
Effects of a Recombinant Gonadotropin-Releasing Hormone Vaccine on Reproductive Function in Adult Male ICR Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine Preparation
2.3. Immunization
2.4. Sample Collection
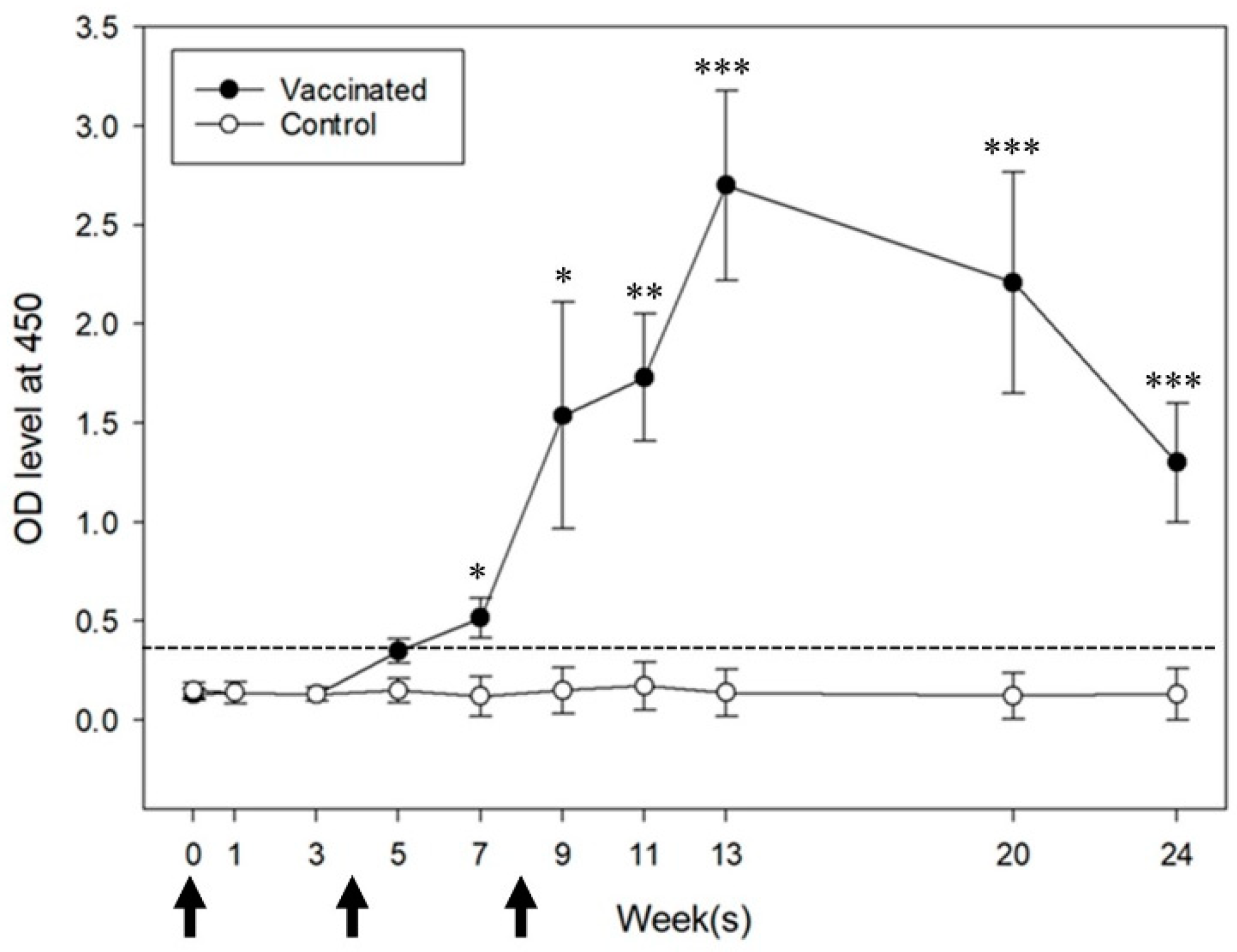
2.5. ELISA Measurement of Anti-GnRH IgG Antibody
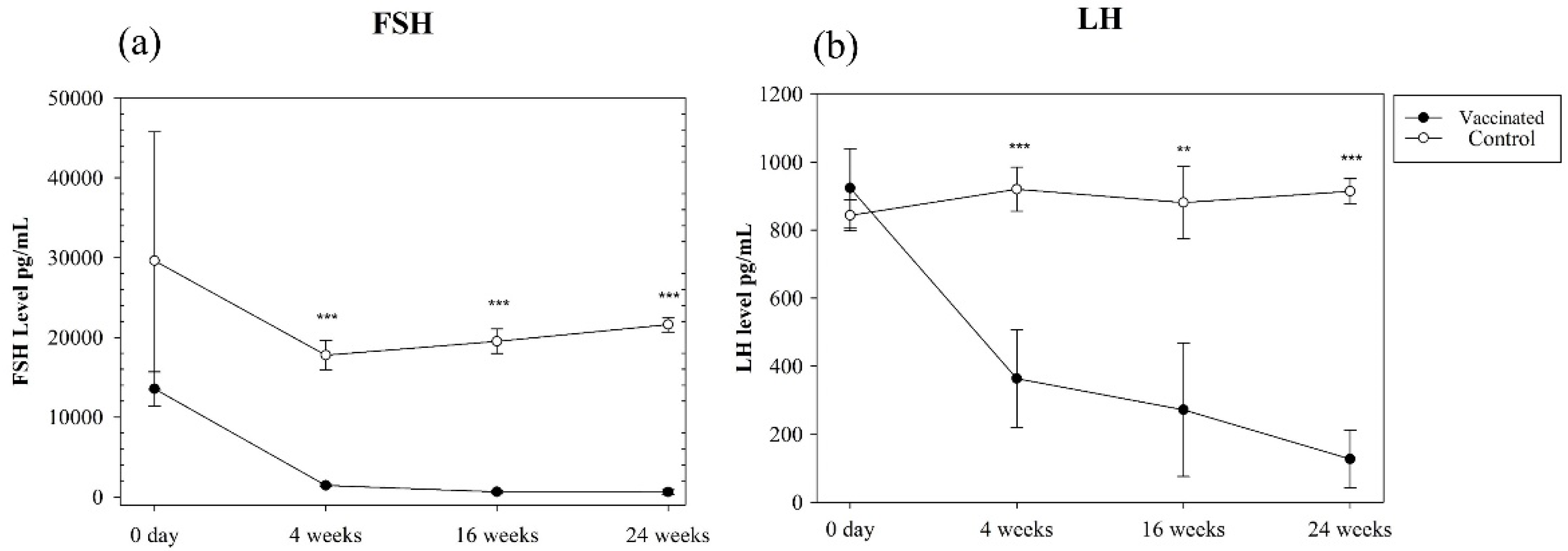
2.6. Measurement of Serum LH/FSH Concentrations
2.7. Measurement of Serum Testosterone Concentrations
2.8. Intra-Assay and Inter-Assay Variation
2.9. Analysis of Testis Tissue and Sperm Quality
2.9.1. Testis Morphology Measurement
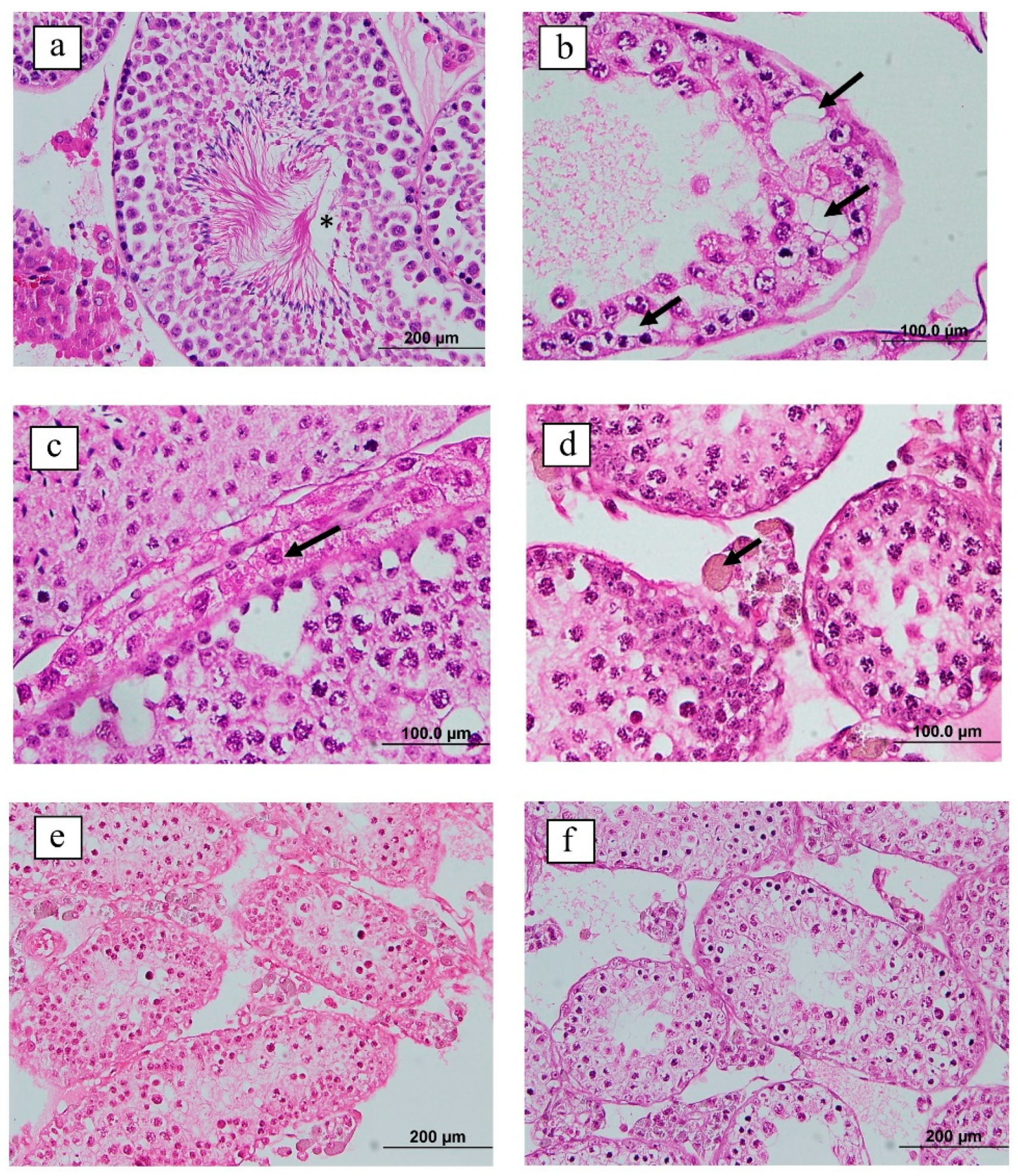
2.9.2. Histological Examination of the Testis
2.9.3. Sperm Collection
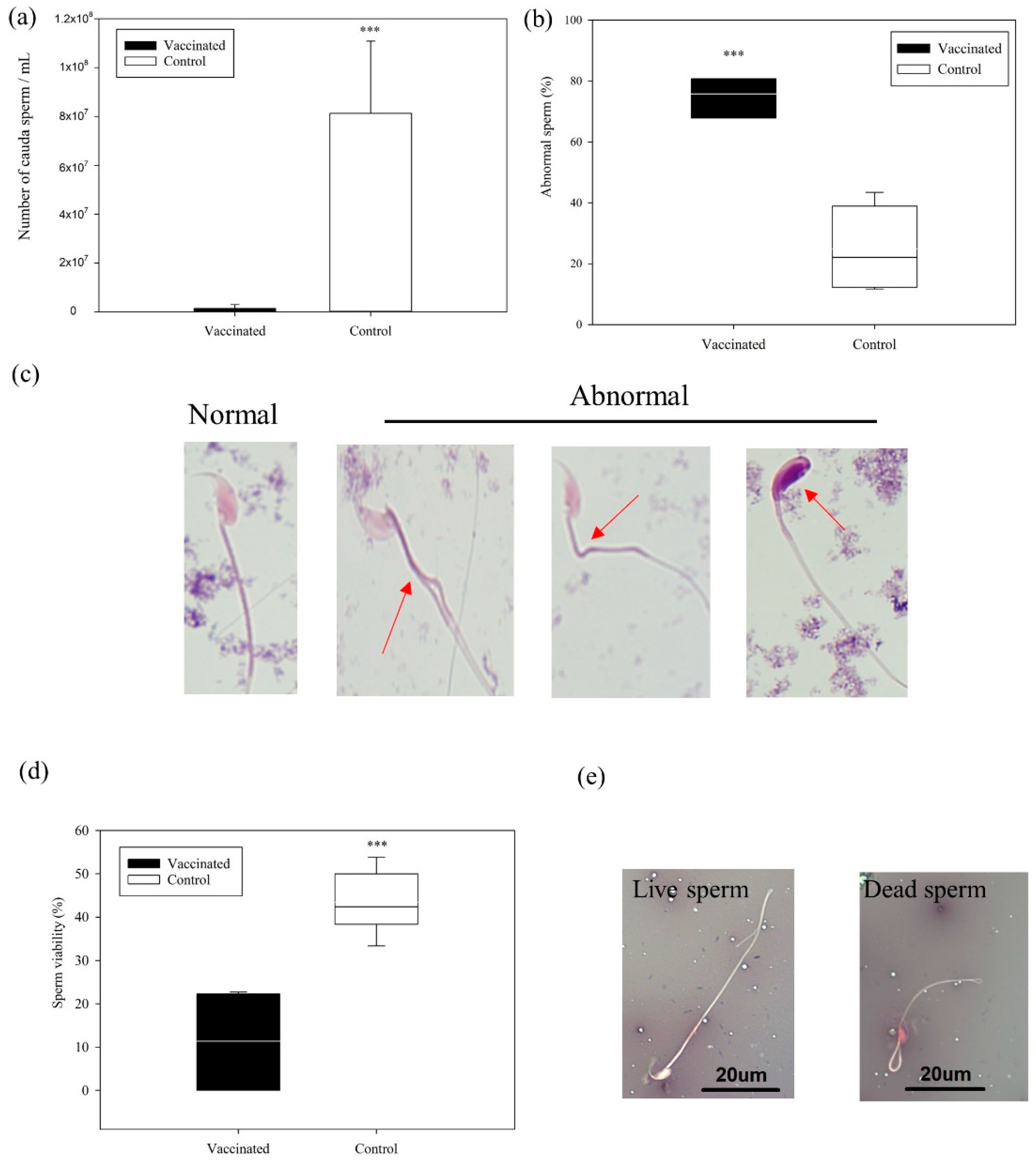
2.9.4. Analysis of Sperm Quality
2.10. Fertility Test
2.11. Statistical Analysis
3. Results
3.1. Specific Anti-GnRH Immune Response and Serum FSH, LH, and Testosterone Concentrations
3.2. Effects of GnRH Vaccine on Testis Weight and Histomorphology
3.3. Cauda Epididymal Sperm Analysis
3.4. Mating Behavior and Fertility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, D.R. The Problems of Overpopulation of Deer in North America; Academic Press: New York, NY, USA, 1981; Volume 119. [Google Scholar]
- Rowan, A.N. Shelters and Pet Overpopulation: A Statistical Black Hole; Taylor & Francis: Abingdon, UK, 1992. [Google Scholar]
- Bertschinger, H.J.; Caldwell, P. Fertility suppression of some wildlife species in southern Africa—A review. Reprod. Domest. Anim. 2016, 51, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Bomford, M.; O’Brien, P. A role for fertility control wildlife management in Australia? In Proceedings of the Vertebrate Pest Conference, Newport Beach, CA, USA, 3–5 March 1992. [Google Scholar]
- Driscoll, D.A.; Worboys, G.L.; Allan, H.; Banks, S.C.; Beeton, N.J.; Cherubin, R.C.; Doherty, T.S.; Finlayson, C.M.; Green, K.; Hartley, R. Impacts of feral horses in the Australian Alps and evidence-based solutions. Ecol. Manag. Restor. 2019, 20, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Warren, R. Deer overabundance in the USA: Recent advances in population control. Anim. Prod. Sci. 2011, 51, 259–266. [Google Scholar] [CrossRef]
- Hsu, M.J.; Kao, C.C.; Agoramoorthy, G. Interactions between visitors and Formosan macaques (Macaca cyclopis) at Shou-Shan Nature Park, Taiwan. Am. J. Primatol. Off. J. Am. Soc. Primatol. 2009, 71, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Loydi, A.; Zalba, S.M. Feral horses dung piles as potential invasion windows for alien plant species in natural grasslands. In Herbaceous Plant Ecology; Springer: Berlin, Germany, 2008; pp. 107–116. [Google Scholar]
- Stewart, C.M.; Mcshea, W.J.; Piccolo, B.P. The impact of white-tailed deer on agricultural landscapes in 3 National Historical Parks in Maryland. J. Wildl. Manag. 2007, 71, 1525–1530. [Google Scholar] [CrossRef]
- Massei, G.; Cowan, D. Fertility control to mitigate human–wildlife conflicts: A review. Wildl. Res. 2014, 41, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Duka, T.; Masters, P. Confronting a tough issue: Fertility control and translocation for over-abundant Koalas on Kangaroo Island, South Australia. Ecol. Manag. Restor. 2005, 6, 172–181. [Google Scholar] [CrossRef]
- Fagerstone, K.A.; Miller, L.A.; Killian, G.; Yoder, C.A. Review of issues concerning the use of reproductive inhibitors, with particular emphasis on resolving human-wildlife conflicts in North America. Integr. Zool. 2010, 5, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Carter, S.P.; Delahay, R.J.; Smith, G.C.; Macdonald, D.W.; Riordan, P.; Etherington, T.R.; Pimley, E.R.; Walker, N.J.; Cheeseman, C.L. Culling-induced social perturbation in Eurasian badgers Meles meles and the management of TB in cattle: An analysis of a critical problem in applied ecology. Proc. R. Soc. B Biol. Sci. 2007, 274, 2769–2777. [Google Scholar] [CrossRef] [Green Version]
- Massei, G. Catch, inject and release: Immunocontraception as alternative to culling and surgical sterilisation to control rabies in freeroaming dogs. In Proceedings of the Compendium of the Office International Epizooties Global Conference on Rabies Control, Seoul, Korea, 7–9 September 2011; pp. 181–187. [Google Scholar]
- Pepin, K.M.; Davis, A.J.; Cunningham, F.L.; Ver Cauteren, K.C.; Eckery, D.C. Potential effects of incorporating fertility control into typical culling regimes in wild pig populations. PLoS ONE 2017, 12, e0183441. [Google Scholar] [CrossRef]
- Lieury, N.; Ruette, S.; Devillard, S.; Albaret, M.; Drouyer, F.; Baudoux, B.; Millon, A. Compensatory immigration challenges predator control: An experimental evidence-based approach improves management. J. Wildl. Manag. 2015, 79, 425–434. [Google Scholar] [CrossRef]
- Lawton, C.; Rochford, J. The recovery of grey squirrel (Sciurus carolinensis) populations after intensive control programmes. R. Ir. Acad. 2007, 107, 19–29. [Google Scholar] [CrossRef]
- Massei, G.; Quy, R.J.; Gurney, J.; Cowan, D.P. Can translocations be used to mitigate human–wildlife conflicts? Wildl. Res. 2010, 37, 428–439. [Google Scholar] [CrossRef]
- McLeod, S.R.; Saunders, G. Fertility control is much less effective than lethal baiting for controlling foxes. Ecol. Model. 2014, 273, 1–10. [Google Scholar] [CrossRef]
- Croft, S.; Franzetti, B.; Gill, R.; Massei, G. Too many wild boar? Modelling fertility control and culling to reduce wild boar numbers in isolated populations. PLoS ONE 2020, 15, e0238429. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F.; Lyda, R.O.; Frank, K.M. Contraceptive vaccines for wildlife: A review. Am. J. Reprod. Immunol. 2011, 66, 40–50. [Google Scholar] [CrossRef]
- Lescheid, D.; Powell, J.; Fischer, W.; Park, M.; Craig, A.; Bukovskaya, O.; Barannikova, I.; Sherwood, N. Mammalian gonadotropin-releasing hormone (GnRH) identified by primary structure in Russian sturgeon, Acipenser gueldenstaedti. Regul. Pept. 1995, 55, 299–309. [Google Scholar] [CrossRef]
- Miller, L.A.; Johns, B.E.; Killian, G.J. Immunocontraception of White-Tailed Deer with GnRH Vaccine. Am. J. Reprod. Immunol. 2000, 44, 256–273. [Google Scholar] [CrossRef] [Green Version]
- Sabeur, K.; Ball, B.; Nett, T.; Ball, H.; Liu, I. Effect of GnRH conjugated to pokeweed antiviral protein on reproductive function in adult male dogs. Reproduction 2003, 125, 801–806. [Google Scholar] [CrossRef]
- Millar, R.P. GnRHs and GnRH receptors. Anim. Reprod. Sci. 2005, 88, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.H.; Lee, L.T.; Chow, B.K. Gonadotropin-releasing hormone: Regulation of the GnRH gene. FEBS J. 2008, 275, 5458–5478. [Google Scholar] [CrossRef]
- Killian, G.; Wagner, D.; Miller, L. Observations on the use of the GnRH vaccine GonaCon™ in male white-tailed deer (Odocoileus virginianus). In Proceedings of the 11th Wildlife Damage Management Conference, Traverse City, MI, USA, 16–19 May 2005. [Google Scholar]
- Fagerstone, K.A.; Miller, L.A.; Eisemann, J.D.; O’Hare, J.R.; Gionfriddo, J.P. Registration of wildlife contraceptives in the United States of America, with OvoControl and GonaCon immunocontraceptive vaccines as examples. Wildl. Res. 2008, 35, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.A.; Fagerstone, K.A.; Eckery, D.C. Twenty years of immunocontraceptive research: Lessons learned. J. Zoo Wildl. Med. 2013, 44, S84–S96. [Google Scholar] [CrossRef] [Green Version]
- Food, U.; Administration, D. Freedom of Information Summary. Original New animal Drug Application; New Animal Drug Application (NADA): Washington, DC, USA, 2006; pp. 141–258.
- Vargas-Pino, F.; Gutiérrez-Cedillo, V.; Canales-Vargas, E.J.; Gress-Ortega, L.R.; Miller, L.A.; Rupprecht, C.E.; Bender, S.C.; García-Reyna, P.; Ocampo-López, J.; Slate, D. Concomitant administration of GonaCon™ and rabies vaccine in female dogs (Canis familiaris) in Mexico. Vaccine 2013, 31, 4442–4447. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ferro, V.A.; Koyama, S.; Kinugasa, Y.; Song, M.; Ogita, K.; Tsutsui, T.; Murata, Y.; Kimura, T. Immunisation of male mice with a plasmid DNA vaccine encoding gonadotrophin releasing hormone (GnRH-I) and T-helper epitopes suppresses fertility in vivo. Vaccine 2007, 25, 3544–3553. [Google Scholar] [CrossRef]
- Siel, D.; Vidal, S.; Sevilla, R.; Paredes, R.; Carvallo, F.; Lapierre, L.; Maino, M.; Pérez, O.; Sáenz, L. Effectiveness of an immunocastration vaccine formulation to reduce the gonadal function in female and male mice by Th1/Th2 immune response. Theriogenology 2016, 86, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.G.; Barthold, S.; Davisson, M.; Newcomer, C.E.; Quimby, F.W.; Smith, A. The Mouse in Biomedical Research: Normative Biology, Husbandry, and Models; Elsevier: Amsterdam, The Netherlands, 2006; Volume 3. [Google Scholar]
- Kimura, K.; Takeuchi, K. Aging and longevity of the Jel: ICR mouse. Okajimas Folia Anat. Jpn. 1988, 65, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Talwar, G.P.; Raina, K.; Gupta, J.C.; Ray, R.; Wadhwa, S.; Ali, M.M. A recombinant luteinising-hormone-releasing-hormone immunogen bioeffective in causing prostatic atrophy. Vaccine 2004, 22, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Epididymal sperm count. Curr. Protoc. Toxicol. 2002, 14, 16.16.11–16.16.15. [Google Scholar] [CrossRef]
- Classen, D.C.; Morningstar, J.M.; Shanley, J.D. Detection of antibody to murine cytomegalovirus by enzyme-linked immunosorbent and indirect immunofluorescence assays. J. Clin. Microbiol. 1987, 25, 600–604. [Google Scholar] [CrossRef] [Green Version]
- Pitetti, J.-L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M. An essential role for insulin and IGF1 receptors in regulating sertoli cell proliferation, testis size, and FSH action in mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef]
- Reed, G.F.; Lynn, F.; Meade, B.D. Use of coefficient of variation in assessing variability of quantitative assays. Clin. Diagn. Lab. Immunol. 2002, 9, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- Spears, J.; Matthews, A.; Hartwig, S. Evaluation of noninvasive methods of predicting testicular weight and development in an inbred and outbred strain of laboratory mouse. Theriogenology 2013, 79, 566–573. [Google Scholar] [CrossRef]
- Luthfi, M.J.F. A simple and practical method for rat epididymal sperm count (Rattus norvegicus). Biol. Med. Nat. Prod. Chem. 2015, 4, 1–3. [Google Scholar] [CrossRef]
- Didion, B.; Dobrinsky, J.; Giles, J.; Graves, C. Staining procedure to detect viability and the true acrosome reaction in spermatozoa of various species. Gamete Res. 1989, 22, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Gupta, S.; Sharma, R. Eosin-Nigrosin staining procedure. In Andrological Evaluation of Male Infertility; Springer: Berlin/Heidelberg, Germany, 2016; pp. 73–77. [Google Scholar]
- Wyrobek, A.J.; Laurie, A.G.; James, G.B.; Mary, W.F.; Robert, W.K., Jr.; Gideon, L.; Heinrich, V.M.; John, C.T.; Whorton, M.D. An evaluation of the mouse sperm morphology test and other sperm tests in nonhuman mammals: A report of the US Environmental Protection Agency Gene-Tox Program. Mutat. Res. Rev. Genet. Toxicol. 1983, 115, 1–72. [Google Scholar] [CrossRef]
- Khan, M.A.; Ogita, K.; Ferro, V.A.; Kumasawa, K.; Tsutsui, T.; Kimura, T. Immunisation with a plasmid DNA vaccine encoding gonadotrophin releasing hormone (GnRH-I) and T-helper epitopes in saline suppresses rodent fertility. Vaccine 2008, 26, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, S. Progestins to control feline reproduction: Historical abuse of high doses and potentially safe use of low doses. J. Feline Med. Surg. 2015, 17, 743–752. [Google Scholar] [CrossRef]
- Massei, G.; Miller, L.A. Nonsurgical fertility control for managing free-roaming dog populations: A review of products and criteria for field applications. Theriogenology 2013, 80, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, R.K.; Gupta, S.K.; Gupta, J.C.; Vyas, H.K.; Talwar, G.P. Recent advances in contraceptive vaccine development: A mini-review. Hum. Reprod. 2005, 20, 3271–3283. [Google Scholar] [CrossRef] [Green Version]
- Naz, R.K. Recent progress toward development of vaccines against conception. Expert Rev. Vaccines 2014, 13, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K.; Saver, A.E. Immunocontraception for animals: Current status and future perspective. Am. J. Reprod. Immunol. 2016, 75, 426–439. [Google Scholar] [CrossRef]
- Junco, J.A.; Peschke, P.; Zuna, I.; Ehemann, V.; Fuentes, F.; Bover, E.; Pimentel, E.; Basulto, R.; Reyes, O.; Calzada, L. Immunotherapy of prostate cancer in a murine model using a novel GnRH based vaccine candidate. Vaccine 2007, 25, 8460–8468. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Hinds, L.A. Formulation and delivery of vaccines: Ongoing challenges for animal management. J. Pharm. Bioallied Sci. 2012, 4, 258. [Google Scholar]
- Ferro, V.A.; Khan, M.A.; Earl, E.R.; Harvey, M.J.; Colston, A.; Stimson, W.H. Influence of Carrier Protein Conjugation Site and Terminal Modification of a GnRH-I Peptide Sequence in the Development of a Highly Specific Anti-fertility Vaccine. Part I. Am. J. Reprod. Immunol. 2002, 48, 361–371. [Google Scholar] [CrossRef]
- Gupta, J.C.; Raina, K.; Talwar, G.; Verma, R.; Khanna, N. Engineering, cloning, and expression of genes encoding the multimeric luteinizing-hormone-releasing hormone linked to T cell determinants in Escherichia coli. Protein Expr. Purif. 2004, 37, 1–7. [Google Scholar] [CrossRef]
- Leenaars, P.P.; Koedam, M.A.; Wester, P.W.; Baumans, V.; Claassen, E.; Hendriksen, C. Assessment of side effects induced by injection of different adjuvant/antigen combinations in rabbits and mice. Lab. Anim. 1998, 32, 387–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, S.K.; Van Vuren, D.H.; Laursen, C.; Kelt, D.A. Behavioral effects of an immunocontraceptive vaccine on eastern fox squirrels. J. Wildl. Manag. 2015, 79, 1255–1263. [Google Scholar] [CrossRef]
- Baker, D.L.; Powers, J.G.; Ransom, J.I.; McCann, B.E.; Oehler, M.W.; Bruemmer, J.E.; Galloway, N.L.; Eckery, D.C.; Nett, T.M. Reimmunization increases contraceptive effectiveness of gonadotropin-releasing hormone vaccine (GonaCon-Equine) in free-ranging horses (Equus caballus): Limitations and side effects. PLoS ONE 2018, 13, e0201570. [Google Scholar] [CrossRef]
- Barnett, P.; Pullen, L.; Williams, L.; Doel, T. International bank for foot-and-mouth disease vaccine: Assessment of Montanide ISA 25 and ISA 206, two commercially available oil adjuvants. Vaccine 1996, 14, 1187–1198. [Google Scholar] [CrossRef]
- Aucouturier, J.; Ascarateil, S.; Dupuis, L. The use of oil adjuvants in therapeutic vaccines. Vaccine 2006, 24, S44–S45. [Google Scholar] [CrossRef]
- Esbenshade, K.; Britt, J. Active immunization of gilts against gonadotropin-releasing hormone: Effects on secretion of gonadotropins, reproductive function, and responses to agonists of gonadotropin-releasing hormone. Biol. Reprod. 1985, 33, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.; Wettemann, R.; Yelich, J.; Spicer, L. Ovarian response after gonadotropin treatment of heifers immunized against gonadotropin-releasing hormone. J. Anim. Sci. 1996, 74, 1092–1097. [Google Scholar] [CrossRef]
- Clarke, I.; Fraser, H.; McNeilly, A. Active immunization of ewes against luteinizing hormone releasing hormone, and its effects on ovulation and gonadotrophin, prolactin and ovarian steroid secretion. J. Endocrinol. 1978, 78, 39–47. [Google Scholar] [CrossRef]
- Fraser, H.; Baker, T. Changes in the ovaries of rats after immunization against luteinizing hormone releasing hormone. J. Endocrinol. 1978, 77, 85–93. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Weinbauer, G.F. Endocrine control of spermatogenesis: Role of FSH and LH/testosterone. Spermatogenesis 2014, 4, e996025. [Google Scholar] [CrossRef] [PubMed]
- Berndtson, W.E.; Thompson, T.L. Changing relationships between testis size, Sertoli cell number and spermatogenesis in Sprague-Dawley rats. J. Androl. 1990, 11, 429–435. [Google Scholar]
- Scudamore, C.L. A Practical Guide to the Histology of the Mouse; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Misro, M.; Ganguly, A.; Das, R. Adverse effects of chronic GnRH antagonist administration on seminiferous epithelium in adult rats. Arch. Androl. 1992, 29, 69–78. [Google Scholar] [CrossRef]
- Boyce, A.M.; Chong, W.H.; Shawker, T.H.; Pinto, P.A.; Linehan, W.M.; Bhattacharryya, N.; Merino, M.J.; Singer, F.R.; Collins, M.T. Characterization and management of testicular pathology in McCune-Albright syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E1782–E1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganaei, J.; Shrivastava, V.K. The effect of active immunization with gonadotropin releasing hormone conjugate (GnRH-BSA) on gonadosomatic indices (GSI) and sperm parameters in mice. Iran. J. Reprod. Med. 2008, 6, 119–123. [Google Scholar]
- Ross, M.H.; Pawlina, W. Histology; Lippincott Williams & Wilkins: Philadelphia PA, USA, 2006. [Google Scholar]
- Pourentezari, M.; Talebi, A.; Abbasi, A.; Khalili, M.A.; Mangoli, E.; Anvari, M. Effects of acrylamide on sperm parameters, chromatin quality, and the level of blood testosterone in mice. Iran. J. Reprod. Med. 2014, 12, 335. [Google Scholar] [PubMed]
- Chen, Y.; Xu, J.; Li, Y.; Han, X. Decline of sperm quality and testicular function in male mice during chronic low-dose exposure to microcystin-LR. Reprod. Toxicol. 2011, 31, 551–557. [Google Scholar] [CrossRef]
- Check, J.; Bollendorf, A.; Press, M.; Blue, T. Standard sperm morphology as a predictor of male fertility potential. Arch. Androl. 1992, 28, 39–41. [Google Scholar] [CrossRef] [Green Version]
- Rima, U.; Kimura, T.; Gebril, A.M.; Islam, M.; Bari, A.; Ferro, V.; Khan, M. Immunization of Female Mice with a Plasmid DNA Vaccine Coding EightRepeats of Gonadotrophin Releasing Hormone (Gnrh-I) and Eight T-HelperEpitopes Suppress Fertility In Vivo. J. Vaccin. 2015, 6, 1000282. [Google Scholar]
- Hull, E.M.; Meisel, R.L.; Sachs, B.D. Male sexual behavior. In Hormones, Brain and Behavior; Elsevier: Amsterdam, The Netherlands, 2002; pp. 3–137. [Google Scholar]
- Phoenix, C.H.; Goy, R.W.; Gerall, A.A.; Young, W.C. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology 1959, 65, 369–382. [Google Scholar] [CrossRef]
- Quy, R.J.; Massei, G.; Lambert, M.S.; Coats, J.; Miller, L.A.; Cowan, D.P. Effects of a GnRH vaccine on the movement and activity of free-living wild boar (Sus scrofa). Wildl. Res. 2014, 41, 185–193. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g ± SD) | Testis Weight (g ± SD) | Testis Length (cm ± SD) | Testis Width (cm ± SD) | Testis Volume | Testis Volume/Body Weight (g) | |
|---|---|---|---|---|---|---|
| Vaccinated (n = 6) | 45.18 (±8.3) | 0.033 (±0.01) | 0.49 (±0.04) | 0.34 (±0.07) | 76.84 (±11.42) | 1.72 (±0.26) |
| Control (n = 6) | 53.76 (±8.0) | 0.143 (±0.02) | 0.90 (±0.48) | 0.55 (±0.04) | 380.76 (±60.35) | 7.24 (±1.42) |
| p-value | p = 0.132 | p = 0.003 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, A.-M.; Chen, C.-C.; Hou, D.-L.; Ke, G.-M.; Lee, J.-W. Effects of a Recombinant Gonadotropin-Releasing Hormone Vaccine on Reproductive Function in Adult Male ICR Mice. Vaccines 2021, 9, 808. https://doi.org/10.3390/vaccines9080808
Chang A-M, Chen C-C, Hou D-L, Ke G-M, Lee J-W. Effects of a Recombinant Gonadotropin-Releasing Hormone Vaccine on Reproductive Function in Adult Male ICR Mice. Vaccines. 2021; 9(8):808. https://doi.org/10.3390/vaccines9080808
Chicago/Turabian StyleChang, Ai-Mei, Chen-Chih Chen, Ding-Liang Hou, Guan-Ming Ke, and Jai-Wei Lee. 2021. "Effects of a Recombinant Gonadotropin-Releasing Hormone Vaccine on Reproductive Function in Adult Male ICR Mice" Vaccines 9, no. 8: 808. https://doi.org/10.3390/vaccines9080808