wei, W.Z.; Liu, H. Induction of necrotic cell death and activation of STING in the tumor mi-croenvironment via cationic silica nanoparticles leading to enhanced antitumor immunity. Nanoscale 2018, 10, 9311–9319. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.R.; Sen, R.; Sunshine, J.; Pardoll, D.M.; Green, J.J.; Kim, Y.J. Biodegradable STING agonist nanoparticles for enhanced cancer immunotherapy. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Ager, C.R.; Reilley, M.J.; Nicholas, C.; Bartkowiak, T.; Jaiswal, A.; Curran, M.A. Intratumoral STING Activation with T-cell Checkpoint Modulation Generates Systemic Antitumor Immunity. Cancer Immunol. Res. 2017, 5, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, N.; Watkins-Schulz, R.; Junkins, R.D.; David, C.N.; Johnson, B.M.; Montgomery, S.A.; Peine, K.J.; Darr, D.B.; Yuan, H.; McKinnon, K.P.; et al. A nanoparticle-incorporated STING activator enhances antitumor immunity in PD-L1–insensitive models of triple-negative breast cancer. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.T.; Moffett, H.; Stephan, S.B.; Opel, C.F.; Dumigan, A.; Jiang, X.; Pillarisetty, V.G.; Pillai, S.P.S.; Wittrup, K.D.; Stephan, M. Biopolymers codelivering engineered T cells and STING agonists can eliminate heterogeneous tumors. J. Clin. Investig. 2017, 127, 2176–2191. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; An, X.; Zhang, X.; Qiao, Y.; Zheng, T.; Li, X. STING: A master regulator in the cancer-immunity cycle. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Atukorale, P.U.; Raghunathan, S.P.; Raguveer, V.; Moon, T.J.; Zheng, C.; Bielecki, P.A.; Wiese, M.L.; Goldberg, A.L.; Covarrubias, G.; Hoimes, C.J.; et al. Nanoparticle Encapsulation of Synergistic Immune Agonists Enables Systemic Codelivery to Tumor Sites and IFNbeta-Driven Antitumor Immunity. Cancer Res. 2019, 79, 5394–5406. [Google Scholar] [CrossRef] [Green Version]
- Temizoz, B.; Kuroda, E.; Ohata, K.; Jounai, N.; Ozasa, K.; Kobiyama, K.; Aoshi, T.; Ishii, K.J. TLR9 and STING agonists synergistically induce innate and adaptive type-II IFN. Eur. J. Immunol. 2015, 45, 1159–1169. [Google Scholar] [CrossRef]
- Francica, B.J.; Ghasemzadeh, A.; Desbien, A.L.; Theodros, D.; Sivick, K.E.; Reiner, G.L.; Glickman, L.H.; Marciscano, A.E.; Sharabi, A.B.; Leong, M.L.; et al. TNFalpha and Radioresistant Stromal Cells Are Essential for Therapeutic Efficacy of Cyclic Dinucleotide STING Agonists in Nonimmunogenic Tumors. Cancer Immunol. Res. 2018, 6, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Qi, J.; Zhao, Q.; Wu, Q.-N.; Wei, D.-L.; Wei, X.-L.; Liu, J.; Chen, J.; Zeng, Z.-L.; Ju, H.-Q.; et al. Targeting the STING pathway in tumor-associated macrophages regulates innate immune sensing of gastric cancer cells. Theranostics 2020, 10, 498–515. [Google Scholar] [CrossRef]
- Ohkuri, T.; Kosaka, A.; Ishibashi, K.; Kumai, T.; Hirata, Y.; Ohara, K.; Nagato, T.; Oikawa, K.; Aoki, N.; Harabuchi, Y.; et al. Intratumoral administration of cGAMP transiently ac-cumulates potent macrophages for anti-tumor immunity at a mouse tumor site. Cancer Immunol. Immunother. 2017, 66, 705–716. [Google Scholar] [CrossRef]
- Nicolai, C.J.; Wolf, N.; Chang, I.-C.; Kirn, G.; Marcus, A.; Ndubaku, C.O.; McWhirter, S.M.; Raulet, D.H. NK cells mediate clearance of CD8+ T cell–resistant tumors in response to STING agonists. Sci. Immunol. 2020, 5, eaaz2738. [Google Scholar] [CrossRef] [PubMed]
- Ebensen, T.; Schulze, K.; Riese, P.; Link, C.; Morr, M.; Guzmán, C.A. The bacterial second messenger cyclic diGMP exhibits potent adjuvant properties. Vaccine 2007, 25, 1464–1469. [Google Scholar] [CrossRef]
- Ebensen, T.; Schulze, K.; Riese, P.; Morr, M.; Guzmán, C.A. The Bacterial Second Messenger cdiGMP Exhibits Promising Activity as a Mucosal Adjuvant. Clin. Vaccine Immunol. 2007, 14, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Libanova, R.; Ebensen, T.; Schulze, K.; Bruhn, D.; Nörder, M.; Yevsa, T.; Morr, M.; Guzmán, C.A. The member of the cyclic di-nucleotide family bis-(3′, 5′)-cyclic dimeric inosine monophosphate exerts potent activity as mucosal adjuvant. Vaccine 2010, 28, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Škrnjug, I.; Guzmán, C.; Ruecker, C. Cyclic GMP-AMP Displays Mucosal Adjuvant Activity in Mice. PLoS ONE 2014, 9, e110150. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.M.; Forrest, G.; Wisniewski, T.; Porter, G.; Freed, D.C.; DeMartino, J.A.; Zaller, D.M.; Guo, Z.; Leone, J.; Fu, T.-M.; et al. Evidence for cyclic diguanylate as a vaccine adjuvant with novel immunostimulatory activities. Cell. Immunol. 2012, 278, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Volckmar, J.; Knop, L.; Stegemann-Koniszewski, S.; Schulze, K.; Ebensen, T.; Guzman, C.A.; Bruder, D. The STING activator c-di-AMP exerts superior adjuvant properties than the formulation poly(I:C)/CpG after subcutaneous vaccination with soluble protein antigen or DEC-205-mediated antigen targeting to dendritic cells. Vaccine 2019, 37, 4963–4974. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, S.; Alpdundar, E.; Gungor, B.; Kahraman, T.; Bayyurt, B.; Gursel, I.; Gursel, M. Enhanced immunostimulatory activity of cyclic dinucleotides on mouse cells when complexed with a cell-penetrating peptide or combined with CpG. Eur. J. Immunol. 2015, 45, 1170–1179. [Google Scholar] [CrossRef]
- Guo, J.; Wang, J.; Fan, J.; Zhang, Y.; Dong, W.; Chen, C.P. Distinct Dynamic and Conformational Features of Human STING in Re-sponse to 2′3′-cGAMP and c-di-GMP. Chembiochem Eur. J. Chem. Biol. 2019, 20, 1838–1847. [Google Scholar]
- Yan, H.; Wang, X.; KuoLee, R.; Chen, W. Synthesis and immunostimulatory properties of the phosphorothioate analogues of cdiGMP. Bioorganic Med. Chem. Lett. 2008, 18, 5631–5634. [Google Scholar] [CrossRef]
- Li, J.; Lee, R.K.; Chen, W.; Yan, H. 2′-Fluoro-c-di-GMP as an oral vaccine adjuvant. RSC Adv. 2019, 9, 41481–41489. [Google Scholar] [CrossRef] [Green Version]
- Flores-Valdez, M.A.; Aceves-Sánchez, M.D.J.; Montero-Pérez, S.A.; Sánchez-López, A.D.; Gutiérrez-Pabello, J.A.; Hernández-Pando, R. Vaccination of mice with recombinant bacille Calmette-Guérin harboring Rv1357c protects similarly to native BCG. Int. J. Tuberc. Lung Dis. 2012, 16, 774–776. [Google Scholar] [CrossRef]
- Pedroza-Roldan, C.; Guapillo, C.; Barrios-Payán, J.; Mata-Espinosa, D.; Aceves-Sánchez, M.D.J.; Marquina-Castillo, B.; Hernández-Pando, R.; Flores-Valdez, M.A. The BCGΔBCG1419c strain, which produces more pellicle in vitro, improves control of chronic tuberculosis in vivo. Vaccine 2016, 34, 4763–4770. [Google Scholar] [CrossRef] [PubMed]
- Ning, H.; Wang, L.; Zhou, J.; Lu, Y.; Kang, J.; Ding, T.; Shen, L.; Xu, Z.; Bai, Y. Recombinant BCG With Bacterial Signaling Molecule Cyclic di-AMP as Endogenous Adjuvant Induces Elevated Immune Responses After Mycobacterium tuberculosis Infection. Front. Immunol. 2019, 10, 1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, R.J.; Dey, B.; Singh, A.K.; Praharaj, M.; Bishai, W. Bacillus Calmette-Guérin Overexpressing an Endogenous Stimulator of Interferon Genes Agonist Provides Enhanced Protection Against Pulmonary Tuberculosis. J. Infect. Dis. 2020, 221, 1048–1056. [Google Scholar] [CrossRef] [Green Version]
- Madhun, A.S.; Haaheim, L.R.; Nostbakken, J.K.; Ebensen, T.; Chichester, J.; Yusibov, V.; Guzmán, C.A.; Cox, R.J. Intranasal c-di-GMP-adjuvanted plant-derived H5 influenza vaccine induces multifunctional Th1 CD4+ cells and strong mucosal and systemic antibody re-sponses in mice. Vaccine 2011, 29, 4973–4982. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, G.K.; Ebensen, T.; Gjeraker, I.H.; Svindland, S.; Bredholt, G.; Guzmán, C.A.; Cox, R.J. Evaluation of the Sublingual Route for Administration of Influenza H5N1 Virosomes in Combination with the Bacterial Second Messenger c-di-GMP. PLoS ONE 2011, 6, e26973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svindland, S.C.; Pedersen, G.; Pathirana, R.D.; Bredholt, G.; Nøstbakken, J.K.; Jul-Larsen, Å.; Guzmán, C.; Montomoli, E.; Lapini, G.; Piccirella, S.; et al. A study of Chitosan and c-di-GMP as mucosal adjuvants for intranasal influenza H5N1 vaccine. Influenza Other Respir. Viruses 2012, 7, 1181–1193. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, V.; Chichester, J.A.; Ebensen, T.; Schwarz, K.; Hartman, C.E.; Shoji, Y.; Guzmán, C.A.; Yusibov, V.; Sewald, K.; Braun, A. A new adjuvanted nanoparticle-based H1N1 influenza vaccine induced antigen-specific local mucosal and systemic immune responses after administration into the lung. Vaccine 2014, 32, 3216–3222. [Google Scholar] [CrossRef]
- Major, D.; Chichester, J.A.; Pathirana, R.D.; Guilfoyle, K.; Shoji, Y.; Guzmán, C.; Yusibov, V.; Cox, R.J. Intranasal vaccination with a plant-derived H5 HA vaccine protects mice and ferrets against highly pathogenic avian influenza virus challenge. Hum. Vaccines Immunother. 2015, 11, 1235–1243. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Lee, J.-H.; Jeong, S.D.; Noh, J.-Y.; Lee, H.W.; Song, C.-S.; Kim, Y.-C. C-di-GMP with influenza vaccine showed enhanced and shifted immune responses in microneedle vaccination in the skin. Drug Deliv. Transl. Res. 2020, 10, 815–825. [Google Scholar] [CrossRef]
- Allen, A.C.; Wilk, M.; Misiak, A.; Borkner, L.; Murphy, D.; Mills, K.H.G. Sustained protective immunity against Bordetella pertussis nasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting TRM cells. Mucosal Immunol. 2018, 11, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.-L.; Narita, K.; Hyodo, M.; Hayakawa, Y.; Nakane, A.; Karaolis, D.K. c-di-GMP as a vaccine adjuvant enhances protection against systemic methicillin-resistant Staphylococcus aureus (MRSA) infection. Vaccine 2009, 27, 4867–4873. [Google Scholar] [CrossRef] [PubMed]
- Latanova, A.A.; Petkov, S.; Kilpelainen, A.; Jansons, J.; Latyshev, O.E.; Kuzmenko, Y.V.; Hinkula, J.; Abakumov, M.A.; Valuev-Eliston, V.T.; Gomelsy, M.; et al. Codon optimization and improved delivery/immunization regimen enhance the immune response against wild-type and drug-resistant HIV-1 reverse tran-scriptase, preserving its Th2-polarity. Sci. Rep. 2018, 8, 8078. [Google Scholar] [CrossRef]
- Lee, M.J.; Jo, H.; Shin, S.H.; Kim, S.M.; Kim, B.; Shim, H.S.; Park, J.H. Mincle and STING-Stimulating Adjuvants Elicit Robust Cellular Immunity and Drive Long-Lasting Memory Responses in a Foot-and-Mouth Disease Vaccine. Front. Immunol. 2019, 10, 2509. [Google Scholar] [CrossRef]
- Azegami, T.; Yuki, Y.; Sawada, S.; Mejima, M.; Ishige, K.; Akiyoshi, K.; Itoh, H.; Kiyono, H. Nanogel-based nasal ghrelin vaccine prevents obesity. Mucosal Immunol. 2017, 10, 1351–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azegami, T.; Yuki, Y.; Hayashi, K.; Hishikawa, A.; Sawada, S.-I.; Ishige, K.; Akiyoshi, K.; Kiyono, H.; Itoh, H. Intranasal vaccination against angiotensin II type 1 receptor and pneumococcal surface protein A attenuates hypertension and pneumococcal infection in rodents. J. Hypertens. 2018, 36, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Ebensen, T.; Debarry, J.; Pedersen, G.K.; Blazejewska, P.; Weissmann, S.; Schulze, K.; McCullough, K.C.; Cox, R.J.; Guzman, C.A. Mucosal Administration of Cy-cle-Di-Nucleotide-Adjuvanted Virosomes Efficiently Induces Protection against Influenza H5N1 in Mice. Front. Immunol. 2017, 8, 1223. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.; Ebensen, T.; Schulze, K.; Cargnelutti, D.E.; Blazejewska, P.; Scodeller, E.A.; Guzmán, C.A. Intranasal Delivery of Influenza rNP Adjuvanted with c-di-AMP Induces Strong Humoral and Cellular Immune Responses and Provides Protection against Virus Challenge. PLoS ONE 2014, 9, e104824. [Google Scholar] [CrossRef] [PubMed]
- Ning, H.; Zhang, W.; Kang, J.; Ding, T.; Liang, X.; Lu, Y.; Guo, C.; Sun, W.; Wang, H.; Bai, Y.; et al. Subunit Vaccine ESAT-6:c-di-AMP Delivered by Intranasal Route Elicits Immune Responses and Protects Against Mycobacterium tuberculosis Infection. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Schulze, K.; Ebensen, T.; Chandrudu, S.; Skwarczynski, M.; Toth, I.; Olive, C.; Guzman, C.A. Bivalent mucosal peptide vaccines administered using the LCP carrier system stimulate protective immune responses against Streptococcus pyogenes infection. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2463–2474. [Google Scholar] [CrossRef]
- Landi, A.; Law, J.; Hockman, D.; Logan, M.; Crawford, K.; Chen, C.; Kundu, J.; Ebensen, T.; Guzman, C.; Deschatelets, L.; et al. Superior immunogenicity of HCV envelope glycoproteins when adjuvanted with cyclic-di-AMP, a STING activator or archaeosomes. Vaccine 2017, 35, 6949–6956. [Google Scholar] [CrossRef] [PubMed]
- Munoz Gonzalez, F.; Sycz, G.; Alonso Paiva, I.M.; Linke, D.; Zorreguieta, A.; Baldi, P.C.; Ferero, M.C. The BtaF Adhesin Is Necessary for Full Virulence during Respiratory Infection by Brucella suis and Is a Novel Immunogen for Nasal Vaccination Against Brucella Infection. Front. Immunol. 2019, 10, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos, M.N.; Cazorla, S.I.; Schulze, K.; Ebensen, T.; Guzmán, C.A.; Malchiodi, E. Immunization with Tc52 or its amino terminal domain adjuvanted with c-di-AMP induces Th17+Th1 specific immune responses and confers protection against Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2017, 11, e0005300. [Google Scholar] [CrossRef] [PubMed]
- Sanchez Alberti, A.; Bivona, A.E.; Cerny, N.; Schulze, K.; Weissmann, S.; Ebensen, T.; Morales, C.; Padilla, A.M.; Cazorla, S.I.; Tarleton, R.L.; et al. Engineered trivalent immunogen adju-vanted with a STING agonist confers protection against Trypanosoma cruzi infection. NPJ Vaccines 2017, 2, 9. [Google Scholar] [CrossRef]
- Sanchez Alberti, A.; Bivona, A.E.; Matos, M.N.; Cerny, N.; Schulze, K.; Weissmann, S.; Ebensen, T.; Gonzalez, G.; Morales, C.; Cardoso, A.C.; et al. Mucosal Heterologous Prime/Boost Vaccination Induces Polyfunctional Systemic Immunity, Improving Protection Against Trypanosoma cruzi. Front. Immunol. 2020, 11, 128. [Google Scholar] [CrossRef] [Green Version]
- Matthijs, A.M.F.; Auray, G.; Jakob, V.; García-Nicolás, O.; Braun, R.O.; Keller, I.; Bruggman, R.; Devriendt, B.; Boyen, F.; Guzman, C.A.; et al. Systems Immunology Characterization of Novel Vaccine Formulations for Mycoplasma hyopneumoniae Bacterins. Front. Immunol. 2019, 10, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Liu, X.-P.; Xiong, F.-F.; Gao, F.-X.; Yi, Y.-L.; Zhang, M.; Chen, Z.; Tan, W.-S. Enhancing Immune Response and Heterosubtypic Protection Ability of Inactivated H7N9 Vaccine by Using STING Agonist as a Mucosal Adjuvant. Front. Immunol. 2019, 10, 2274. [Google Scholar] [CrossRef] [Green Version]
- Takaki, H.; Takashima, K.; Oshiumi, H.; Ainai, A.; Suzuki, T.; Hasegawa, H.; Matsumoto, M.; Seya, T. cGAMP Promotes Germinal Center Formation and Production of IgA in Nasal-Associated Lymphoid Tissue. Med. Sci. 2017, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, P.; Wu, M.X. Natural STING Agonist as an “Ideal” Adjuvant for Cutaneous Vaccination. J. Investig. Dermatol. 2016, 136, 2183–2191. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, P.; Yu, Y.; Fu, Y.; Jiang, H.; Lu, M.; Sun, Z.; Jiang, S.; Lu, L.; Wu, M.X. Pulmonary surfactant–biomimetic nanoparticles potentiate heterosubtypic influenza immunity. Science 2020, 367, eaau0810. [Google Scholar] [CrossRef]
- Vassilieva, E.V.; Li, S.; Korniychuk, H.; Taylor, D.M.; Wang, S.; Prausnitz, M.R.; Compans, R.W. cGAMP/Saponin Adjuvant Combination Improves Protective Response to Influenza Vaccination by Microneedle Patch in an Aged Mouse Model. Front. Immunol. 2021, 11, 583251. [Google Scholar] [CrossRef]
- Chen, J.; Zhong, Y.; Liu, Y.; Tang, C.; Zhang, Y.; Wei, B.; Chen, W.; Liu, M. Parenteral immunization with a cyclic guanosine monophos-phate-adenosine monophosphate (cGAMP) adjuvanted Helicobacter pylori vaccine induces protective immunity against H. pylori infection in mice. Hum. Vaccines Immunother. 2020, 16, 2849–2854. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, S.; Pan, B.; Guan, Z.; Yang, Z.; Duan, L.; Cai, H. A subunit vaccine based on rH-NS induces protection against Mycobac-terium tuberculosis infection by inducing the Th1 immune response and activating macrophages. Acta Biochim. Biophys. Sin. 2016, 48, 909–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorostkar, F.; Arashkia, A.; Roohvand, F.; Shoja, Z.; Navari, M.; Mashhadi Abolghasem Shirazi, M. Co-administration of 2’3’-cGAMP STING activator and CpG-C adjuvants with a mutated form of HPV 16 E7 protein leads to tumor growth inhibition in the mouse model. Infect. Agents Cancer 2021, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Kanbe, A.; Hara, A.; Ishikawa, T. Induction of humoral and cellular immune response to HBV vaccine can be up-regulated by STING ligand. Virology 2019, 531, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Borriello, F.; Pietrasanta, C.; Lai, J.C.Y.; Walsh, L.M.; Sharma, P.; O’Driscoll, D.N.; Ramirez, J.; Brightman, S.; Pugni, L.; Mosca, F.; et al. Identification and Characterization of Stimulator of Interferon Genes as a Robust Adjuvant Target for Early Life Immunization. Front. Immunol. 2017, 8, 1772. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Gallovic, M.D.; Tiet, P.; Ting, J.P.-Y.; Ainslie, K.; Bachelder, E.M. Investigation of tunable acetalated dextran microparticle platform to optimize M2e-based influenza vaccine efficacy. J. Control. Release 2018, 289, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Jee, J.; Kim, E.; Steiner, H.E.; Cormet-Boyaka, E.; Boyaka, P.N. Sublingual targeting of STING with 3′3′-cGAMP promotes systemic and mucosal immunity against anthrax toxins. Vaccine 2017, 35, 2511–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dis, E.; Sogi, K.M.; Rae, C.S.; Sivick, K.E.; Surh, N.H.; Leong, M.L. Faculty Opinions recommendation of STING-Activating Adjuvants Elicit a Th17 Immune Response and Protect against Mycobacterium tuberculosis Infection. Fac. Opin. 2018, 23, 1435–1447. [Google Scholar] [CrossRef] [Green Version]
- Wagner, D.A.; Kelly, S.M.; Petersen, A.C.; Peroutka-Bigus, N.; Darling, R.; Bellaire, B.H.; Wannemuehler, M.J.; Narasimhan, B. Single-dose combination nanovaccine induces both rapid and long-lived protection against pneumonic plague. Acta Biomater. 2019, 100, 326–337. [Google Scholar] [CrossRef]
- Latasa, C.; Echeverz, M.; Garcia, B.; Gil, C.; García-Ona, E.; Burgui, S.; Casares, N.; Hervas-Stubbs, S.; Lasarte, J.J.; Lasa, I.; et al. Evaluation of a Salmonella Strain Lacking the Secondary Messenger C-di-GMP and RpoS as a Live Oral Vaccine. PLoS ONE 2016, 11, e0161216. [Google Scholar] [CrossRef] [Green Version]
- Gil, C.; Latasa, C.; García-Ona, E.; Lázaro, I.; Labairu, J.; Echeverz, M.; Burgui, S.; García, B.; Lasa, I.; Solano, C. A DIVA vaccine strain lacking RpoS and the secondary messenger c-di-GMP for protection against salmonellosis in pigs. Veter. Res. 2020, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vassilieva, E.V.; Taylor, D.W.; Compans, R.W. Combination of STING Pathway Agonist with Saponin Is an Effective Adjuvant in Immunosenescent Mice. Front. Immunol. 2019, 10, 3006. [Google Scholar] [CrossRef] [Green Version]
- Gogoi, H.; Mansouri, S.; Katikaneni, D.S.; Jin, L. New MoDC-Targeting TNF Fusion Proteins Enhance Cyclic Di-GMP Vaccine Adjuvanticity in Middle-Aged and Aged Mice. Front. Immunol. 2020, 11, 1674. [Google Scholar] [CrossRef]
- Gravekamp, C.; Chandra, D. Targeting STING pathways for the treatment of cancer. OncoImmunology 2015, 4, e988463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, D.; Quispe-Tintaya, W.; Jahangir, A.; Asafu-Adjei, D.; Ramos, I.; Sintim, H.O.; Zhou, J.; Hayakawa, Y.; Karaolis, D.K.; Gravekamp, C. STING Ligand c-di-GMP Improves Cancer Vaccination against Metastatic Breast Cancer. Cancer Immunol. Res. 2014, 2, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-J.; Li, W.-H.; Chen, P.-G.; Zhang, B.-D.; Hu, H.-G.; Li, Q.-Q.; Zhao, L.; Chen, Y.-X.; Zhao, Y.-F.; Li, Y.-M. Targeting STING with cyclic di-GMP greatly augmented immune responses of glycopeptide cancer vaccines. Chem. Commun. 2018, 54, 9655–9658. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Celis, E. STING activator c-di-GMP enhances the anti-tumor effects of peptide vaccines in melanoma-bearing mice. Cancer Immunol. Immunother. 2015, 64, 1057–1066. [Google Scholar] [CrossRef]
- Shae, D.; Baljon, J.J.; Wehbe, M.; Christov, P.P.; Becker, K.W.; Kumar, A.; Suryadevara, N.; Carson, C.S.; Palmer, C.R.; Knight, F.C.; et al. Co-delivery of Peptide Neoantigens and Stimulator of Interferon Genes Agonists Enhances Response to Cancer Vaccines. ACS Nano 2020, 14, 9904–9916. [Google Scholar] [CrossRef]
- Ebensen, T.; Delandre, S.; Prochnow, B.; Guzmán, C.A.; Schulze, K. The Combination Vaccine Adjuvant System Alum/c-di-AMP Results in Quantitative and Qualitative Enhanced Immune Responses Post Immunization. Front. Cell. Infect. Microbiol. 2019, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.G.; Wu, J.J.; Zhang, B.D.; Li, W.H.; Li, Y.M. Pam3CSK4-CDG(SF) Augments Antitumor Immunotherapy by Synergistically Activating TLR1/2 and STING. Bioconjug. Chem. 2020, 31, 2499–2503. [Google Scholar] [CrossRef]
- Alyaqoub, F.S.; Aldhamen, Y.A.; Koestler, B.J.; Bruger, E.L.; Seregin, S.S.; Pereira-Hicks, C.; Godbehere, S.; Waters, C.M.; Amalfitano, A. In Vivo Synthesis of Cyclic-di-GMP Using a Recombinant Adenovirus Preferentially Improves Adaptive Immune Responses against Extracellular Antigens. J. Immunol. 2016, 196, 1741–1752. [Google Scholar] [CrossRef]
- Koestler, B.; Seregin, S.S.; Rastall, D.P.W.; Aldhamen, Y.A.; Godbehere, S.; Amalfitano, A.; Waters, C.M. Stimulation of Innate Immunity byIn VivoCyclic di-GMP Synthesis Using Adenovirus. Clin. Vaccine Immunol. 2014, 21, 1550–1559. [Google Scholar] [CrossRef] [Green Version]
- Bridgeman, A.; Maelfait, J.; Davenne, T.; Partridge, T.; Peng, Y.; Mayer, A.; Dong, T.; Kaever, V.; Borrow, P.; Rehwinkel, J. Viruses transfer the antiviral second messenger cGAMP between cells. Science 2015, 349, 1228–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana, I.; Espariz, M.; Villar, S.R.; Gonzalez, F.B.; Pacini, M.F.; Cabrera, G.; Bontempi, I.; Prochetto, E.; Stulke, J.; Perez, A.R.; et al. Genetic Engineering of Lactococcus lactis Co-producing Antigen and the Mucosal Adjuvant 3′ 5′- cyclic di Adenosine Monophosphate (c-di-AMP) as a Design Strategy to Develop a Mucosal Vaccine Prototype. Front. Microbiol. 2018, 9, 2100. [Google Scholar] [CrossRef]
- Konno, H.; Chinn, I.K.; Hong, D.; Orange, J.S.; Lupski, J.R.; Mendoza, A.; Pedroza, L.A.; Barber, G.N. Pro-inflammation Associated with a Gain-of-Function Mutation (R284S) in the Innate Immune Sensor STING. Cell Rep. 2018, 23, 1112–1123. [Google Scholar] [CrossRef] [Green Version]
- Ozasa, K.; Temizoz, B.; Kusakabe, T.; Kobari, S.; Momota, M.; Coban, C.; Ito, S.; Kobiyama, K.; Kuroda, E.; Ishii, K.J. Cyclic GMP-AMP Triggers Asthma in an IL-33-Dependent Manner That Is Blocked by Amlexanox, a TBK1 Inhibitor. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Konno, H.; Konno, K.; Barber, G.N. Cyclic Dinucleotides Trigger ULK1 (ATG1) Phosphorylation of STING to Prevent Sustained Innate Immune Signaling. Cell 2013, 155, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Rueckert, C.; Rand, U.; Roy, U.; Kasmapour, B.; Strowig, T.; Guzmán, C.A. Cyclic dinucleotides modulate induced type I IFN responses in innate immune cells by degradation of STING. FASEB J. 2017, 31, 3107–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyabe, H.; Hyodo, M.; Nakamura, T.; Sato, Y.; Hayakawa, Y.; Harashima, H. A new adjuvant delivery system ‘cyclic di-GMP/YSK05 liposome’ for cancer immunotherapy. J. Control. Release 2014, 184, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Kocabas, B.B.; Almacioglu, K.; Bulut, E.A.; Gucluler, G.; Tincer, G.; Bayik, D.; Gursel, M.; Gursel, I. Dual-adjuvant effect of pH-sensitive liposomes loaded with STING and TLR9 agonists regress tumor development by enhancing Th1 immune response. J. Control. Release 2020, 328, 587–595. [Google Scholar] [CrossRef]
- Hanson, M.C.; Crespo, M.P.; Abraham, W.; Moynihan, K.D.; Szeto, G.L.; Chen, S.H.; Melo, M.B.; Mueller, S.; Irvine, D.J. Nanoparticulate STING agonists are potent lymph node–targeted vaccine adjuvants. J. Clin. Investig. 2015, 125, 2532–2546. [Google Scholar] [CrossRef] [Green Version]
- Shakya, A.K.; Lee, C.H.; Uddin, J.; Gill, H.S. Assessment of Th1/Th2 Bias of STING Agonists Coated on Microneedles for Possible Use in Skin Allergen Immunotherapy. Mol. Pharm. 2018, 15, 5437–5443. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Schulze, K.; Ebensen, T.; Weissmann, S.; Hansen, S.; Guzmán, C.A.; Lehr, C.-M. Inverse micellar sugar glass (IMSG) nanoparticles for transfollicular vaccination. J. Control. Release 2015, 206, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Jang, H.-E.; Kang, Y.Y.; Kim, J.; Ahn, J.-H.; Mok, H. Submicron-sized hydrogels incorporating cyclic dinucleotides for selective delivery and elevated cytokine release in macrophages. Acta Biomater. 2016, 29, 271–281. [Google Scholar] [CrossRef]
- Koshy, S.T.; Cheung, A.; Gu, L.; Graveline, A.; Mooney, D.J. Liposomal Delivery Enhances Immune Activation by STING Agonists for Cancer Immunotherapy. Adv. Biosyst. 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qiu, M.; Ye, Z.; Nyalile, T.; Li, Y.; Glass, Z.; Zhao, X.; Yang, L.; Chen, J.; Xu, Q. In situ cancer vaccination using lipidoid nanoparticles. Sci. Adv. 2021, 7, eabf1244. [Google Scholar] [CrossRef]
- Collier, M.A.; Junkins, R.D.; Gallovic, M.D.; Johnson, B.M.; Johnson, M.M.; Macintyre, A.N.; Sempowski, G.D.; Bachelder, E.M.; Ting, J.P.Y.; Ainslie, K.M. Acetalated Dextran Microparticles for Codelivery of STING and TLR7/8 Agonists. Mol. Pharm. 2018, 15, 4933–4946. [Google Scholar] [CrossRef]
- Junkins, R.D.; Gallovic, M.D.; Johnson, B.M.; Collier, M.A.; Watkins-Schulz, R.; Cheng, N.; David, C.N.; McGee, C.E.; Sempowski, G.D.; Shterev, I.; et al. A robust microparticle platform for a STING-targeted adjuvant that enhances both humoral and cellular immunity during vaccination. J. Control. Release 2018, 270, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.H.; Cossette, B.J.; Varadhan, A.K.; Wu, Y.; Collier, J.H. Titrating Polyarginine into Nanofibers Enhances Cyclic-Dinucleotide Adjuvanticity in Vitro and after Sublingual Immunization. ACS Biomater. Sci. Eng. 2021, 7, 1876–1888. [Google Scholar] [CrossRef]
- Shae, D.; Becker, K.W.; Christov, P.; Yun, D.S.; Lytton-Jean, A.K.R.; Sevimli, S.; Ascano, M.; Kelley, M.; Johnson, D.B.; Balko, J.M.; et al. Endosomolytic polymersomes increase the activity of cyclic dinucleotide STING agonists to enhance cancer immunotherapy. Nat. Nanotechnol. 2019, 14, 269–278. [Google Scholar] [CrossRef]
- Schulze, K.; Ebensen, T.; Babiuk, L.A.; Gerdts, V.; Guzman, C.A. Intranasal vaccination with an adjuvanted polyphosphazenes na-noparticle-based vaccine formulation stimulates protective immune responses in mice. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2169–2178. [Google Scholar] [CrossRef]
- Leach, D.G.; Dharmaraj, N.; Piotrowski, S.L.; Lopez-Silva, T.L.; Lei, Y.; Sikora, A.G.; Young, S.; Hartgerink, J.D. STINGel: Controlled release of a cyclic dinucleotide for enhanced cancer immunotherapy. Biomaterials 2018, 163, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Miyabe, H.; Hyodo, M.; Sato, Y.; Hayakawa, Y.; Harashima, H. Liposomes loaded with a STING pathway ligand, cyclic di-GMP, enhance cancer immunotherapy against metastatic melanoma. J. Control. Release 2015, 216, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demoulins, T.; Ruggli, N.; Gerber, M.; Thomann-Harwood, L.J.; Ebensen, T.; Schulze, K.; Guzman, C.A.; McCullough, K.C. Self-Amplifying Pestivirus Replicon RNA Encoding Influenza Virus Nucleoprotein and Hemagglutinin Promote Humoral and Cellular Immune Responses in Pigs. Front. Immunol. 2020, 11, 622385. [Google Scholar] [CrossRef]
- Gogoi, H.; Mansouri, S.; Jin, L. The Age of Cyclic Dinucleotide Vaccine Adjuvants. Vaccines 2020, 8, 453. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Brendel, V.P.; Shu, C.; Li, P.; Palanathan, S.; Kao, C.C. Single Nucleotide Polymorphisms of Human STING Can Affect Innate Immune Response to Cyclic Dinucleotides. PLoS ONE 2013, 8, e77846. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CDNs | Others | Species | Immunization Route | Antigens | Immunogenicity | Efficacy | References |
|---|---|---|---|---|---|---|---|
| c-di-GMP | Mice | SC | Beta-gal | Serum IgG, Th1/Th2 | [91] | ||
| c-di-GMP | Mice | IN | Beta-gal, Ova | Serum IgG, mucosal IgA, Th1, CTL | [92] | ||
| c-di-GMP | Mice | IN, IM | Influenza H5N1 (A/Anhui/1/05) | Mucosal IgA, serum IgG, balanced Th1/Th2, high frequencies of multifunctional Th1 CD4+ cells. IN > IM | [105] | ||
| c-di-GMP | Mice | IN, sublingual, IM | Influenza H5N1 (A/Anhui/1/05) | Mucosal IgA, serum IgG, balanced Th1/Th2, high frequencies of splenic H5N1-specific multifunctional (IL-2+TNF-α+) CD4+ T cells. IN or SL > IM | [106] | ||
| c-di-GMP | Chitosan | Mice | IN | NIBRG-14 (H5N1) HA | Serum and local antibody responses, HI antibody, higher frequencies of virus-specific polyfunctional CD4+ T cells, more Th1 cytokines (IFN-γ, IL-2, TNF-α). 7.5 μg > 1.5 or 0.3 | [107] | |
| c-di-GMP | Silica nanoparticles (SiO2) | Mice | Intratracheal | Plant-produced H1N1 influenza HA | High systemic antibody responses, local IgG and IgA responses, local T-cell response (IL-2 and IFN-γ) | [108] | |
| c-di-GMP | Mice, ferrets | IN | HA A/Indonesia/05/05 (H5N1) | IgG and IgA antibodies | Survived the viral challenge | [109] | |
| c-di-GMP | microneedle | Mice | Microneedle skin patches | Influenza microneedle vaccine | Enhanced IgG, IgG subtypes, and cellular immune responses | 100% survival rate and rapid weight recovery | [110] |
| c-di-GMP | B. pertussis LP1569 (a TLR2 agonist) | Mice | SC, IP, IN | Acellular pertussis vaccine | IFN-β, IL-12 and IL-23 responses, maturation of dendritic cells; IN: Th1 and Th17 responses; Th17 response and IL-17-secreting TRM cells | IN: protection against nasal colonization and lung infection (sustained for at least 10 months) | [111] |
| c-di-GMP | Mice | SC | ClfA or mSEC (S. aureus) | High titers of IgG1, IgG2a, IgG2b and IgG3 compared to alum adjuvant mSEC > ClfA | Higher survival rates at day 7 with S. aureus challenge | [112] | |
| c-di-GMP | Mice | IN, IP | S. pneumoniae PdB or PspA | IP induced higher antigen-specific antibodies | Significant decrease in bacterial load in lungs and blood; IP: increased survival compared to that with alum adjuvant | [62] | |
| c-di-GMP | Mice | IN | S. pneumonia PsaA | Strong IgG and IgA | Significantly reduced nasopharyngeal colonization | [61] | |
| c-di-GMP | DNA vaccine | Mice | ID | HIV-1 reverse transcriptase | Transient increase in IFN-γ production | [113] | |
| c-di-GMP | Mice, cattle, pigs | IM | Foot and mouth disease vaccine | Early onset of high neutralizing antibody titers; long-lasting immune memory response | [114] | ||
| c-di-GMP | Mice | IN | Ghrelin-PspA | Specific antibody responses | Reduced body weight gain in diet-induced obesity mice | [115] | |
| c-di-GMP | Rats, mice | IN | Angiotensin II type 1 receptor (AT1R) peptide conjugated with PspA | Specific antibody responses | Prevented the development of hypertension in spontaneously hypertensive rats; Sera protected mice against lethal pneumococcal infection | [116] | |
| c-di-GMP and c-di-AMP | Mice | IN, sublingual | H5N1 virosomes | Local and systemic humoral and cellular immune responses; long-lasting immunity; dose-sparing | Effective protection against influenza H5N1 | [117] | |
| c-di-AMP | Mice | SC | Ova (soluble or DC targeted) | Serum IgG, Th1, CTL, and IFNγ-producing CD8 memory T-cell response, better than PolyI:C/CpG | [96] | ||
| c-di-AMP | Mice | IN | Influenza nucleoprotein | IgG and IgA responses, strong Th1 response (IFN-γ and IL-2) | Protection against A/Puerto Rico/8/34 (H1N1) virus | [118] | |
| c-di-AMP | Mice | IN | ESAT-6 | Innate and adaptive immune responses and regulated autophagy of macrophages | Protection against i.v. challenge similar to antigen alone group | [119] | |
| c-di-AMP | Lipopeptide-based nano carrier systems | Mice | IN | T and B cell epitopes of S. pyogenes M protein | Antigen dose-sparing | [120] | |
| c-di-AMP | Archaeosomes | Mice | IN-IN-IN, IM-IN-IN, or IM-IM-IM | HCV rE1E2 | Induced more robust polyfunctional CD4+ T-cell responses | [121] | |
| c-di-AMP | Mice | IN | BtaF trimeric autotransporter | Local and systemic antibody responses, central memory CD4+ T cells, and strong Th1 responses | Protection against intragastric, but not respiratory, challenge with B. suis | [122] | |
| c-di-AMP | Conjugated at N-terminal of target antigen | Mice | IN | T. cruzi recombinant Tc52 | Predominantly Th17 and Th1 immune responses | Better protection against infection with lower parasitemia and weight loss, and higher survival rates | [123] |
| c-di-AMP | Mice | IN | Traspain (trivalent T. cruzi antigen) | Primed Th1/Th17 immune response | Reduced parasite load and chronic inflammation | [124,125] | |
| c-di-AMP | Poly(lactic-co-glycolic acid (PLGA) microparticles | Piglets | IM | Inactivated M. hyopneumoniae field isolate BA 2940-99 | Strong innate immune responses and robust Th1 or Th17 responses | [126] | |
| c-di-AMP or c-di-IMP | Mice | IN | Beta-gal, Ova | Serum IgG, mucosal IgA, Th1/Th2/Th17, CTL | [49,93] | ||
| cGAMP | Mice | IN, single dose | Inactivated whole virus H7N9 vaccine | Enhanced humoral, cellular, and mucosal immune responses. significantly higher nucleoprotein-specific CD4+ and CD8+ T-cell responses | Protection against a high lethal dose, effective cross-protection against H1N1, H3N2, and H9N2 influenza virus | [127] | |
| cGAMP | Mice | IN | HA vaccine | Enhanced IgA, IgG, and T cell responses, including in nasal-associated lymphoid tissue | [128] | ||
| cGAMP | Pigs | ID | H5N1 and 2009 H1N1 pandemic influenza vaccines | Vigorous immune responses elicited with no overt skin irritation | [129] | ||
| cGAMP | Biomimetic liposomes | Mice, ferrets | IN | H1N1 vaccine | Vigorously augmented humoral and CD8+ T-cell responses, importance of alveolar epithelial cells in heterosubtypic immunity | Strong cross-protection against distant H1N1 and heterosubtypic H3N2, H5N1, and H7N9 viruses | [130] |
| cGAMP | Microneedle patches and saponin (Quil-A) | Aged mice | Microneedle patches | H1N1 vaccine | Increased IgG and IgG2a | Complete protection | [131] |
| cGAMP | Mice | IN, SC, IM | H. pylori urease A, urease B, and neutrophil-activating protein | Serum IgG and mucosal IgA, Th1, and particularly Th17 responses (IN, SC), IL-17 responses (IN) | Significantly reduced gastric mucosal H. pylori colonization (IN, SC) | [132] | |
| cGAMP | Mice | IM | M. tuberculosis Rv3852 (H-NS) | High serum levels of IL-2, IL-12p40, and TNF-α, specific CD4 and CD8 T-cell responses | Reduced bacterial counts in the spleen but not in the lung after H37Rv challenge | [133] | |
| cGAMP | CpG-C ODN | Mice | SC | HPV 16 E7 (E7GRG) | High IgG level and cell proliferation; IFN-γ and granzyme B levels | Suppressed TC-1 tumour growth | [134] |
| cGAMP | Mice | Not specified | HBsAg | Significantly enhanced humoral and cellular immune responses | [135] | ||
| cGAMP | Alum (Alhydrogel) | Mice (newborns) | IM | rHA influenza vaccine (Flublok) | Enhanced HA-specific IgG2a/c titers, IFNγ production by CD4+ T cells, increased T follicular helper cell and GL-7+CD138+ germinal centre B cell responses in newborns | [136] | |
| 3′3′ cGAMP | Acetalated dextran (Ace-DEX) microparticules | Mice | IM | Influenza matrix protein 2 (M2e) | Humoral and cellular responses, broadly protective immunity | Substantial cross-protection against distant H1N1 and heterosubtypic H3N2, H5N1, and H7N9 viruses | [137] |
| 3′3′ cGAMP | Mice | SL | Bacillus anthracis protective antigen (PA) | Higher serum IgG and saliva IgA than PA + cholera toxin; Th1, Th2, and Th17 responses; rapid IFN-β and IL-10 responses in sublingual tissues and cervical lymph | [138] | ||
| RR-CDG | Mice | IN, SC | Antigen-85B, ESAT-6, Rv1733c, Rv2626c, and RpfD fusion protein | IN: significantly boosted BCG-induced immunity; elicited both Th1 and Th17 immune responses | IN: superior protection than BCG | [139] | |
| RR-CDG | Nanoparticles | Mice | SC | Y. pestis F1-V | Induced rapid and long-lived protective immunity | 100% protected from Y. pestis lethal challenge within 14 days post-immunization, and 75% protected at 182 days post-immunization | [140] |
| rBCG-disA-OE | Endogenous c-di-AMP | Guinea pigs | ID | BCG | Significantly stronger TNF-α, IL-6, IL-1β, IRF3, and IFN-β levels than BCG in murine macrophages culture | Significantly reduced lung weights, pathology scores, and bacterial counts in lungs vs. BCG | [104] |
| rBCG-DisA | Endogenous c-di-AMP | Mice | SC | BCG | Produced more IFN-γ, IL-2, and IL-10, stronger expression of H3K4me3 vs. BCG | No additional protection against M. tuberculosis infection | [103] |
| rBCG (BCGΔBCG1419c) | Mice | SC | BCG | Better activation of specific T-cell population vs. BCG | No difference in protection vs. BCG | [102] | |
| rBCG-Rv1357c | c-di-GMP phosphodiesterase gene Rv1357c | Mice | ID | BCG | More phagocytosed vs. BCG | Similar protection against M. tuberculosis challenge vs. BCG | [101] |
| CDNs | Deliver Systems | Function | Reference |
|---|---|---|---|
| CDN | Biodegradable, poly(beta-amino ester) nanoparticles | Cytosol delivery and 10-fold improved treatment potency in established B16 melanoma tumours with anti-PD-1 in mice compared to free CDN | [80] |
| c-di-GMP | Liposomes made of a synthetic, pH-sensitive lipid of high fusogenicity (YSK05) | Deliver into the cytosol and induced melanoma regression | [160,161] |
| c-di-GMP | Liposome nanoparticles | Targeting lymphatics and draining lymph nodes after parental injection | [162] |
| c-di-GMP and 3′3′-cGAMP | Cell-penetrating peptide | Enhanced both cellular delivery and biological activity of the c-di-GMP in murine splenocytes | [97] |
| c-di-GMP | Cytotoxic cationic silica nanoparticles | Local retention and potent melanoma growth inhibition | [79] |
| c-di-GMP | Mesoporous silica nanoparticle (immuno-MSN) | Brain delivery for treatment of glioblastoma multiforme | [77] |
| c-di-GMP | Rhodamine B isothiocyanate fluorescent mesoporous silica nanoparticles, synthesized and modified with poly(ethylene glycol) and an ammonium-based cationic molecule | “In situ vaccination” strategy; dramatic inhibition of 4T1 breast tumour growth in mice | [78] |
| c-di-GMP and c-di-AMP | Microneedles (MN) | Skin delivery better than SC and MN-based alum | [163] |
| c-di-AMP | Surfactant-based inverse micellar sugar glass nanoparticles (NPs) | Needle-free transfollicular antigen delivery from intact skin; better than polylactic-co-glycolic acid (PLGA) and chitosan-PLGA NPs | [164] |
| c-di-AMP | Cationic liposomes | Enhanced M. hyopneumoniae-specific antibody and T-cell responses | [126] |
| cGAMP | A polymersome delivery platform | Improved half-life of cGAMP, enhanced immune activation, and tumour growth suppression | [74] |
| cGAMP | Linear polyethyleneimine/hyaluronic acid hydrogels | Delivered into phagocytic macrophage cells | [165] |
| cGAMP | Cationic liposomes with varying surface polyethylene glycol levels | Cytosolic delivery and delivery to metastatic melanoma tumour sites and APCs | [166] |
| cGAMP | Microneedle skin patches (MNPs) | Increased IgG and IgG2a responses to influenza vaccines in aged mice (21-month-old), and full protection against challenge | [131] |
| cGAMP | Lipidoid nanoparticle (93-O17S-F) | Cytosolic delivery | [167] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Chen, W. The Promise and Challenges of Cyclic Dinucleotides as Molecular Adjuvants for Vaccine Development. Vaccines 2021, 9, 917. https://doi.org/10.3390/vaccines9080917
Yan H, Chen W. The Promise and Challenges of Cyclic Dinucleotides as Molecular Adjuvants for Vaccine Development. Vaccines. 2021; 9(8):917. https://doi.org/10.3390/vaccines9080917
Chicago/Turabian StyleYan, Hongbin, and Wangxue Chen. 2021. "The Promise and Challenges of Cyclic Dinucleotides as Molecular Adjuvants for Vaccine Development" Vaccines 9, no. 8: 917. https://doi.org/10.3390/vaccines9080917