
Local Sustained GM-CSF Delivery by Genetically Engineered Encapsulated Cells Enhanced Both Cellular and Humoral SARS-CoV-2 Spike-Specific Immune Response in an Experimental Murine Spike DNA Vaccination Model

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Plasmid Amplification, Purification, and Validation
2.3. Plasmid Injection and Electroporation
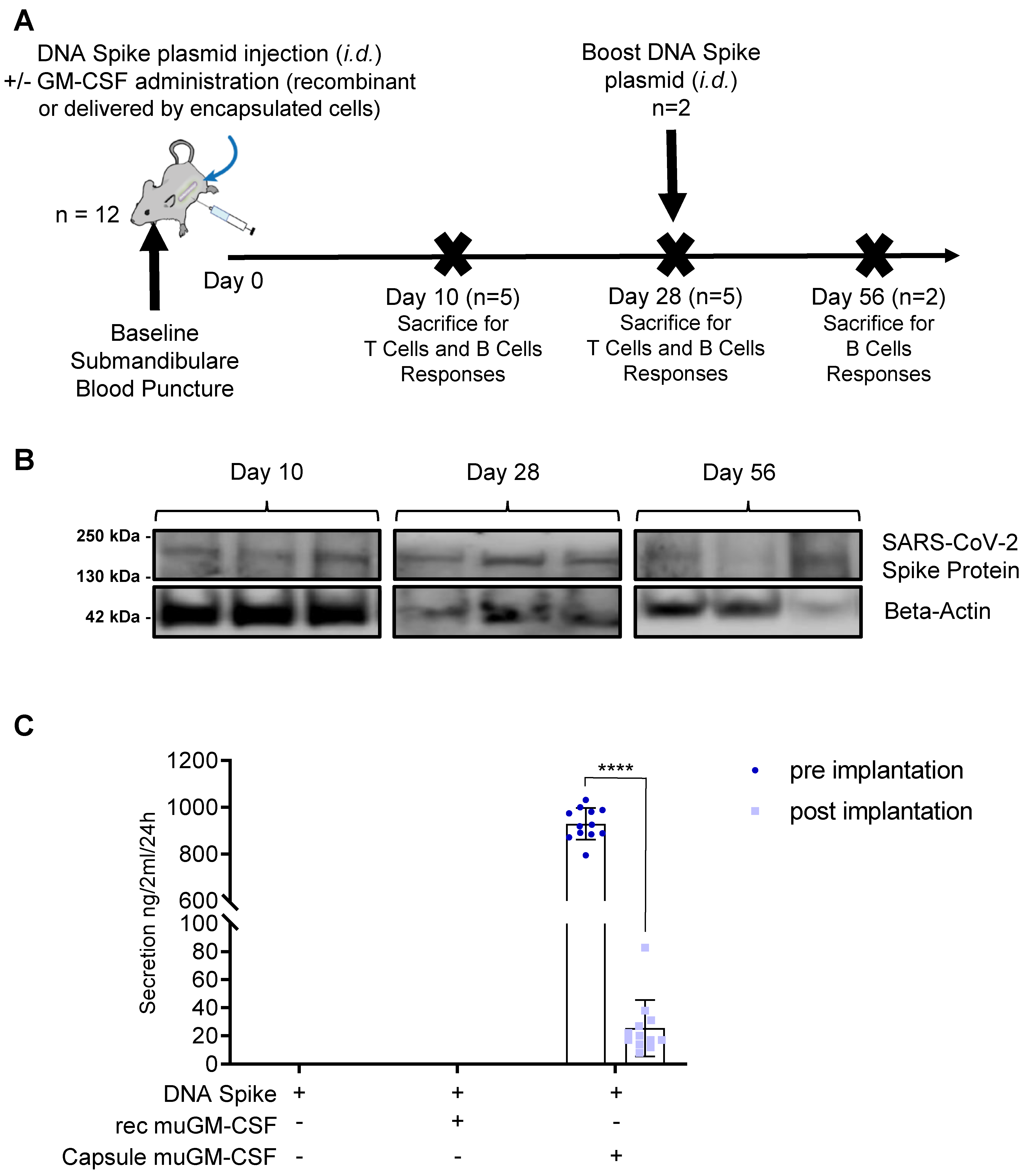
2.4. Dermal SARS-CoV-2 Spike Protein Expression by Western Blot Analysis
2.5. muGM-CSF Adjuvant Administration
2.6. muGM-CSF Adjuvant Quantification by ELISA
2.7. Immunization Scheme
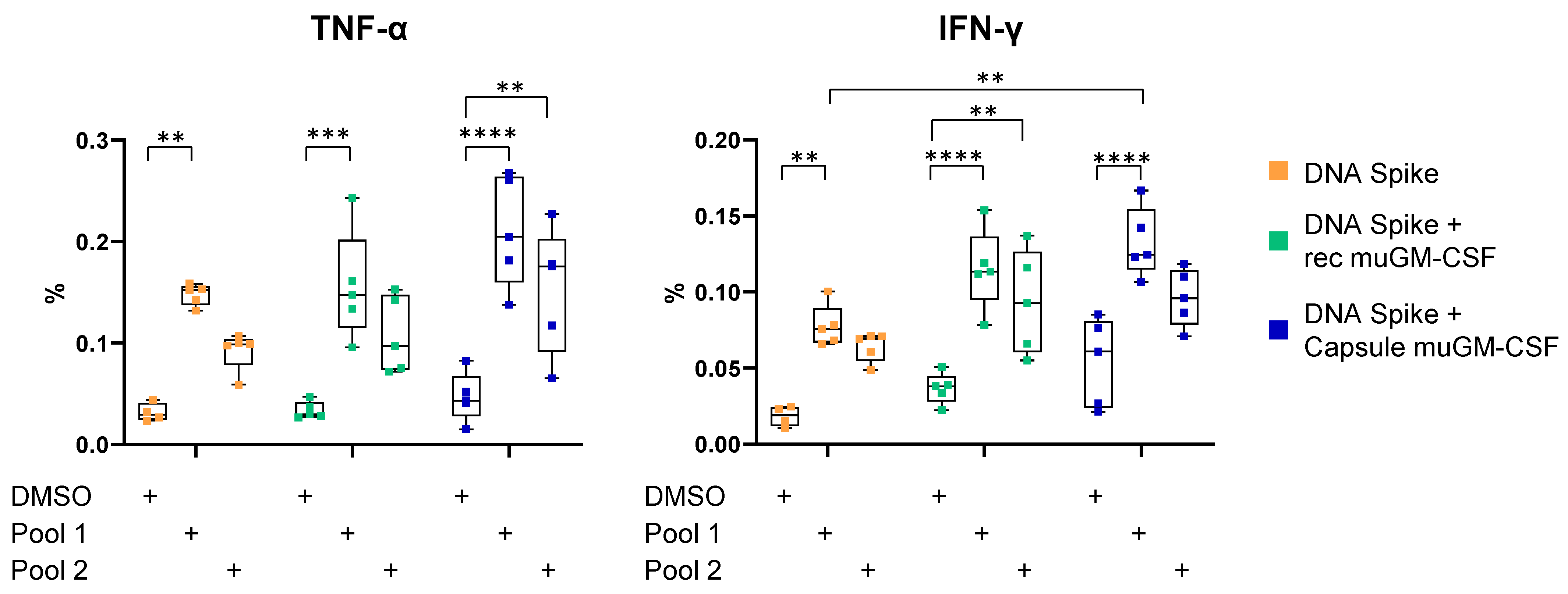
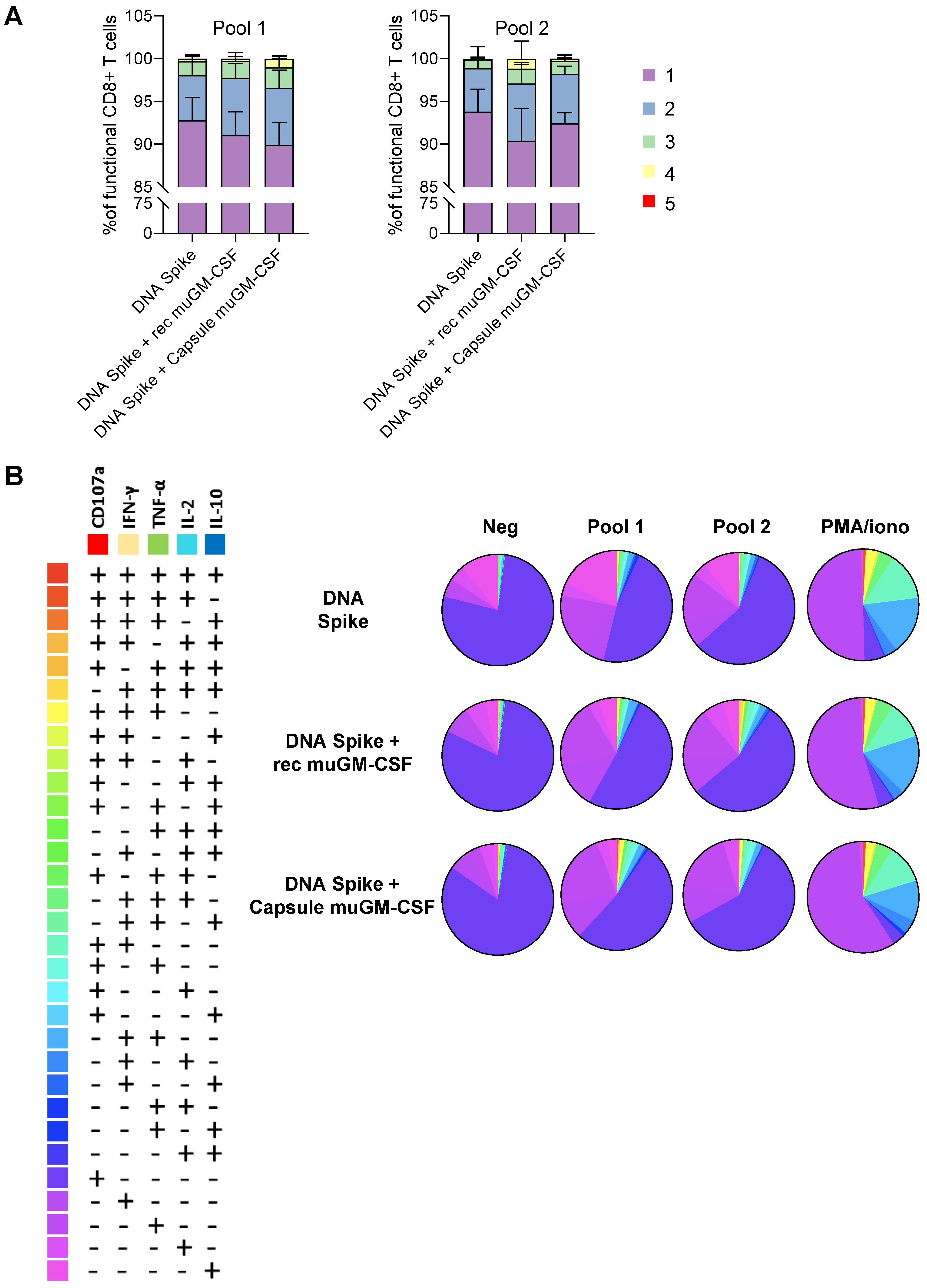
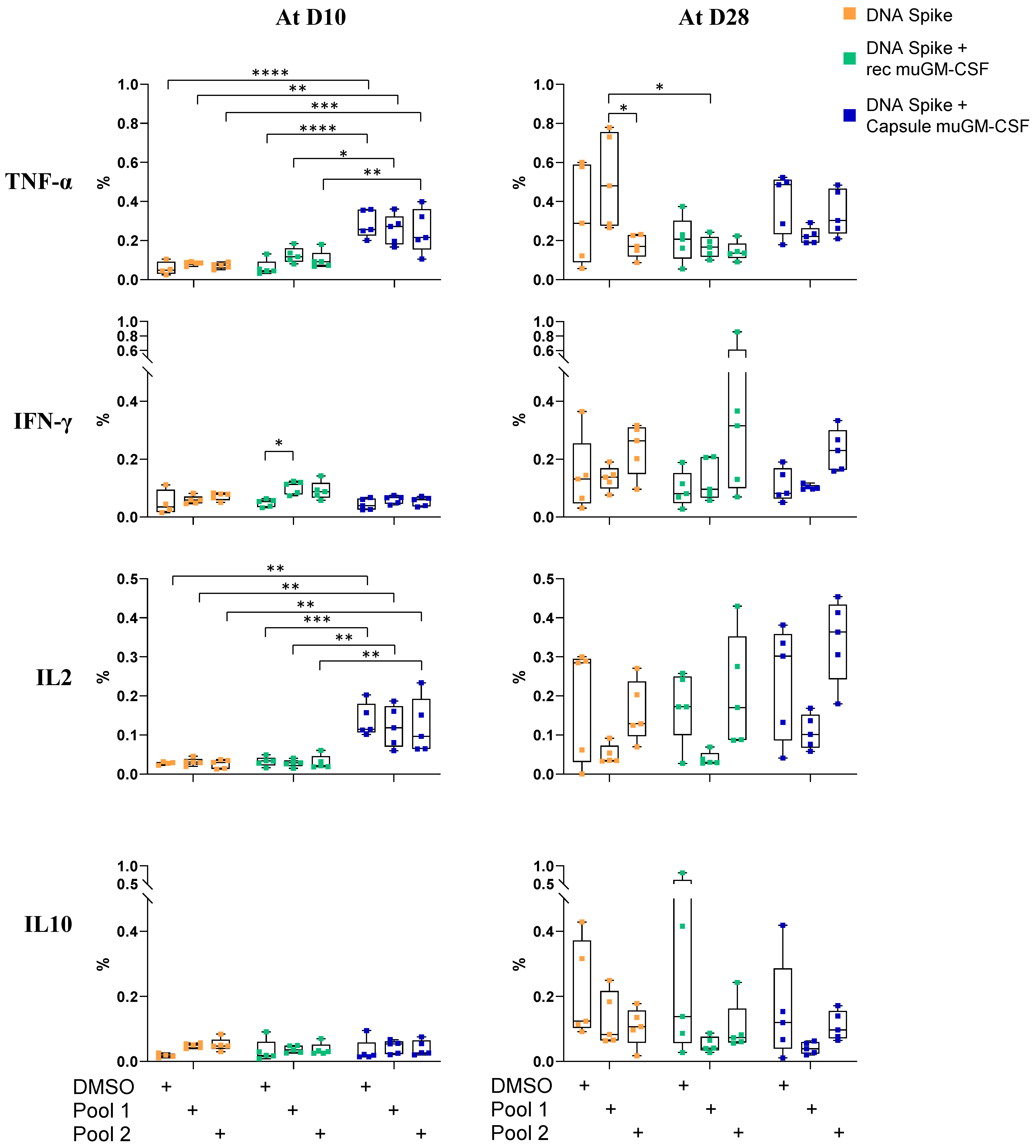
2.8. T Cell Characterization by Intracellular Cytokine Staining (ICS) and Flow Cytometry Analysis
2.9. Quantification of SARS-CoV-2 Spike Protein Antibodies by ELISA
2.10. SARS-CoV-2 Surrogate Virus Neutralization Test (sVNT)
2.11. Statistical Analysis
3. Results
3.1. Validation of the Vaccination Strategy
3.2. Cellular-Mediated Immune Responses to SARS-CoV-2 Spike Protein
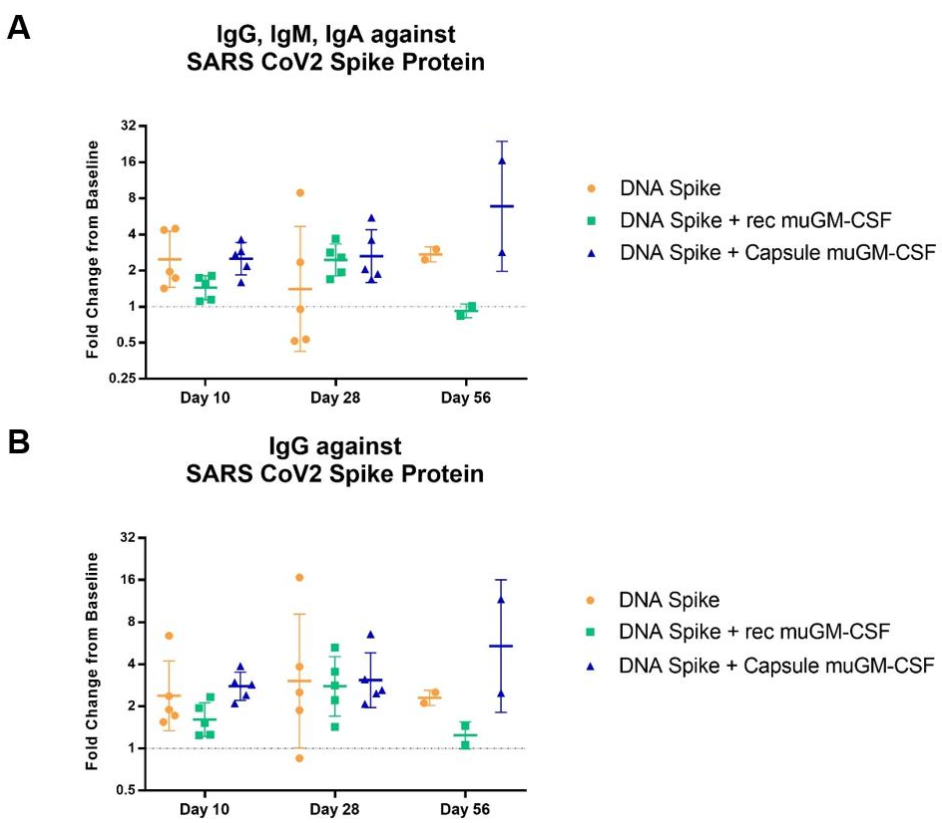
3.3. Humoral-Mediated Immune Responses to SARS-CoV-2 Spike Protein
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (Previously 2019-nCoV) Infection by a Highly Potent Pan-Coronavirus Fusion Inhibitor Targeting Its Spike Protein That Harbors a High Capacity to Mediate Membrane Fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, H.; Bouslama, Z.; Brandenburg, J.-T.; da Rocha, J.; Hamdi, Y.; Ghedira, K.; Srairi-Abid, N.; Hazelhurst, S. Interaction of the Spike Protein RBD from SARS-CoV-2 with ACE2: Similarity with SARS-CoV, Hot-Spot Analysis and Effect of the Receptor Polymorphism. Biochem. Biophys. Res. Commun. 2020, 527, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, U.J.; Bukreyev, A.; Yang, L.; Lamirande, E.W.; Murphy, B.R.; Subbarao, K.; Collins, P.L. Contributions of the Structural Proteins of Severe Acute Respiratory Syndrome Coronavirus to Protective Immunity. Proc. Natl. Acad. Sci. USA 2004, 101, 9804–9809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Hillyer, C.; Du, L. Neutralizing Antibodies against SARS-CoV-2 and Other Human Coronaviruses. Trends Immunol. 2020, 41, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Mai, J.; Zhou, W.; Yu, W.; Zhan, Y.; Wang, N.; Epstein, N.; Yang, Y. Immunoinformatic Analysis of T- and B-Cell Epitopes for SARS-CoV-2 Vaccine Design. Vaccines 2020, 8, 355. [Google Scholar] [CrossRef] [PubMed]
- Mubarak, A.; Alturaiki, W.; Hemida, M.G. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Infection, Immunological Response, and Vaccine Development. J. Immunol. Res. 2019, 2019, e6491738. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gao, X. Immunological Responses against SARS-Coronavirus Infection in Humans. Cell. Mol. Immunol. 2004, 1, 119–122. [Google Scholar]
- McConkey, S.J.; Reece, W.H.H.; Moorthy, V.S.; Webster, D.; Dunachie, S.; Butcher, G.; Vuola, J.M.; Blanchard, T.J.; Gothard, P.; Watkins, K.; et al. Enhanced T-cell Immunogenicity of Plasmid DNA Vaccines Boosted by Recombinant Modified Vaccinia Virus Ankara in Humans. Nat. Med. 2003, 9, 729–735. [Google Scholar] [CrossRef]
- Wang, R.; Doolan, D.L.; Le, T.P.; Hedstrom, R.C.; Coonan, K.M.; Charoenvit, Y.; Jones, T.R.; Hobart, P.; Margalith, M.; Ng, J.; et al. Induction of Antigen-Specific Cytotoxic T Lymphocytes in Humans by a Malaria DNA Vaccine. Science 1998, 282, 476–480. [Google Scholar] [CrossRef]
- Kutzler, M.A.; Weiner, D.B. DNA Vaccines: Ready for Prime Time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Lu, S.; Wang, S.; Grimes-Serrano, J.M. Current Progress of DNA Vaccine Studies in Humans. Expert Rev. Vaccines 2008, 7, 175–191. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, W.; Huang, Y.; Roberts, A.; Murphy, B.R.; Subbarao, K.; Nabel, G.J. A DNA Vaccine Induces SARS Coronavirus Neutralization and Protective Immunity in Mice. Nature 2004, 428, 561–564. [Google Scholar] [CrossRef] [Green Version]
- Sajadian, A.; Tabarraei, A.; Soleimanjahi, H.; Fotouhi, F.; Gorji, A.; Ghaemi, A. Comparing the Effect of Toll-Like Receptor Agonist Adjuvants on the Efficiency of a DNA Vaccine. Arch. Virol. 2014, 159, 1951–1960. [Google Scholar] [CrossRef] [Green Version]
- Kwissa, M.; Amara, R.R.; Robinson, H.L.; Moss, B.; Alkan, S.; Jabbar, A.; Villinger, F.; Pulendran, B. Adjuvanting a DNA Vaccine with a TLR9 Ligand Plus Flt3 Ligand Results in Enhanced Cellular Immunity against the Simian Immunodeficiency Virus. J. Exp. Med. 2007, 204, 2733–2746. [Google Scholar] [CrossRef] [Green Version]
- Barouch, D.H.; Santra, S.; Tenner-Racz, K.; Racz, P.; Kuroda, M.J.; Schmitz, J.E.; Jackson, S.S.; Lifton, M.A.; Freed, D.C.; Perry, H.C.; et al. Potent CD4+T Cell Responses Elicited by a Bicistronic HIV-1 DNA Vaccine Expressing gp120 and GM-CSF. J. Immunol. 2002, 168, 562–568. [Google Scholar] [CrossRef] [Green Version]
- Okada, E.; Sasaki, S.; Ishii, N.; Aoki, I.; Yasuda, T.; Nishioka, K.; Fukushima, J.; Miyazaki, J.; Wahren, B.; Okuda, K. Intranasal Immunization of a DNA Vaccine with IL-12- and Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF)-Expressing Plasmids in Liposomes Induces Strong Mucosal and Cell-Mediated Immune Responses against HIV-1 Antigens. J. Immunol. 1997, 159, 3638–3647. [Google Scholar]
- Weiss, W.R.; Ishii, K.J.; Hedstrom, R.C.; Sedegah, M.; Ichino, M.; Barnhart, K.; Klinman, D.M.; Hoffman, S.L. A Plasmid Encoding Murine Granulocyte-Macrophage Colony-Stimulating Factor Increases Protection Conferred by a Malaria DNA Vaccine. J. Immunol. 1998, 161, 2325–2332. [Google Scholar]
- Bråve, A.; Johansson, U.; Hallengärd, D.; Heidari, S.; Gullberg, H.; Wahren, B.; Hinkula, J.; Spetz, A.-L. Induction of HIV-1-Specific Cellular and Humoral Immune Responses Following Immunization with HIV-DNA Adjuvanted with Activated Apoptotic Lymphocytes. Vaccine 2010, 28, 2080–2087. [Google Scholar] [CrossRef]
- Sardesai, N.Y.; Weiner, D.B. Electroporation Delivery of DNA Vaccines: Prospects for Success. Curr. Opin. Immunol. 2011, 23, 421–429. [Google Scholar] [CrossRef] [Green Version]
- DNA Vaccination in the Skin Using Microneedles Improves Protection against Influenza n.d. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3392990/ (accessed on 26 November 2020).
- Heller, R.; Cruz, Y.; Heller, L.C.; Gilbert, R.A.; Jaroszeski, M.J. Electrically Mediated Delivery of Plasmid DNA to the Skin, Using a Multielectrode Array. Hum. Gene Ther. 2010, 21, 357–362. [Google Scholar] [CrossRef]
- Gong, J.-Y.; Liu, X.; Dong, Y.; Zhou, T.-H.; Li, J.-W. Construction and Expression of a Recombinant Eukaryotic Expression Plasmid Containing the preS1-preS2-S Genes of Hepatitis B Virus and the Granulocyte-Macrophage Colony Stimulating Factor Gene: A Study of Its Immunomodulatory Effects. Biomed. Rep. 2013, 1, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Encke, J.; Bernardin, J.; Geib, J.; Barbakadze, G.; Bujdoso, R.; Stremmel, W. Genetic Vaccination with Flt3-L and GM-CSF as Adjuvants: Enhancement of Cellular and Humoral Immune Responses That Results in Protective Immunity in a Murine Model of Hepatitis C Virus Infection. World J. Gastroenterol. 2006, 12, 7118–7125. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-W.; Chang, J.-L.; Tsao, Y.-P.; Huang, C.-W.; Kuo, S.-W.; Chen, S.-L. Co-Vaccination with Adeno-Associated Virus Vectors Encoding Human Papillomavirus 16 L1 Proteins and Adenovirus Encoding Murine GM-CSF Can Elicit Strong and Prolonged Neutralizing Antibody. Int. J. Cancer 2005, 113, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, L.; Zhou, S.; Wang, Z.; Ruan, J.; Tang, L.; Jia, Z.; Cui, M.; Zhao, L.; Fu, Z.F. Recombinant Rabies Virus Expressing Dog GM-CSF Is an Efficacious Oral Rabies Vaccine for Dogs. Oncotarget 2015, 6, 38504–38516. [Google Scholar] [CrossRef] [Green Version]
- Suschak, J.J.; Bagley, K.; Shoemaker, C.J.; Six, C.; Kwilas, S.; Dupuy, L.C.; Schmaljohn, C.S. The Genetic Adjuvants Interleukin-12 and Granulocyte-Macrophage Colony Stimulating Factor Enhance the Immunogenicity of an Ebola Virus Deoxyribonucleic Acid Vaccine in Mice. J. Infect. Dis. 2018, 218, S519–S527. [Google Scholar] [CrossRef]
- Manara, C.; Brazzoli, M.; Piccioli, D.; Taccone, M.; D’Oro, U.; Maione, D.; Frigimelica, E. Co-Administration of GM-CSF Expressing RNA Is a Powerful Tool to Enhance Potency of SAM-Based Vaccines. Vaccine 2019, 37, 4204–4213. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Jia, Y.; Wang, C.; Han, Q.; Lu, Z.H.; Yang, Z. Adenoviral-Expressed Recombinant Granulocyte Monocyte Colony-Stimulating Factor (GM-CSF) Enhances Protective Immunity Induced by Inactivated Newcastle Disease Virus (NDV) Vaccine. Antivir. Res. 2017, 144, 322–329. [Google Scholar] [CrossRef]
- Immunogenicity Evaluation of Modified Adenovirus Vaccines Expressing Porcine Circovirus Type 2 Capsid Protein in Pigs—Abstract—Europe PMC n.d. Available online: https://europepmc.org/article/med/27996636 (accessed on 3 December 2020).
- Santana, V.C.; Almeida, R.R.; Ribeiro, S.P.; Ferreira, L.C.D.S.; Kalil, J.; Rosa, D.S.; Neto, E.C. Co-Administration of Plasmid-Encoded Granulocyte-Macrophage Colony-Stimulating Factor Increases Human Immunodeficiency Virus-1 DNA Vaccine-induced Polyfunctional CD4+ T-Cell Responses. Mem. Inst. Oswaldo Cruz 2015, 110, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Reali, E.; Canter, D.; Zeytin, H.; Schlom, J.; Greiner, J. Comparative Studies of Avipox-GM-CSF versus Recombinant GM-CSF Protein as Immune Adjuvants with Different Vaccine Platforms. Vaccine 2005, 23, 2909–2921. [Google Scholar] [CrossRef]
- Serafini, P.; Carbley, R.; Noonan, K.A.; Tan, G.; Bronte, V.; Borrello, I. High-Dose Granulocyte-Macrophage Colony-Stimulating Factor-Producing Vaccines Impair the Immune Response through the Recruitment of Myeloid Suppressor Cells. Cancer Res. 2004, 64, 6337–6343. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, P.; Budnick, I.; Singh, M.; Thiruppathi, M.; Alharshawi, K.; Elshabrawy, H.; Holterman, M.J.; Prabhakar, B.S. Dual Role of GM-CSF as a Pro-Inflammatory and a Regulatory Cytokine: Implications for Immune Therapy. J. Interf. Cytokine Res. 2015, 35, 585–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmiani, G.; Castelli, C.; Pilla, L.; Santinami, M.; Colombo, M.; Rivoltini, L. Opposite Immune Functions of GM-CSF Administered as Vaccine Adjuvant in Cancer Patients. Ann. Oncol. 2007, 18, 226–232. [Google Scholar] [CrossRef]
- Bowen, W.S.; Svrivastava, A.K.; Batra, L.; Barsoumian, H.; Shirwan, H. Current Challenges for Cancer Vaccine Adjuvant Development. Expert Rev. Vaccines 2018, 17, 207–215. [Google Scholar] [CrossRef]
- Gouttefangeas, C.; Rammensee, H.-G. Personalized Cancer Vaccines: Adjuvants Are Important, Too. Cancer Immunol. Immunother. 2018, 67, 1911–1918. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, P.K.; Srinivasan, R.; Perumal, N. Selection of Animal Models for COVID-19 Research. VirusDisease 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A Mouse-Adapted Model of SARS-CoV-2 to Test COVID-19 Countermeasures. Nature 2020, 586, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The Pathogenicity of SARS-CoV-2 in hACE2 Transgenic Mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vernet, R.; Charrier, E.; Cosset, E.; Fièvre, S.; Tomasello, U.; Grogg, J.; Mach, N. Local Sustained GM-CSF Delivery by Genetically Engineered Encapsulated Cells Enhanced Both Cellular and Humoral SARS-CoV-2 Spike-Specific Immune Response in an Experimental Murine Spike DNA Vaccination Model. Vaccines 2021, 9, 484. https://doi.org/10.3390/vaccines9050484
Vernet R, Charrier E, Cosset E, Fièvre S, Tomasello U, Grogg J, Mach N. Local Sustained GM-CSF Delivery by Genetically Engineered Encapsulated Cells Enhanced Both Cellular and Humoral SARS-CoV-2 Spike-Specific Immune Response in an Experimental Murine Spike DNA Vaccination Model. Vaccines. 2021; 9(5):484. https://doi.org/10.3390/vaccines9050484
Chicago/Turabian StyleVernet, Rémi, Emily Charrier, Erika Cosset, Sabine Fièvre, Ugo Tomasello, Julien Grogg, and Nicolas Mach. 2021. "Local Sustained GM-CSF Delivery by Genetically Engineered Encapsulated Cells Enhanced Both Cellular and Humoral SARS-CoV-2 Spike-Specific Immune Response in an Experimental Murine Spike DNA Vaccination Model" Vaccines 9, no. 5: 484. https://doi.org/10.3390/vaccines9050484