
Optimization of Vero Cells Grown on a Polymer Fiber Carrier in a Disposable Bioreactor for Inactivated Coxsackievirus A16 Vaccine Development

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and the Generation of the Cell Bank
2.2. Plaque Purification and Virus Seeds Generation
2.3. Determination of the Viral Titer and Residual BSA in the Harvest Supernatant
2.4. Vero Cell Culture in the Microbioreactor
2.5. Micro Bioreactor Optimization of the Yield of the Virus
2.6. CVA16 Production Using Different Strategies of Medium Supplementation
2.7. Cell Expansion and Bioreactor Inoculation
2.8. ACPB and NBS-Bioflo310 Culture
2.9. Inactivated CVA16 Preparation and Immunization of Mice
3. Results
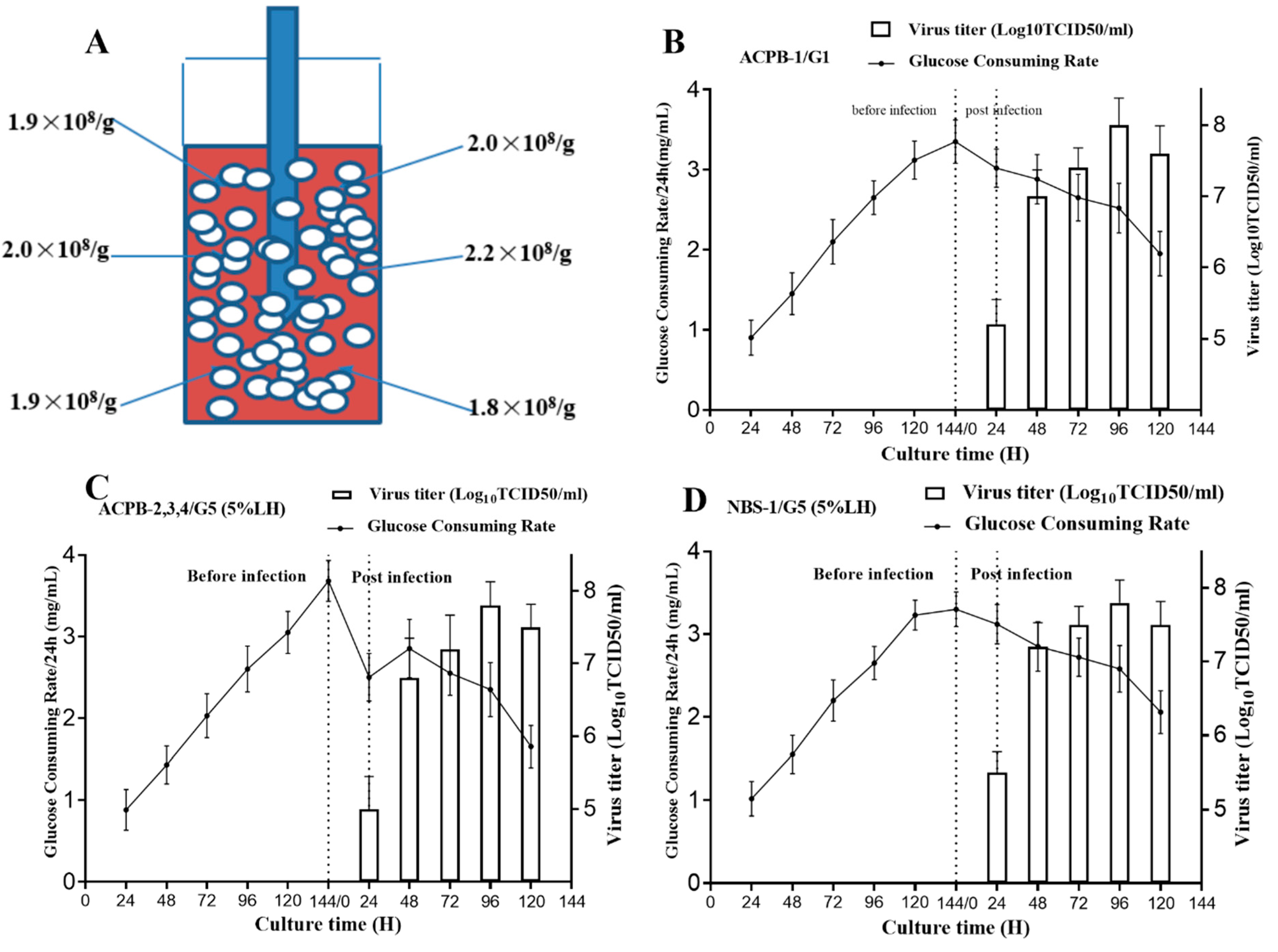
3.1. GCR Determination in the Mini Bioreactor
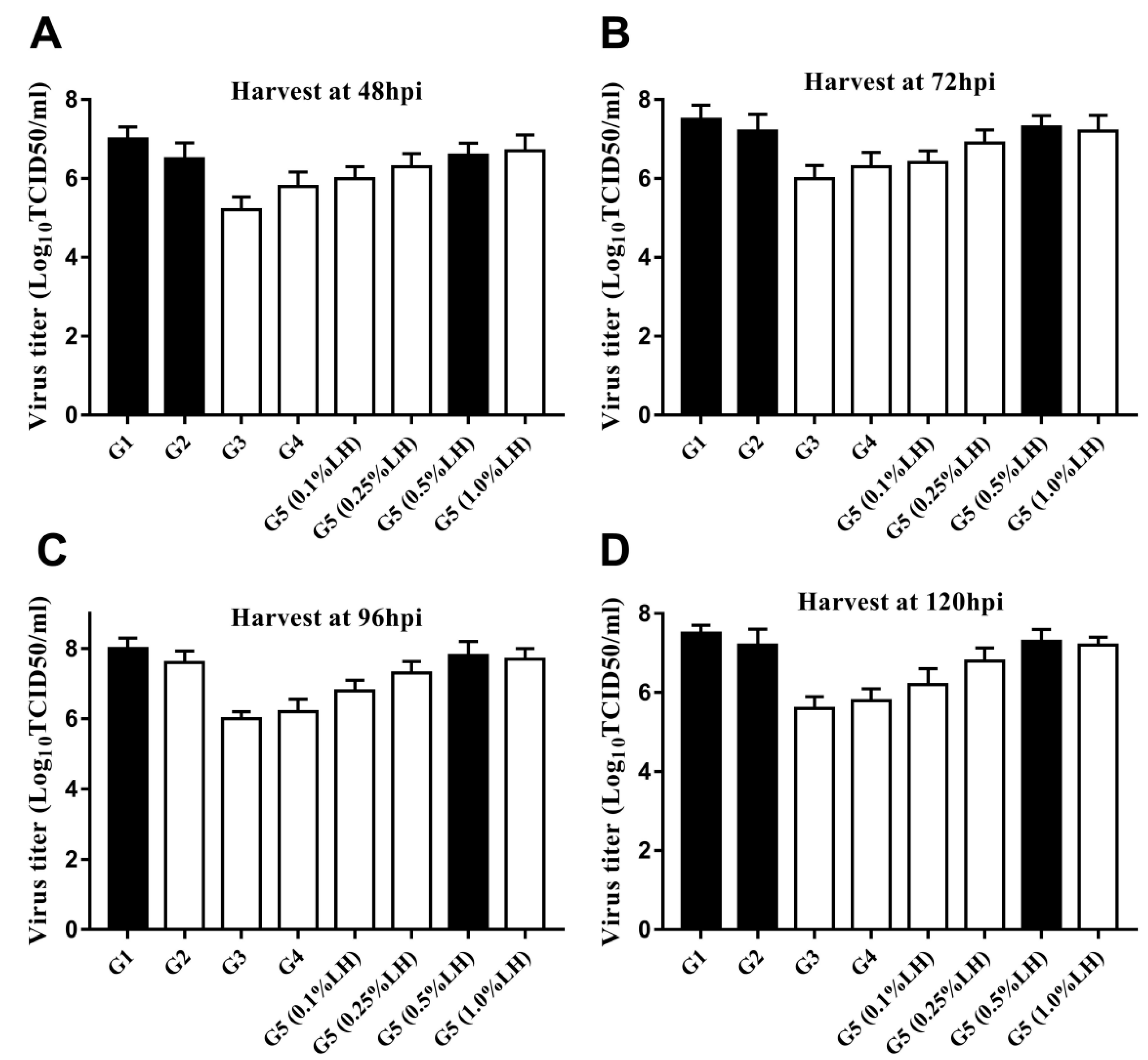
3.2. Infection Parameter Optimization in the Microbioreactors
3.3. Dependence of CVA16 Viral Yield on Different Culture Strategies and Medium Supplements
3.4. ACPB and NBS-Bioflo310 Culture
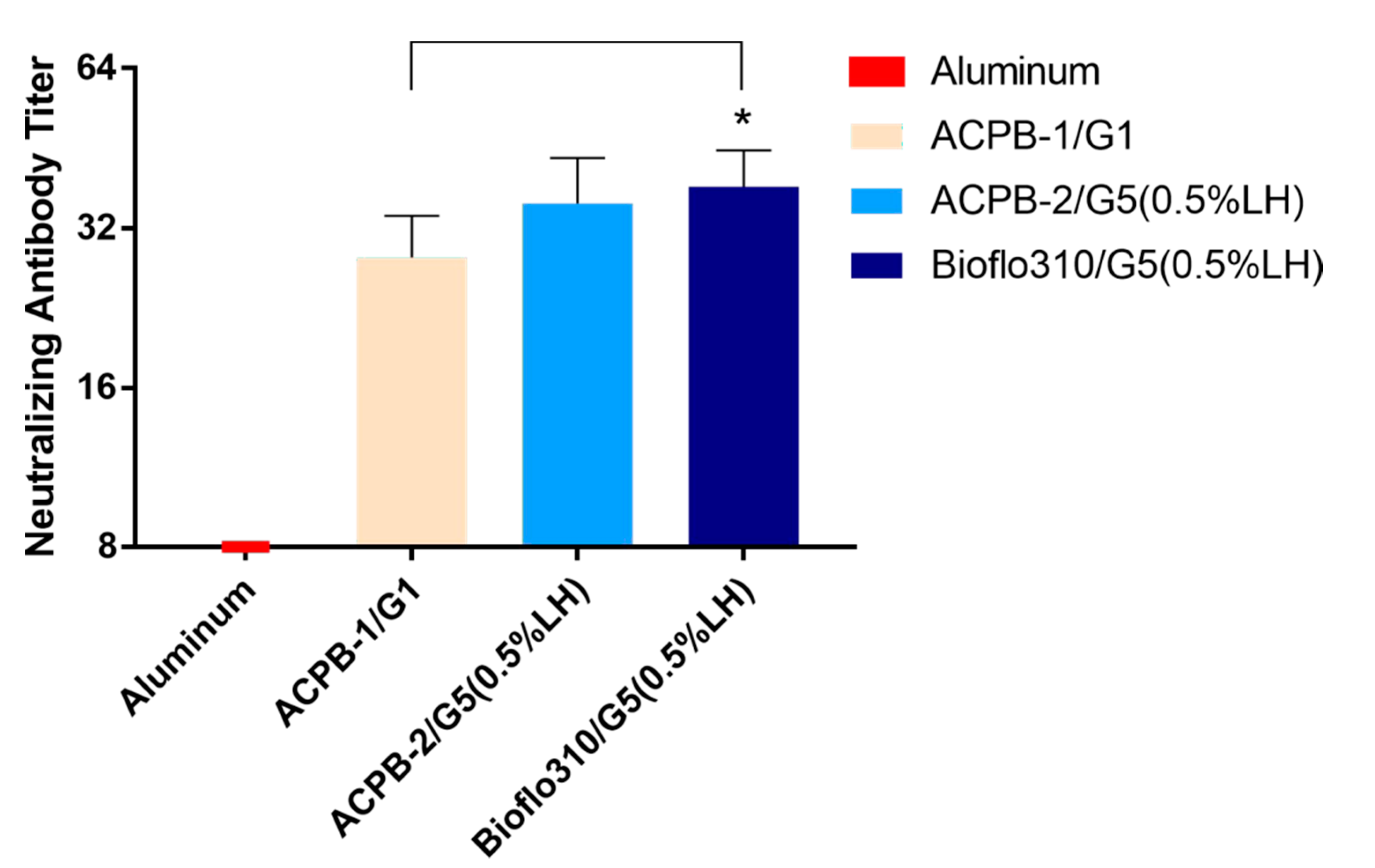
3.5. Formalin-Inactivated CVA16 Immunogenicity in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yi, E.J.; Shin, Y.J.; Kim, J.H.; Kim, T.G.; Chang, S.Y. Enterovirus 71 infection and vaccines. Clin. Exp. Vaccine Res. 2017, 6, 4–14. [Google Scholar] [CrossRef]
- Xiao, X.; Gasparrini, A.; Huang, J.; Liao, Q.; Liu, F.; Yin, F.; Yu, H.; Li, X. The exposure-response relationship between temperature and childhood hand, foot and mouth disease: A multicity study from mainland China. Environ. Int. 2017, 100, 102–109. [Google Scholar] [CrossRef]
- Xiao, X.; Liao, Q.; Kenward, M.G.; Zheng, Y.; Huang, J.; Yin, F.; Yu, H.; Li, X. Comparisons between mild and severe cases of hand, foot and mouth disease in temporal trends: A comparative time series study from mainland China. BMC Public Health 2016, 16, 1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.C.; Kou, Z.Q.; Bai, Y.J.; Cong, X.; Wang, L.H.; Li, C.; Zhao, L.; Yu, X.J.; Wang, Z.Y.; Wen, H.L. Epidemiological Research on Hand, Foot, and Mouth Disease in Mainland China. Viruses 2015, 7, 6400–6411. [Google Scholar] [CrossRef] [Green Version]
- Mizuta, K.; Aoki, Y.; Matoba, Y.; Yahagi, K.; Itagaki, T.; Katsushima, F.; Katsushima, Y.; Ito, S.; Hongo, S.; Matsuzaki, Y. Molecular epidemiology of enterovirus 71 strains isolated from children in Yamagata, Japan, between 1990 and 2013. J. Med. Microbiol. 2014, 63, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.G.; Yi, H.X.; Shu, J.; Wang, X.J.; Wu, X.J.; Yu, L.H. Clinical and epidemiological characteristics of adult hand, foot, and mouth disease in northern Zhejiang, China, May 2008–November 2013. BMC Infect. Dis. 2014, 14, 251. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Shi, J.; Huang, X.; Liu, F.; Cai, Y.; Lan, K.; Huang, Z. A murine model of coxsackievirus A16 infection for anti-viral evaluation. Antivir. Res. 2014, 105, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liu, Q.; Huang, X.; Li, D.; Ku, Z.; Zhang, Y.; Huang, Z. Active immunization with a Coxsackievirus A16 experimental inactivated vaccine induces neutralizing antibodies and protects mice against lethal infection. Vaccine 2013, 31, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.W.; Koh, B.K.; Chan, K.P.; Chua, L.T.; James, L.; Goh, K.T. Epidemiology and control of hand, foot and mouth disease in Singapore, 2001–2007. Ann. Acad. Med. Singap. 2009, 38, 106–112. [Google Scholar] [PubMed]
- Cai, Y.; Ku, Z.; Liu, Q.; Leng, Q.; Huang, Z. A combination vaccine comprising of inactivated enterovirus 71 and coxsackievirus A16 elicits balanced protective immunity against both viruses. Vaccine 2014, 32, 2406–2412. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ku, Z.; Cai, Y.; Sun, B.; Leng, Q.; Huang, Z. Detection, characterization and quantitation of coxsackievirus A16 using polyclonal antibodies against recombinant capsid subunit proteins. J. Virol. Methods 2011, 173, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Chatproedprai, S.; Theanboonlers, A.; Korkong, S.; Thongmee, C.; Wananukul, S.; Poovorawan, Y. Clinical and molecular characterization of hand-foot-and-mouth disease in Thailand, 2008–2009. Jpn. J. Infect. Dis. 2010, 63, 229–233. [Google Scholar] [PubMed]
- Tu, P.V.; Thao, N.T.; Perera, D.; Huu, T.K.; Tien, N.T.; Thuong, T.C.; How, O.M.; Cardosa, M.J.; McMinn, P.C. Epidemiologic and virologic investigation of hand, foot, and mouth disease, southern Vietnam, 2005. Emerg. Infect. Dis. 2007, 13, 1733–1741. [Google Scholar] [CrossRef]
- Li, L.; He, Y.; Yang, H.; Zhu, J.; Xu, X.; Dong, J.; Zhu, Y.; Jin, Q. Genetic characteristics of human enterovirus 71 and coxsackievirus A16 circulating from 1999 to 2004 in Shenzhen, People’s Republic of China. J. Clin. Microbiol. 2005, 43, 3835–3839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoya, M.; Kawasaki, Y.; Sato, M.; Honzumi, K.; Hayashi, A.; Hiroshima, T.; Ishiko, H.; Kato, K.; Suzuki, H. Genetic diversity of coxsackievirus A16 associated with hand, foot, and mouth disease epidemics in Japan from 1983 to 2003. J. Clin. Microbiol. 2007, 45, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tan, X.J.; Wang, H.Y.; Yan, D.M.; Zhu, S.L.; Wang, D.Y.; Ji, F.; Wang, X.J.; Gao, Y.J.; Chen, L.; et al. An outbreak of hand, foot, and mouth disease associated with subgenotype C4 of human enterovirus 71 in Shandong, China. J. Clin. Virol. 2009, 44, 262–267. [Google Scholar] [CrossRef]
- McMinn, P.C. An overview of the evolution of enterovirus 71 and its clinical and public health significance. FEMS Microbiol. Rev. 2002, 26, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.Y.; Lin, T.Y.; Huang, Y.C.; Tsao, K.C.; Shih, S.R.; Kuo, M.L.; Ning, H.C.; Chung, P.W.; Kang, C.M. Comparison of enterovirus 71 and coxsackie-virus A16 clinical illnesses during the Taiwan enterovirus epidemic, 1998. Pediatr. Infect. Dis. J. 1999, 18, 1092–1096. [Google Scholar] [CrossRef]
- Iwai, M.; Masaki, A.; Hasegawa, S.; Obara, M.; Horimoto, E.; Nakamura, K.; Tanaka, Y.; Endo, K.; Tanaka, K.; Ueda, J.; et al. Genetic changes of coxsackievirus A16 and enterovirus 71 isolated from hand, foot, and mouth disease patients in Toyama, Japan between 1981 and 2007. Jpn. J. Infect. Dis. 2009, 62, 254–259. [Google Scholar]
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef]
- Wang, C.Y.; Li Lu, F.; Wu, M.H.; Lee, C.Y.; Huang, L.M. Fatal coxsackievirus A16 infection. Pediatr. Infect. Dis. J. 2004, 23, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.T., Jr.; Landing, B.H.; Lennette, E.H.; Mc, A.R. Fatal infection in an infant associated with Coxsackie virus group A, type 16. N. Engl. J. Med. 1963, 268, 1041–1044. [Google Scholar] [CrossRef]
- Xu, W.; Liu, C.F.; Yan, L.; Li, J.J.; Wang, L.J.; Qi, Y.; Cheng, R.B.; Xiong, X.Y. Distribution of enteroviruses in hospitalized children with hand, foot and mouth disease and relationship between pathogens and nervous system complications. Virol. J. 2012, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.C.; Liu, C.C.; Lian, W.C. Optimization of microcarrier cell culture process for the inactivated enterovirus type 71 vaccine development. Vaccine 2004, 22, 3858–3864. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Lian, W.C.; Butler, M.; Wu, S.C. High immunogenic enterovirus 71 strain and its production using serum-free microcarrier Vero cell culture. Vaccine 2007, 25, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Yu, X.; Kong, W.; Sun, S.; Yang, P.; Zhu, C.; Zhang, H.; Wu, Y.; Chen, Y.; Shi, Y.; et al. Production of influenza H1N1 vaccine from MDCK cells using a novel disposable packed-bed bioreactor. Appl. Microbiol. Biotechnol. 2013, 97, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Shi, M.; Song, Y.; Bao, L.; Yang, W.; Zhang, X.; Ruan, M.; Rishton, G.; Joudi, A.; Teng, Y.; et al. A Single-Use, Scalable Perfusion Bioreactor System. Bioprocess. Int. 2009, 7, 46–54. [Google Scholar]
- Chen, K.D.; Wu, X.X.; Yu, D.S.; Ou, H.L.; Li, Y.H.; Zhou, Y.Q.; Li, L.J. Process optimization for the rapid production of adenoviral vectors for clinical trials in a disposable bioreactor system. Appl. Microbiol. Biotechnol. 2018, 102, 6469–6477. [Google Scholar] [CrossRef]
- Butler, M.; Burgener, A.; Patrick, M.; Berry, M.; Moffatt, D.; Huzel, N.; Barnabé, N.; Coombs, K. Application of a serum-free medium for the growth of Vero cells and the production of reovirus. Biotechnol. Prog. 2000, 16, 854–858. [Google Scholar] [CrossRef]
- Fishbein, D.B.; Yenne, K.M.; Dreesen, D.W.; Teplis, C.F.; Mehta, N.; Briggs, D.J. Risk factors for systemic hypersensitivity reactions after booster vaccinations with human diploid cell rabies vaccine: A nationwide prospective study. Vaccine 1993, 11, 1390–1394. [Google Scholar] [CrossRef]
- Hu, A.Y.; Tseng, Y.F.; Weng, T.C.; Liao, C.C.; Wu, J.; Chou, A.H.; Chao, H.J.; Gu, A.; Chen, J.; Lin, S.C.; et al. Production of inactivated influenza H5N1 vaccines from MDCK cells in serum-free medium. PLoS ONE 2011, 6, e14578. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating 50 percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Chong, P.; Guo, M.S.; Lin, F.H.; Hsiao, K.N.; Weng, S.Y.; Chou, A.H.; Wang, J.R.; Hsieh, S.Y.; Su, I.J.; Liu, C.C. Immunological and biochemical characterization of coxsackie virus A16 viral particles. PLoS ONE 2012, 7, e49973. [Google Scholar] [CrossRef]
- Luo, S.T.; Chiang, P.S.; Chao, A.S.; Liou, G.Y.; Lin, R.; Lin, T.Y.; Lee, M.S. Enterovirus 71 maternal antibodies in infants, Taiwan. Emerg. Infect. Dis. 2009, 15, 581–584. [Google Scholar] [CrossRef]
- Chou, A.H.; Liu, C.C.; Chang, C.P.; Guo, M.S.; Hsieh, S.Y.; Yang, W.H.; Chao, H.J.; Wu, C.L.; Huang, J.L.; Lee, M.S.; et al. Pilot scale production of highly efficacious and stable enterovirus 71 vaccine candidates. PLoS ONE 2012, 7, e34834. [Google Scholar] [CrossRef]
- Kaptein, L.C.; Greijer, A.E.; Valerio, D.; van Beusechem, V.W. Optimized conditions for the production of recombinant amphotropic retroviral vector preparations. Gene Ther. 1997, 4, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Doux, J.M.; Davis, H.E.; Morgan, J.R.; Yarmush, M.L. Kinetics of retrovirus production and decay. Biotechnol. Bioeng. 1999, 63, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Bao, L.L.; Mao, H.Y.; Wang, L.; Xu, K.W.; Yang, M.N.; Li, Y.J.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.T.; Huang, B.Y.; Deng, W.; Quan, Y.; Wang, W.L.; Xu, W.B.; Zhao, Y.X.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source/Harvest Time | 48 hpi | 72 hpi | 96 hpi | 120 hpi |
|---|---|---|---|---|
| ACPB-1/G1 | 8.0 × 106 ng/mL | 7.6 × 106 ng/mL | 8.6 × 106 ng/mL | 8.8 × 106 ng/mL |
| ACPB-2/G5 (0.5% LH) | 280 ng/mL | 86 ng/mL | 16 ng/mL | 5.6 ng/mL |
| Bioflo310/G5 (0.5% LH) | 265 ng/mL | 35 ng/mL | 3.2 ng/mL | undetectable |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Li, C.; Wang, Y.; Shen, Z.; Guo, Y.; Li, X.; Zhang, Y. Optimization of Vero Cells Grown on a Polymer Fiber Carrier in a Disposable Bioreactor for Inactivated Coxsackievirus A16 Vaccine Development. Vaccines 2021, 9, 613. https://doi.org/10.3390/vaccines9060613
Chen K, Li C, Wang Y, Shen Z, Guo Y, Li X, Zhang Y. Optimization of Vero Cells Grown on a Polymer Fiber Carrier in a Disposable Bioreactor for Inactivated Coxsackievirus A16 Vaccine Development. Vaccines. 2021; 9(6):613. https://doi.org/10.3390/vaccines9060613
Chicago/Turabian StyleChen, Keda, Chaonan Li, Ying Wang, Zhenwei Shen, Yikai Guo, Xiaoping Li, and Yanjun Zhang. 2021. "Optimization of Vero Cells Grown on a Polymer Fiber Carrier in a Disposable Bioreactor for Inactivated Coxsackievirus A16 Vaccine Development" Vaccines 9, no. 6: 613. https://doi.org/10.3390/vaccines9060613