
Fusion Protein of Rotavirus VP6 and SARS-CoV-2 Receptor Binding Domain Induces T Cell Responses


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning and Production of VP6-RBD Fusion Protein
2.2. Extraction and Crude Purification of VP6-RBD Fusion Protein
2.3. RV VP6, SARS-CoV-2 Proteins, and Synthetic Peptides
2.4. VP6-RBD Fusion Protein Characterization and Receptor Binding
2.5. Mice Immunization
2.6. SARS-CoV-2 and RV VP6-Specific Serological Immunoassays
2.7. ELISPOT-IFN-γ
2.8. Statistics
3. Results
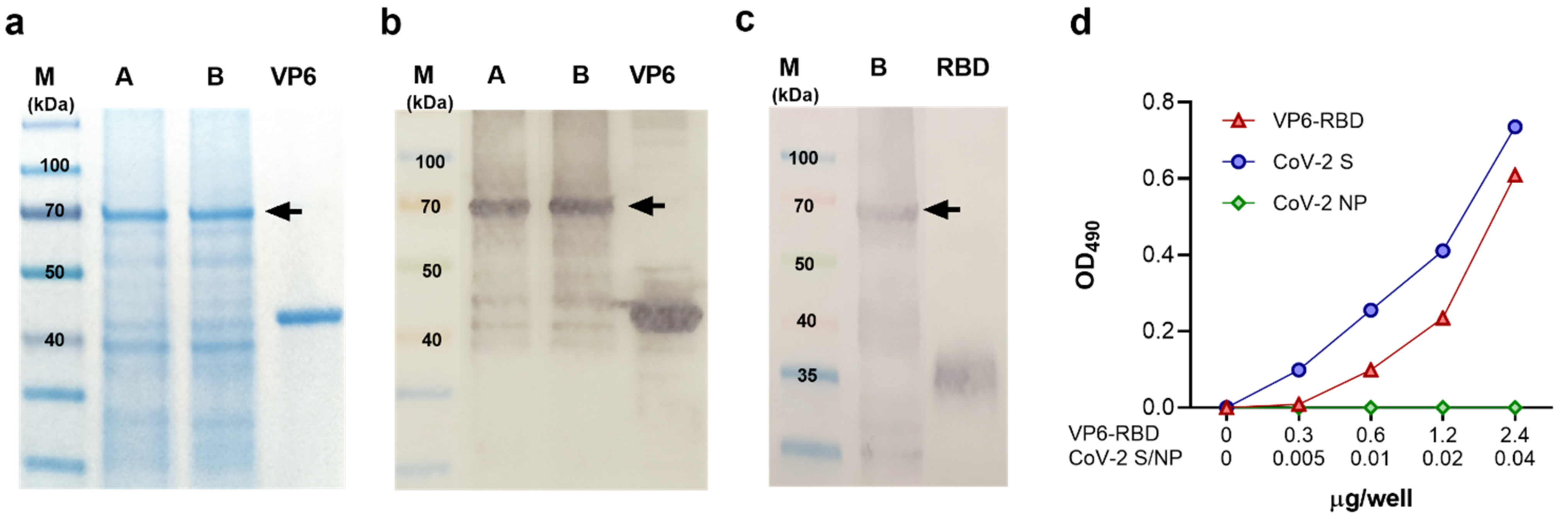
3.1. Production and Characterization of the Crude Purified VP6-RBD Fusion Protein
3.2. SARS-CoV-2 and RV VP6-Specific Antibody Responses
3.3. SARS-CoV-2-Specific T Cell Responses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO declares COVID-19 a pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [PubMed]
- COVID-19 Vaccine Tracker and Landscape. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 14 June 2021).
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Addetia, A.; Crawford, K.H.; Dingens, A.; Zhu, H.; Roychoudhury, P.; Huang, M.L.; Jerome, K.R.; Bloom, J.D.; Greninger, A.L. Neutralizing antibodies correlate with protection from SARS-CoV-2 in humans during a fishery vessel outbreak with high attack rate. J. Clin. Microbiol. 2020, 58, e02107–e02120. [Google Scholar] [CrossRef] [PubMed]
- Golob, J.L.; Lugogo, N.; Lauring, A.S.; Lok, A.S. SARS-CoV-2 vaccines: A triumph of science and collaboration. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2523. [Google Scholar] [CrossRef]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 2013, 19, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.M.; Li, C.K.; Chui, C.S.; Huang, A.K.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274. [Google Scholar] [CrossRef]
- Iqbal, H. The importance of cell-mediated immunity in COVID-19—An opinion. Med. Hypotheses 2020, 143, 110152. [Google Scholar] [CrossRef]
- Blazevic, V.; Lappalainen, S.; Nurminen, K.; Huhti, L.; Vesikari, T. Norovirus VLPs and rotavirus VP6 protein as combined vaccine for childhood gastroenteritis. Vaccine 2011, 29, 8126. [Google Scholar] [CrossRef]
- Lappalainen, S.; Pastor, A.R.; Tamminen, K.; Lopez-Guerrero, V.; Esquivel-Guadarrama, F.; Palomares, L.A.; Vesikari, T.; Blazevic, V. Immune responses elicited against rotavirus middle layer protein VP6 inhibit viral replication in vitro and in vivo. Hum. Vaccines Immunother. 2014, 10, 2039. [Google Scholar] [CrossRef]
- Pastor, A.R.; Rodriguez-Limas, W.A.; Contreras, M.A.; Esquivel, E.; Esquivel-Guadarrama, F.; Ramirez, O.T.; Palomares, L.A. The assembly conformation of rotavirus VP6 determines its protective efficacy against rotavirus challenge in mice. Vaccine 2014, 32, 2874–2877. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.L.; Vaysburd, M.; Wing, M.; Foss, S.; Andersen, J.T.; O’Connell, K.; Mayes, K.; Higginson, K.; Iturriza-Gomara, M.; Desselberger, U.; et al. Intracellular neutralisation of rotavirus by VP6-specific IgG. PLoS Pathog. 2020, 16, e1008732. [Google Scholar] [CrossRef]
- Malm, M.; Tamminen, K.; Lappalainen, S.; Vesikari, T.; Blazevic, V. Rotavirus recombinant VP6 nanotubes act as an immunomodulator and delivery vehicle for norovirus virus-like particles. J. Immunol. Res. 2016, 2016, 9171632. [Google Scholar] [CrossRef] [Green Version]
- Tamminen, K.; Heinimaki, S.; Vesikari, T.; Blazevic, V. Rotavirus VP6 adjuvant effect on norovirus GII.4 virus-like particle uptake and presentation by bone marrow-derived dendritic cells in vitro and in vivo. J. Immunol. Res. 2020, 2020, 3194704. [Google Scholar] [CrossRef]
- Rodriguez, M.; Wood, C.; Sanchez-Lopez, R.; Castro-Acosta, R.M.; Ramirez, O.T.; Palomares, L.A. Understanding internalization of rotavirus VP6 nanotubes by cells: Towards a recombinant vaccine. Arch. Virol. 2014, 159, 1005–1015. [Google Scholar] [CrossRef]
- Tamminen, K.; Heinimaki, S.; Grohn, S.; Blazevic, V. Internalization and antigen presentation by mouse dendritic cells of rotavirus VP6 preparations differing in nanostructure. Mol. Immunol. 2020, 123, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Peralta, A.; Molinari, P.; Taboga, O. Chimeric recombinant rotavirus-like particles as a vehicle for the display of heterologous epitopes. Virol. J. 2009, 6, 192. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Zhao, B.; Pan, X.; Wen, Y.; Chen, Y. A new rotavirus VP6-based foreign epitope presenting vector and immunoreactivity of VP4 epitope chimeric proteins. Viral Immunol. 2014, 27, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Grohn, S.; Heinimaki, S.; Tamminen, K.; Blazevic, V. Expression of influenza A virus-derived peptides on a rotavirus VP6-based delivery platform. Arch. Virol. 2021, 166, 213–217. [Google Scholar] [CrossRef]
- PANGO Lineages. Available online: https://cov-lineages.org/ (accessed on 14 June 2021).
- O’Shaughnessy, L.; Doyle, S. Purification of proteins from baculovirus-infected insect cells. Methods Mol. Biol. 2011, 681, 295–309. [Google Scholar] [PubMed]
- Lappalainen, S.; Tamminen, K.; Vesikari, T.; Blazevic, V. Comparative immunogenicity in mice of rotavirus VP6 tubular structures and virus-like particles. Hum. Vaccines Immunother. 2013, 9, 1991–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, A.A.; Gnanapragasam, P.N.P.; Lee, Y.E.; Hoffman, P.R.; Ou, S.; Kakutani, L.M.; Keeffe, J.R.; Wu, H.J.; Howarth, M.; West, A.P.; et al. Mosaic nanoparticles elicit cross-reactive immune responses to zoonotic coronaviruses in mice. Science 2021, 371, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Tamminen, K.; Huhti, L.; Koho, T.; Lappalainen, S.; Hytonen, V.P.; Vesikari, T.; Blazevic, V. A comparison of immunogenicity of norovirus GII-4 virus-like particles and P-particles. Immunology 2012, 135, 89. [Google Scholar] [CrossRef] [PubMed]
- Foged, C. Subunit vaccines of the future: The need for safe, customized and optimized particulate delivery systems. Ther. Deliv. 2011, 2, 1057–1077. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, N.H.; Frosig, T.M.; Jakobsen, J.T.; Buus, S.; Andersen, M.H.; Jungersen, G. Low antigen dose formulated in CAF09 adjuvant Favours a cytotoxic T-cell response following intraperitoneal immunization in Gottingen minipigs. Vaccine 2017, 35, 5629–5636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Poon, V.K.; Chan, C.C.; Lee, A.C.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the clinical and pathological manifestations of coronavirus disease 2019 (COVID-19) in a golden Syrian hamster model: Implications for disease pathogenesis and transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef]
- Zaira, R.; Ammad, F.; Faraz, B.M. Scouting the receptor binding domain of COVID-19: A comprehensive immunoinformatics inquisition. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Lee, W.S.; Wheatley, A.K.; Kent, S.J.; DeKosky, B.J. Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies. Nat. Microbiol. 2020, 5, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Bolles, M.; Deming, D.; Long, K.; Agnihothram, S.; Whitmore, A.; Ferris, M.; Funkhouser, W.; Gralinski, L.; Totura, A.; Heise, M.; et al. A double-inactivated severe acute respiratory syndrome coronavirus vaccine provides incomplete protection in mice and induces increased eosinophilic proinflammatory pulmonary response upon challenge. J. Virol. 2011, 85, 12201–12215. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.T.; Sbrana, E.; Iwata-Yoshikawa, N.; Newman, P.C.; Garron, T.; Atmar, R.L.; Peters, C.J.; Couch, R.B. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS ONE 2012, 7, e35421. [Google Scholar]
- Morgan, J.; Muskat, K.; Tippalagama, R.; Sette, A.; Burel, J.; Arlehamn, C.S.L. Classical CD4 T cells as the cornerstone of antimycobacterial immunity. Immunol. Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.H.; Simmons, C.; Jong, M.D.D.; Chau, N.V.; Schumacher, R.; Peng, Y.C.; McMichael, A.J.; Farrar, J.J.; Smith, G.L.; Townsend, A.R.; et al. Memory T cells established by seasonal human influenza A infection cross-react with avian influenza A (H5N1) in healthy individuals. J. Clin. Investig. 2008, 118, 3478–3490. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, W.S.; Emerson, R.O.; Lindau, P.; Vignali, M.; Snyder, T.M.; Desmarais, C.; Sanders, C.; Utsugi, H.; Warren, E.H.; McElrath, J.; et al. Dynamics of the cytotoxic T cell response to a model of acute viral infection. J. Virol. 2015, 89, 4517–4526. [Google Scholar] [CrossRef] [Green Version]
- Mothe, B.; Llano, A.; Ibarrondo, J.; Daniels, M.; Miranda, C.; Zamarreno, J.; Bach, V.; Zuniga, R.; Perez-Alvarez, S.; Berger, C.T.; et al. Definition of the viral targets of protective HIV-1-specific T cell responses. J. Transl. Med. 2011, 9, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamminen, K.; Heinimäki, S.; Gröhn, S.; Blazevic, V. Fusion Protein of Rotavirus VP6 and SARS-CoV-2 Receptor Binding Domain Induces T Cell Responses. Vaccines 2021, 9, 733. https://doi.org/10.3390/vaccines9070733
Tamminen K, Heinimäki S, Gröhn S, Blazevic V. Fusion Protein of Rotavirus VP6 and SARS-CoV-2 Receptor Binding Domain Induces T Cell Responses. Vaccines. 2021; 9(7):733. https://doi.org/10.3390/vaccines9070733
Chicago/Turabian StyleTamminen, Kirsi, Suvi Heinimäki, Stina Gröhn, and Vesna Blazevic. 2021. "Fusion Protein of Rotavirus VP6 and SARS-CoV-2 Receptor Binding Domain Induces T Cell Responses" Vaccines 9, no. 7: 733. https://doi.org/10.3390/vaccines9070733