
Effect of Preexisting Immunity to Tetanus Toxoid on the Efficacy of Tetanus Toxoid-Conjugated Heroin Vaccine in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Coupling Procedure
2.2.1. TT Conjugate Antigen Synthesis
2.2.2. BSA-Hapten Conjugate Synthesis
2.3. Vaccine Formulation and Immunization
2.3.1. Vaccine Formulation
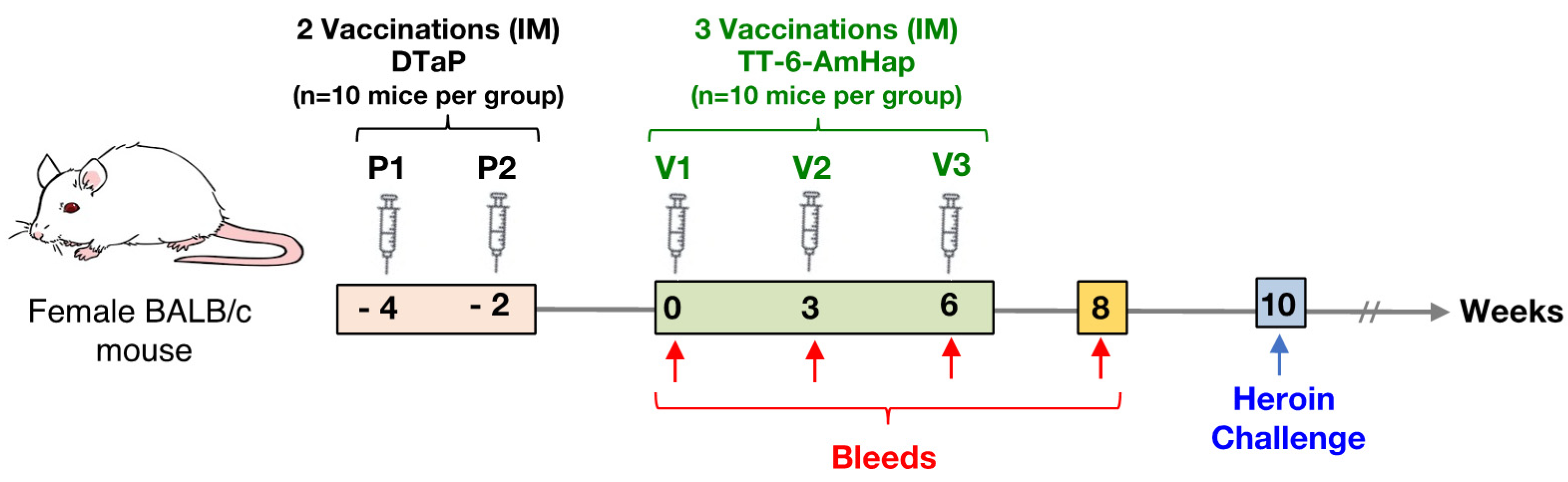
2.3.2. Animal Studies
2.4. Behavioral Assays
2.4.1. Hot Plate Test
2.4.2. Locomotion Test
2.5. Immunogenicity Studies
Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Data Analysis
3. Results
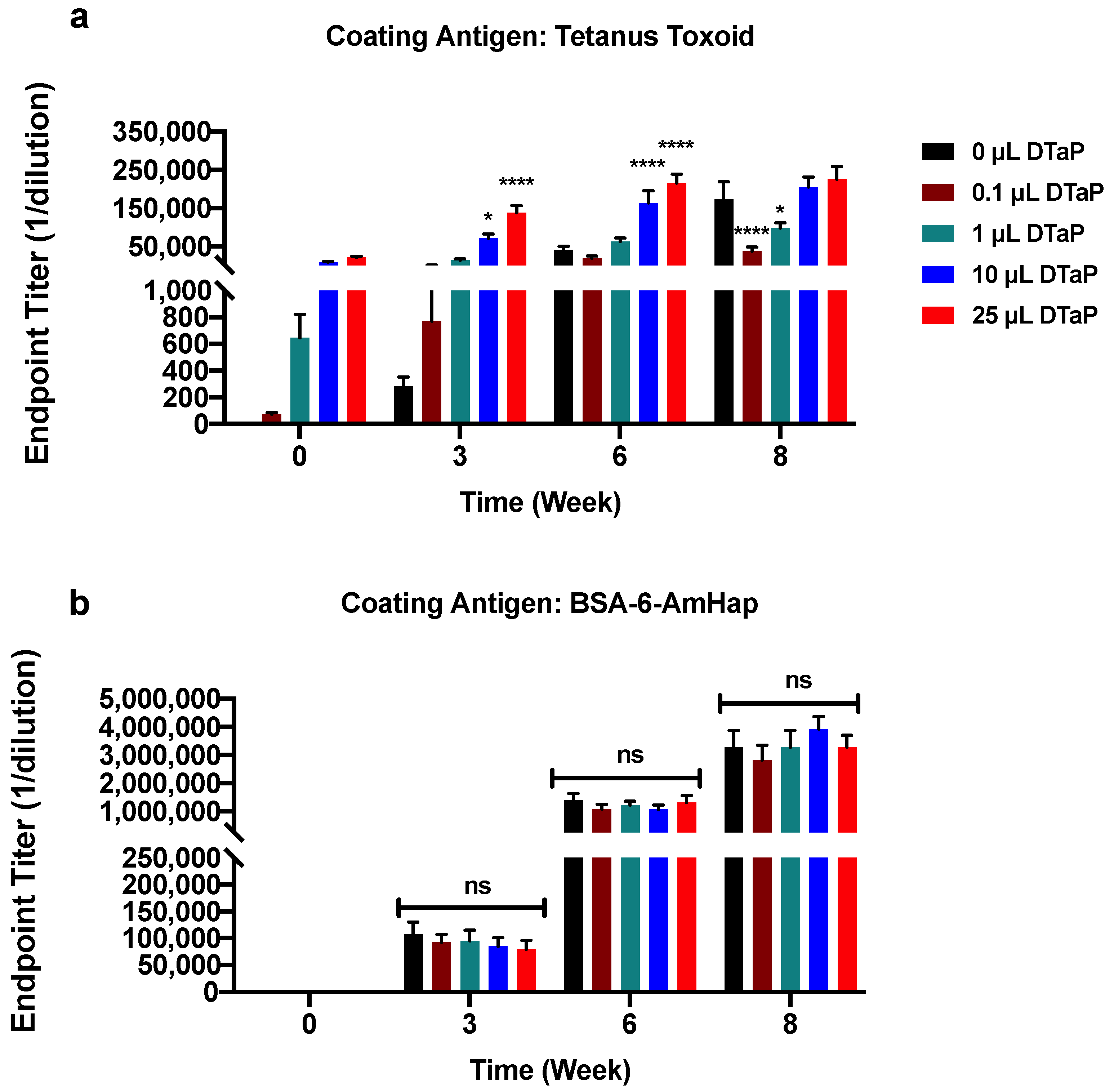
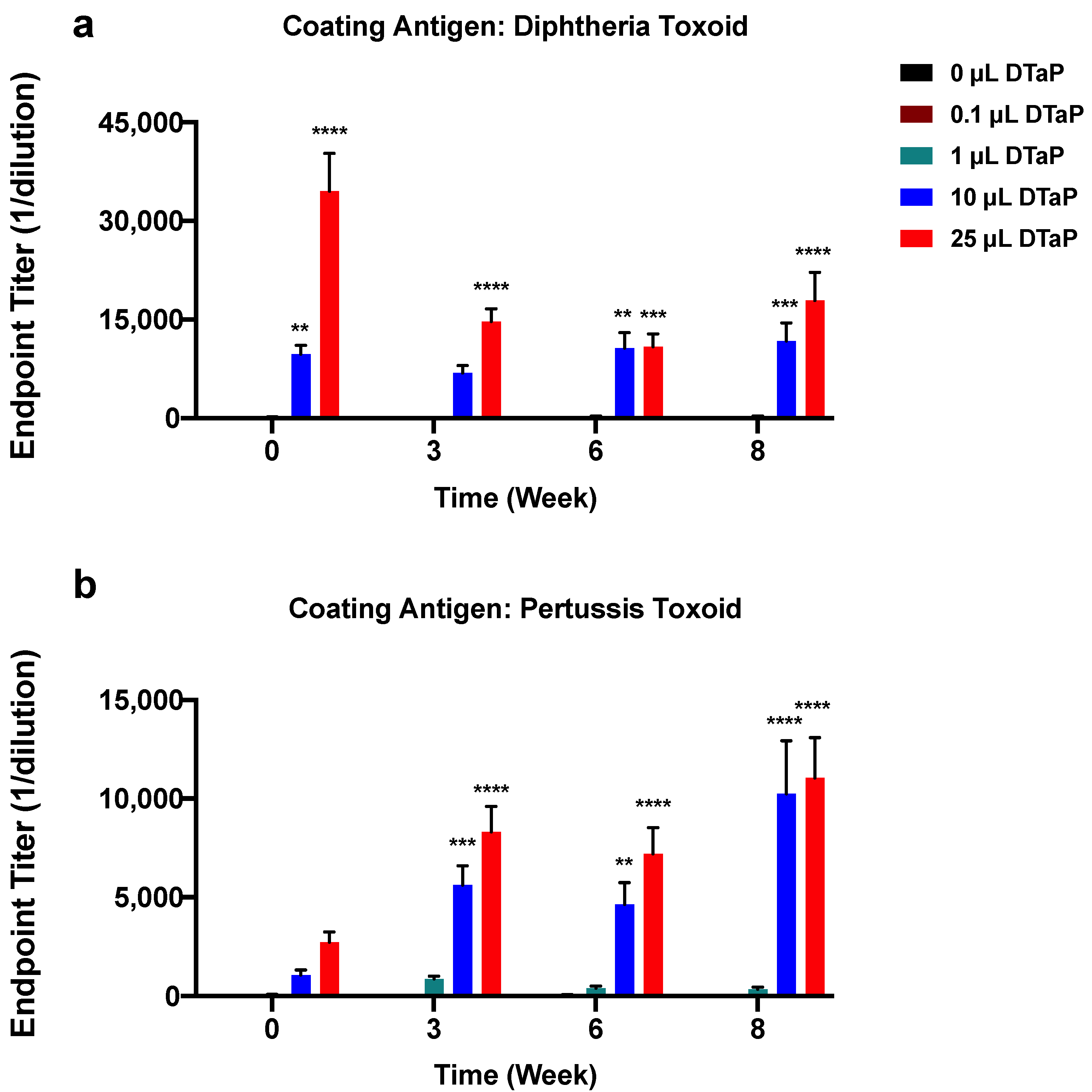
3.1. Effect of DTaP Immunization on the Immunogenicity of the TT-6-AmHap Vaccine
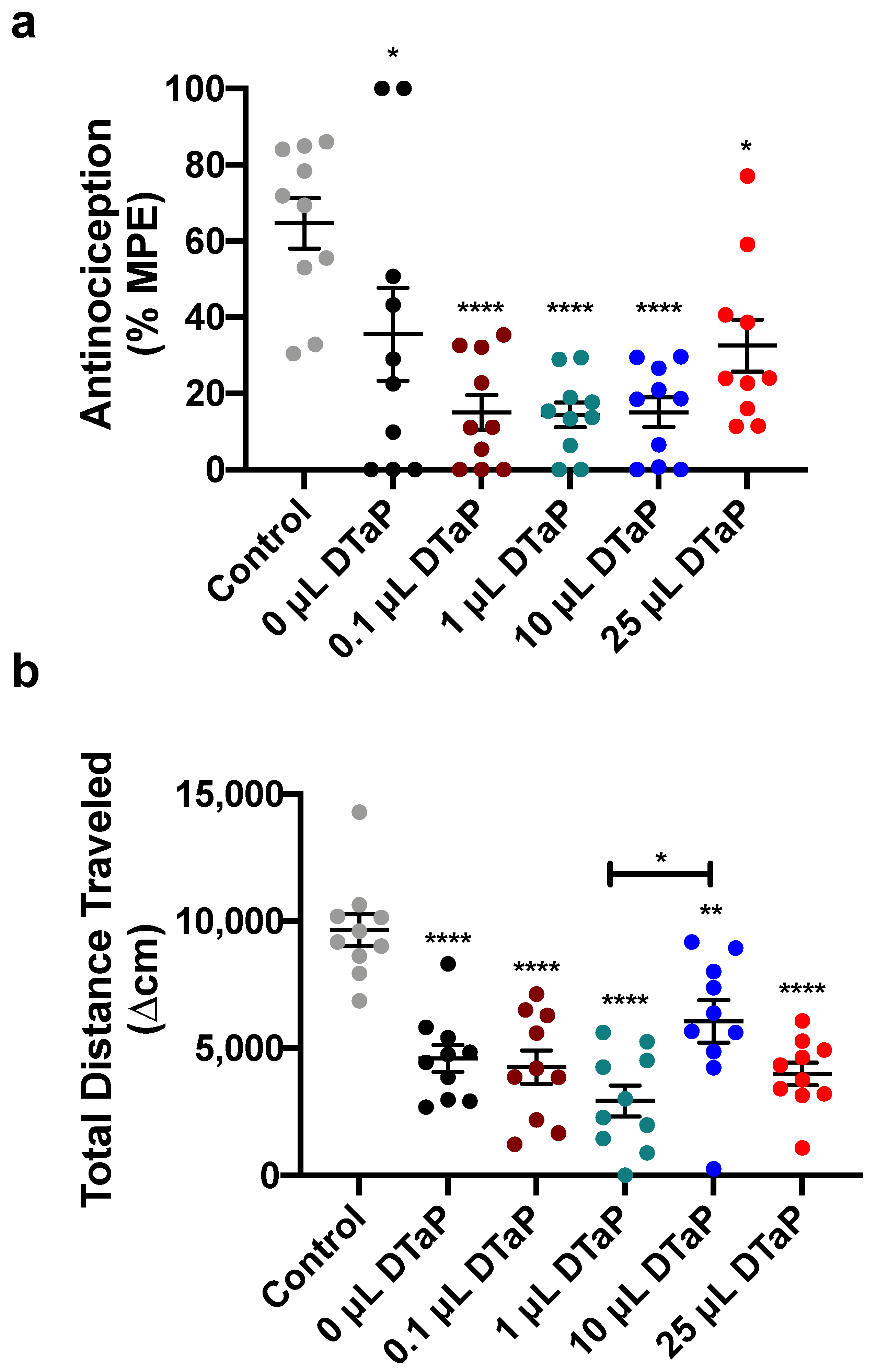
3.2. Efficacy of TT-6-AmHap Heroin Vaccine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scholl, L.; Seth, P.; Kariisa, M.; Wilson, N.; Baldwin, G. Drug and Opioid-Involved Overdose Deaths—United States, 2013–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1419–1427. [Google Scholar] [CrossRef]
- Wilson, N.; Kariisa, M.; Seth, P.; Smith, H.; Davis, N.L. Drug and Opioid-Involved Overdose Deaths—United States, 2017–2018. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Woodcock, J.; Compton, W.M.; Throckmorton, D.C.; Skolnick, P.; Hertz, S.; Wargo, E.M. Medication development in opioid addiction: Meaningful clinical end points. Sci. Transl. Med. 2018, 10, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.H.; Blake, M.S. Effect of Increased CRM 197 Carrier Protein Dose on Meningococcal C Bactericidal Antibody Response. Clin. Vaccine Immunol. 2012, 19, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Tontini, M.; Berti, F.; Romano, M.; Proietti, D.; Zambonelli, C.; Bottomley, M.; De Gregorio, E.; Del Giudice, G.; Rappuoli, R.; Costantino, P.; et al. Comparison of CRM197, diphtheria toxoid and tetanus toxoid as protein carriers for meningococcal glycoconjugate vaccines. Vaccine 2013, 31, 4827–4833. [Google Scholar] [CrossRef] [PubMed]
- Knuf, M.; Kowalzik, F.; Kieninger, D. Comparative effects of carrier proteins on vaccine-induced immune response. Vaccine 2011, 29, 4881–4890. [Google Scholar] [CrossRef] [PubMed]
- Pöllabauer, E.M.; Petermann, R.; Ehrlich, H.J. The influence of carrier protein on the immunogenicity of simultaneously administered conjugate vaccines in infants. Vaccine 2009, 27, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Pobre, K.; Tashani, M.; Ridda, I.; Rashid, H.; Wong, M.; Booy, R. Carrier priming or suppression: Understanding carrier priming enhancement of anti-polysaccharide antibody response to conjugate vaccines. Vaccine 2014, 32, 1423–1430. [Google Scholar] [CrossRef]
- Pecetta, S.; Tontini, M.; Faenzi, E.; Cioncada, R.; Proietti, D.; Seubert, A.; Nuti, S.; Berti, F.; Romano, M. Carrier priming effect of CRM 197 is related to an enhanced B and T cell activation in meningococcal serogroup A conjugate vaccination. Immunological comparison between CRM 197 and diphtheria toxoid. Vaccine 2016, 34, 2334–2341. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Wiesel, M.; Dietmeier, K.; Zabel, F.; Gatto, D.; Saudan, P.; Bachmann, M.F. Carrier induced epitopic suppression of antibody responses induced by virus-like particles is a dynamic phenomenon caused by carrier-specific antibodies. Vaccine 2010, 28, 5503–5512. [Google Scholar] [CrossRef] [PubMed]
- Booy, R.; Heath, P.T.; Slack, M.P.; Begg, N.; Moxon, E.R. Vaccine failures after primary immunisation with Haemophilus influenzae type-b conjugate vaccine without booster. Lancet 1997, 349, 1197–1202. [Google Scholar] [CrossRef]
- Lieberman, J.M.; Greenberg, D.P.; Wong, V.K.; Partridge, S.; Chang, S.-J.; Chiu, C.-Y.; Ward, J.I. Effect of neonatal immunization with diphtheria and tetanus toxoids on antibody responses to Haemophilus influenzae type b conjugate vaccines. J. Pediatr. 1995, 126, 198–205. [Google Scholar] [CrossRef]
- Barington, T.; Kristensen, K.; Henrichsen, J.; Heilmann, C. Influence of prevaccination immunity on the human B-lymphocyte response to a Haemophilus influenzae type b conjugate vaccine. Infect. Immun. 1991, 59, 1057–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barington, T.; Skettrup, M.; Juul, L.; Heilmann, C. Non-epitope-specific suppression of the antibody response to Haemophilus influenzae type b conjugate vaccines by preimmunization with vaccine components. Infect. Immun. 1993, 61, 432–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalah, R.; Torres, O.B.; Mayorov, A.V.; Li, F.; Antoline, J.F.G.; Jacobson, A.E.; Rice, K.C.; Deschamps, J.R.; Beck, Z.; Alving, C.R.; et al. Efficacy, but Not Antibody Titer or Affinity, of a Heroin Hapten Conjugate Vaccine Correlates with Increasing Hapten Densities on Tetanus Toxoid, but Not on CRM 197 Carriers. Bioconjug. Chem. 2015, 26, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Torres, O.B.; Jalah, R.; Rice, K.C.; Li, F.; Antoline, J.F.G.; Iyer, M.R.; Jacobson, A.E.; Boutaghou, M.N.; Alving, C.R.; Matyas, G.R. Characterization and optimization of heroin hapten-BSA conjugates: Method development for the synthesis of reproducible hapten-based vaccines. Anal. Bioanal. Chem. 2014, 406, 5927–5937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyas, G.R.; Muderhwa, J.M.; Alving, C.R. Oil-in-Water Liposomal Emulsions for Vaccine Delivery. Methods Enzymol. 2003, 373, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Matyas, G.R.; Rice, K.C.; Cheng, K.; Li, F.; Antoline, J.F.G.; Iyer, M.R.; Jacobson, A.E.; Mayorov, A.V.; Beck, Z.; Torres, O.B.; et al. Facial recognition of heroin vaccine opiates: Type 1 cross-reactivities of antibodies induced by hydrolytically stable haptenic surrogates of heroin, 6-acetylmorphine, and morphine. Vaccine 2014, 32, 1473–1479. [Google Scholar] [CrossRef] [Green Version]
- Bannon, A.W.; Malmberg, A.B. Models of Nociception: Hot-Plate, Tail-Flick, and Formalin Tests in Rodents. Curr. Protoc. Neurosci. 2007, 41, 8.9.1–8.9.16. [Google Scholar] [CrossRef] [PubMed]
- Dagan, R.; Eskola, J.; Leclerc, C.; Leroy, O. Reduced Response to Multiple Vaccines Sharing Common Protein Epitopes That Are Administered Simultaneously to Infants. Infect. Immun. 1998, 66, 2093–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, C.C.; Tenbergen-Meekes, A.M.; Poolman, J.T.; Beurret, M.; Zegers, B.J.M.; Rijkers, G.T. Effect of Carrier Priming on Immunogenicity of Saccharide-Protein Conjugate Vaccines. Infect. Immun. 1991, 59, 3504–3510. [Google Scholar] [CrossRef] [Green Version]
- Domènech, V.E.S.; Panthel, K.; Meinel, K.M.; Winter, S.E.; Rüssmann, H. Pre-existing anti-Salmonella vector immunity prevents the development of protective antigen-specific CD8 T-cell frequencies against murine listeriosis. Microbes Infect. 2007, 9, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Rollier, C.S.; Reyes-Sandoval, A.; Cottingham, M.G.; Ewer, K.; Hill, A.V. Viral vectors as vaccine platforms: Deployment in sight. Curr. Opin. Immunol. 2011, 23, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Van, T.T.H.; Baird, F.J.; Coloe, P.J.; Smooker, P.M. Pre-existing immunity against vaccine vectors–friend or foe? Microbiology 2013, 159, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutze, M.P.; Leclerc, C.; Jolivet, M.; Audibert, F.; Chedid, L. Carrier-induced epitopic suppression, a major issue for future synthetic vaccines. J. Immunol. 1985, 135, 2319–2322. [Google Scholar] [PubMed]
- Ben-Yedidia, T.; Arnon, R. Effect of pre-existing carrier immunity on the efficacy of synthetic influenza vaccine. Immunol. Lett. 1998, 64, 9–15. [Google Scholar] [CrossRef]
- Sulima, A.; Jalah, R.; Antoline, J.F.G.; Torres, O.B.; Imler, G.H.; Deschamps, J.R.; Beck, Z.; Alving, C.R.; Jacobson, A.E.; Rice, K.C.; et al. A Stable Heroin Analogue That Can Serve as a Vaccine Hapten to Induce Antibodies That Block the Effects of Heroin and Its Metabolites in Rodents and That Cross-React Immunologically with Related Drugs of Abuse. J. Med. Chem 2018, 61, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Schutze, M.P.; Deriaud, E.; Przewlocki, G.; LeClerc, C. Carrier-induced epitopic suppression is initiated through clonal dominance. J. Immunol. 1989, 142, 2635–2640. [Google Scholar]
- McCluskie, M.J.; Evans, D.M.; Zhang, N.; Benoit, M.; McElhiney, S.P.; Unnithan, M.; Demarco, S.C.; Clay, B.; Huber, C.; Deora, A.; et al. The effect of preexisting anti-carrier immunity on subsequent responses to CRM 197 or Qb-VLP conjugate vaccines. Immunopharmacol. Immunotoxicol. 2016, 38, 184–196. [Google Scholar] [CrossRef]
- Pecetta, S.; Surdo, P.L.; Tontini, M.; Proietti, D.; Zambonelli, C.; Bottomley, M.; Biagini, M.; Berti, F.; Costantino, P.; Romano, M. Carrier priming with CRM197 or diphtheria toxoid has a different impact on the immunogenicity of the respective glycoconjugates: Biophysical and immunochemical interpretation. Vaccine 2015, 33, 314–320. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, W.; Wang, Y.; Hu, T. Moderate PEGylation of the carrier protein improves the polysaccharide-specific immunogenicity of meningococcal group A polysaccharide conjugate vaccine. Vaccine 2015, 33, 3208–3214. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komla, E.; Torres, O.B.; Jalah, R.; Sulima, A.; Beck, Z.; Alving, C.R.; Jacobson, A.E.; Rice, K.C.; Matyas, G.R. Effect of Preexisting Immunity to Tetanus Toxoid on the Efficacy of Tetanus Toxoid-Conjugated Heroin Vaccine in Mice. Vaccines 2021, 9, 573. https://doi.org/10.3390/vaccines9060573
Komla E, Torres OB, Jalah R, Sulima A, Beck Z, Alving CR, Jacobson AE, Rice KC, Matyas GR. Effect of Preexisting Immunity to Tetanus Toxoid on the Efficacy of Tetanus Toxoid-Conjugated Heroin Vaccine in Mice. Vaccines. 2021; 9(6):573. https://doi.org/10.3390/vaccines9060573
Chicago/Turabian StyleKomla, Essie, Oscar B. Torres, Rashmi Jalah, Agnieszka Sulima, Zoltan Beck, Carl R. Alving, Arthur E. Jacobson, Kenner C. Rice, and Gary R. Matyas. 2021. "Effect of Preexisting Immunity to Tetanus Toxoid on the Efficacy of Tetanus Toxoid-Conjugated Heroin Vaccine in Mice" Vaccines 9, no. 6: 573. https://doi.org/10.3390/vaccines9060573