
Synthesis of Escherichia coli OmpA Oral Nanoparticles and Evaluation of Immune Functions against the Major Etiologic Agent of Cow Mastitis

Abstract: Background
1. Introduction
2. Materials and Methods
2.1. Animals and Bacterial Strains
2.2. Expression, Purification, and Preparation of Nanoparticles of OmpA
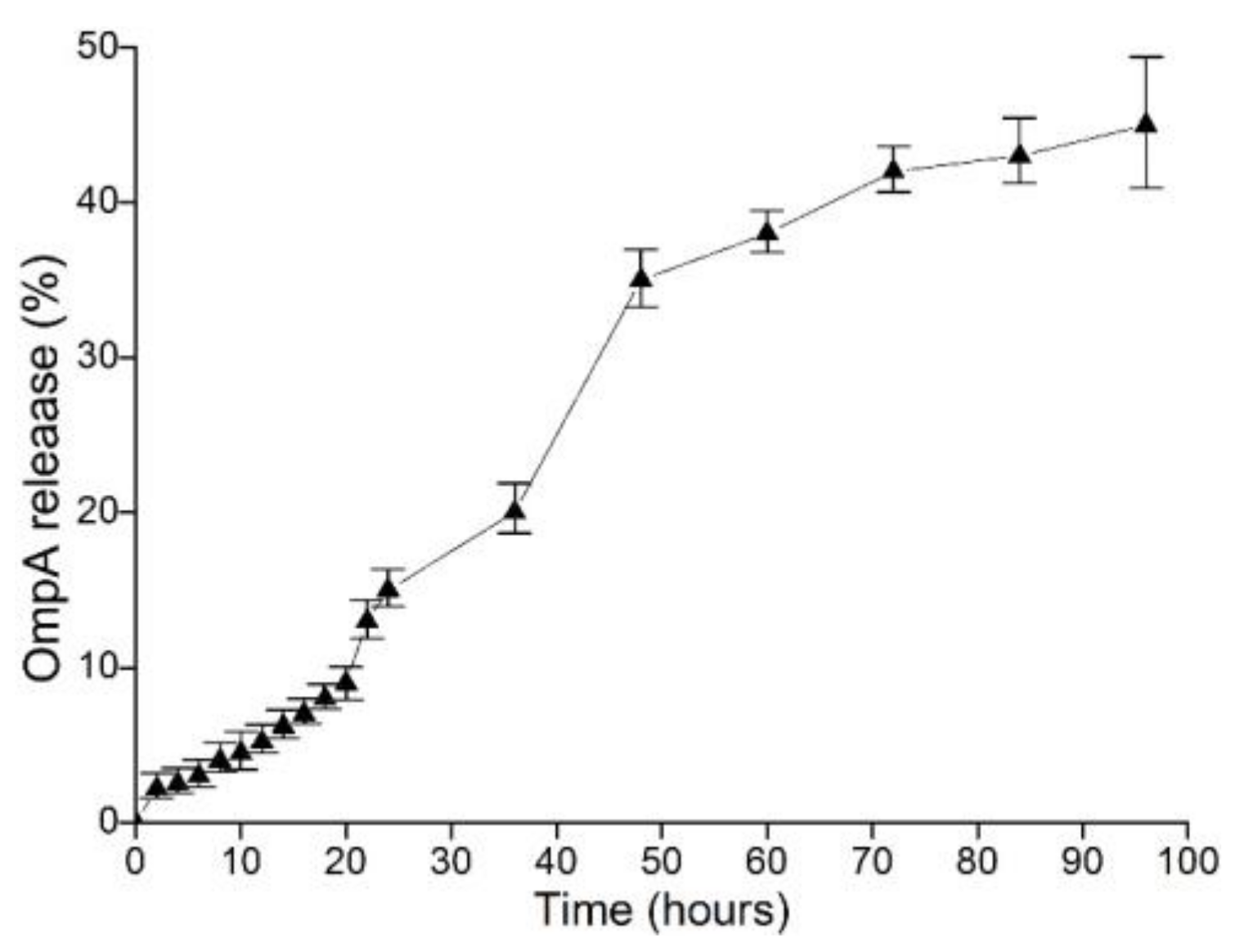
2.3. In Vitro Release of NP-OmpA
2.4. The Optimal Preparation Conditions for NP-OmpA
2.5. Immunoprotective Effect of NP-OmpA
2.6. Organ Index, White Blood Cell (WBC) Count, and Leukocyte Phagocytosis
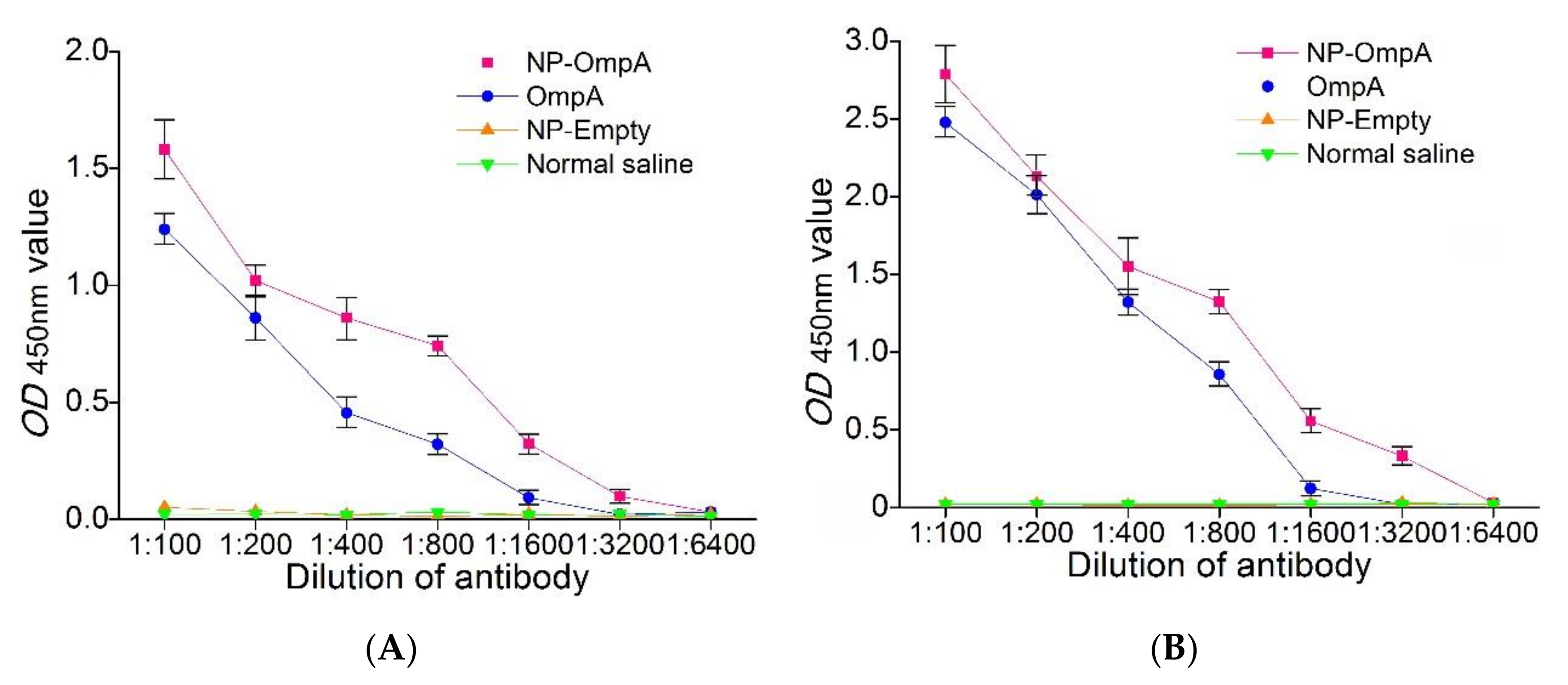
2.7. Detection of the Interaction between Antiserum and E. coli, and Antiserum Titer
2.8. Biochemical Indexes for Physiological Function of Visceral Organs
2.9. Determination of Expression of Immune-Related Genes and Inflammation-Related Genes by qRT-PCR
2.10. Histopathological Morphology of Injury to Visceral Organs
2.11. Statistical Analysis
3. Results
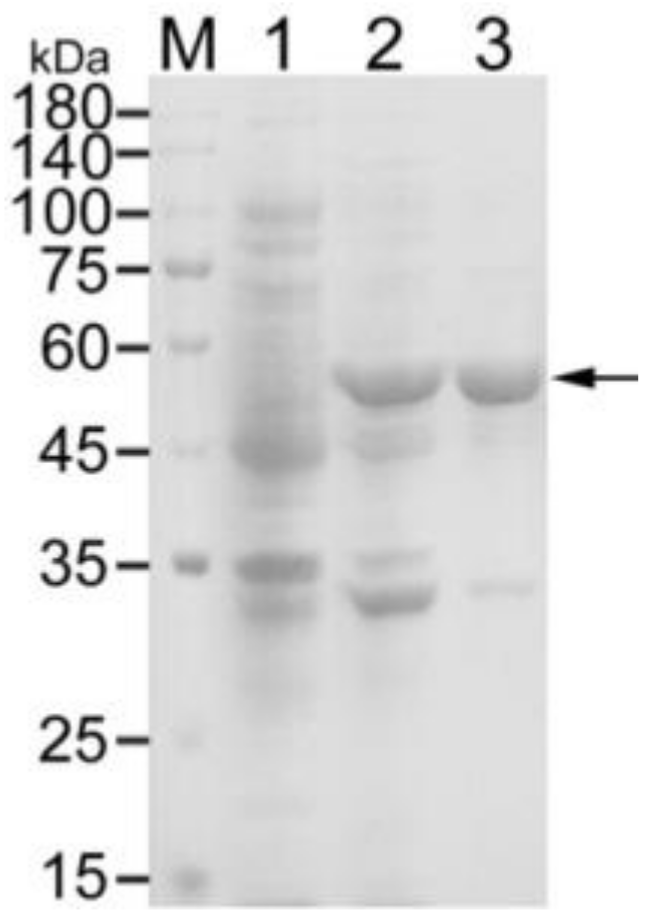
3.1. Expression and Purification of Recombinant OmpA
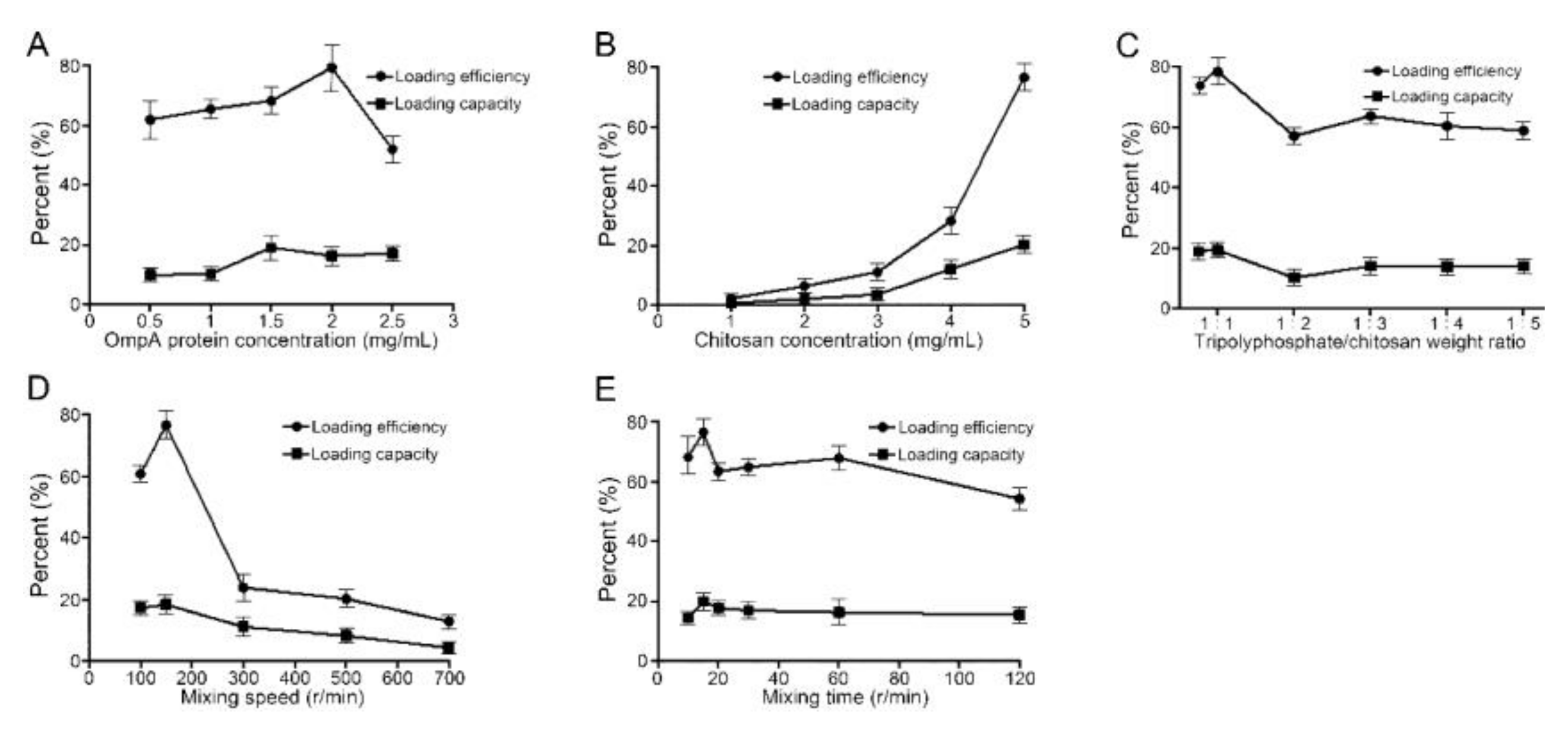
3.2. Optimal Preparation Conditions for NP-OmpA
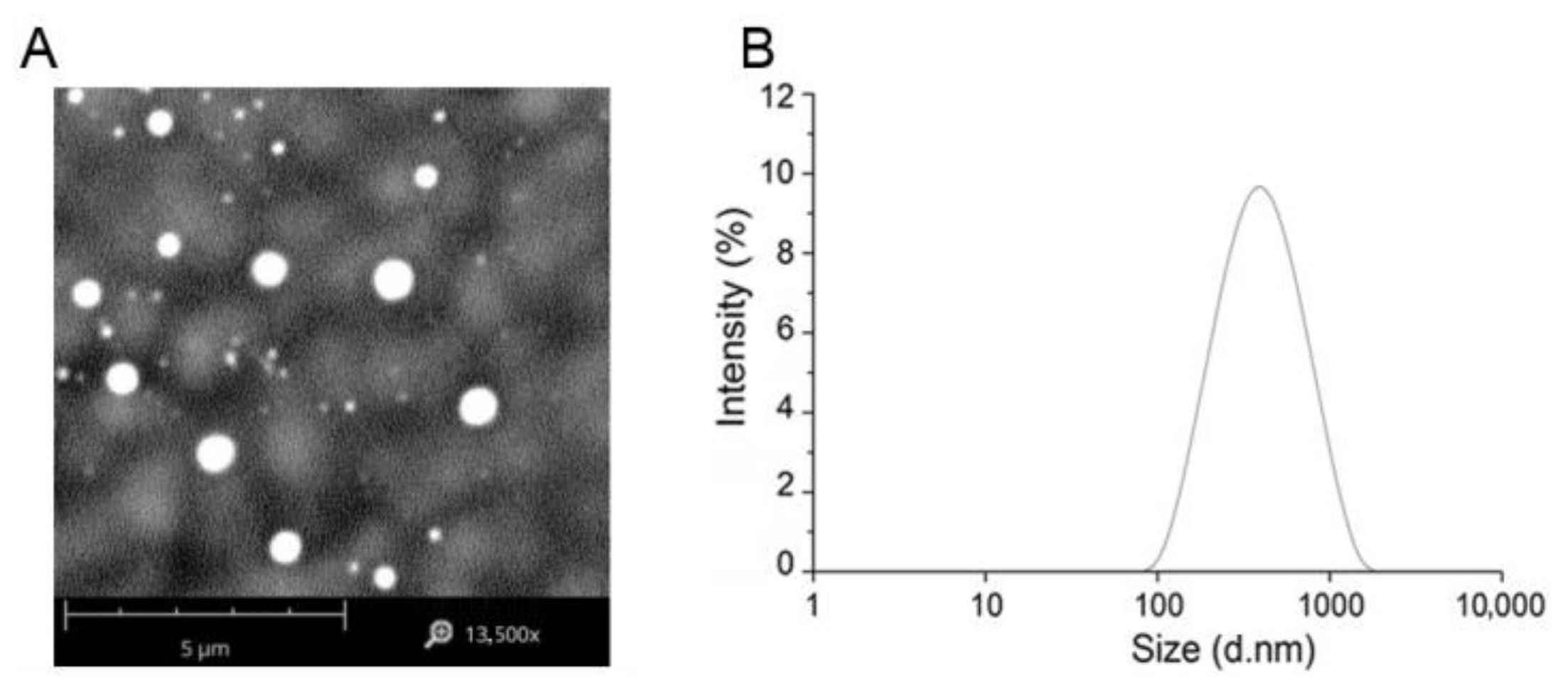
3.3. Preparation of OmpA Nanoparticles
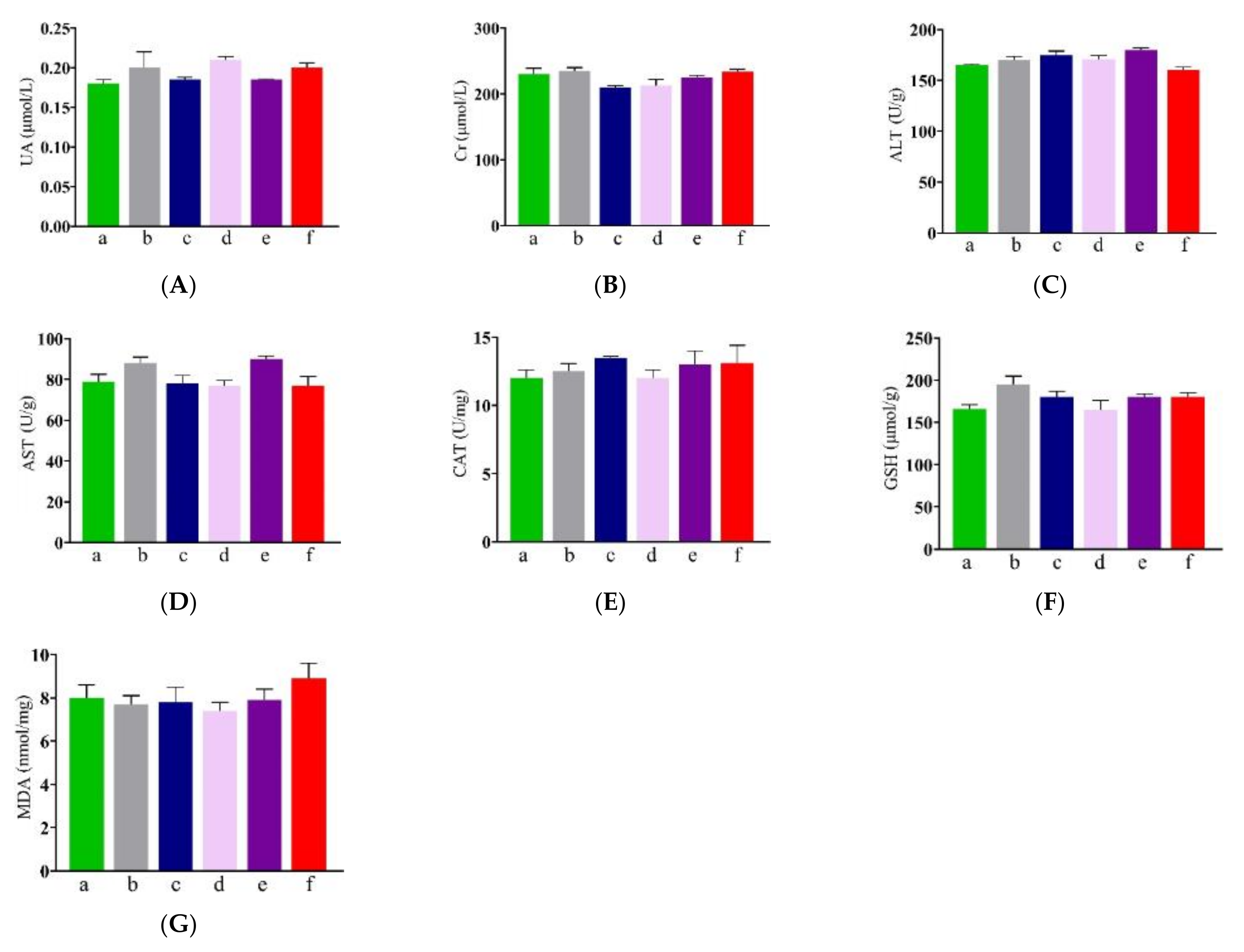
3.4. Effect of NP-OmpA on Mouse Liver and Kidney Functions
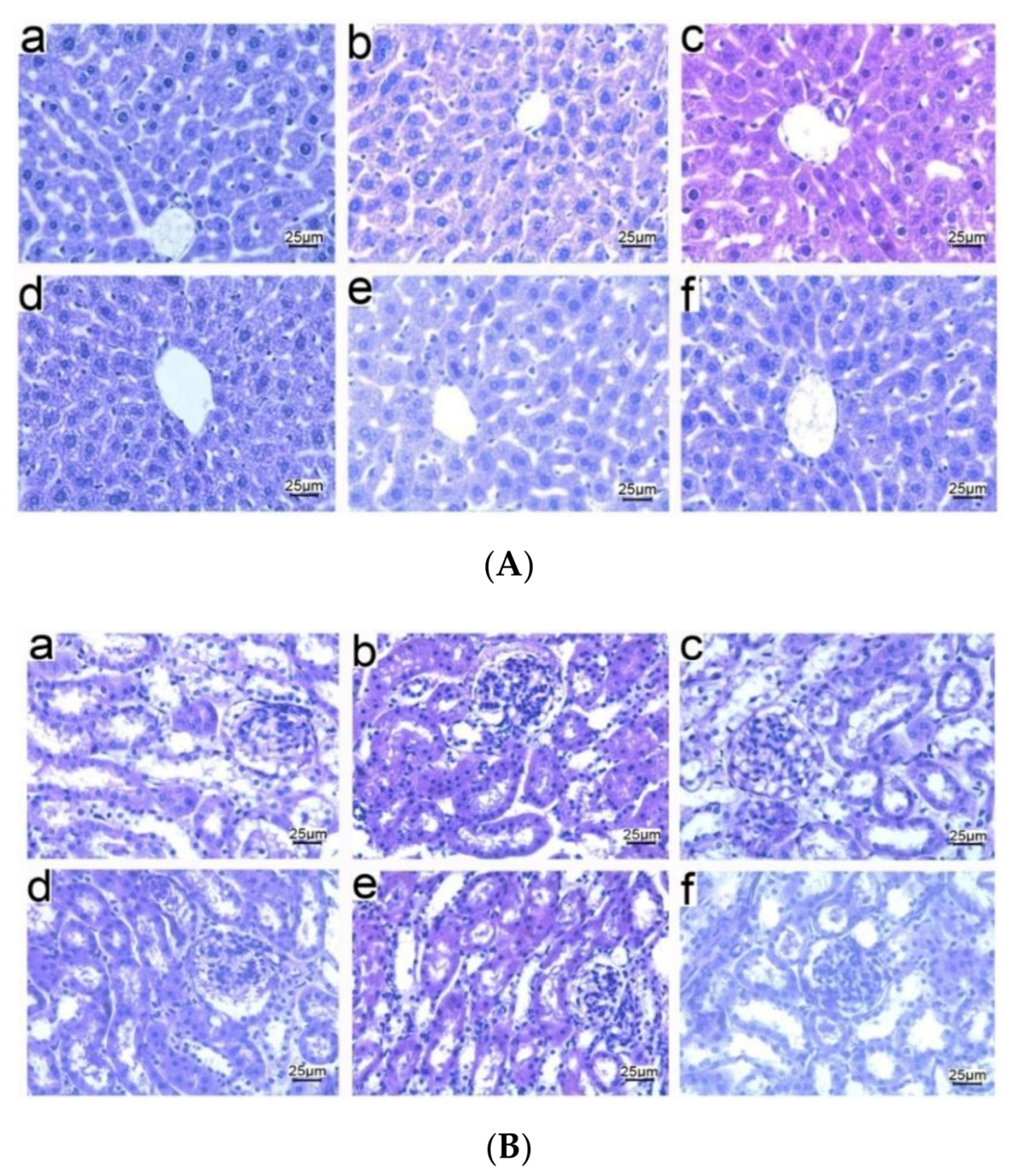
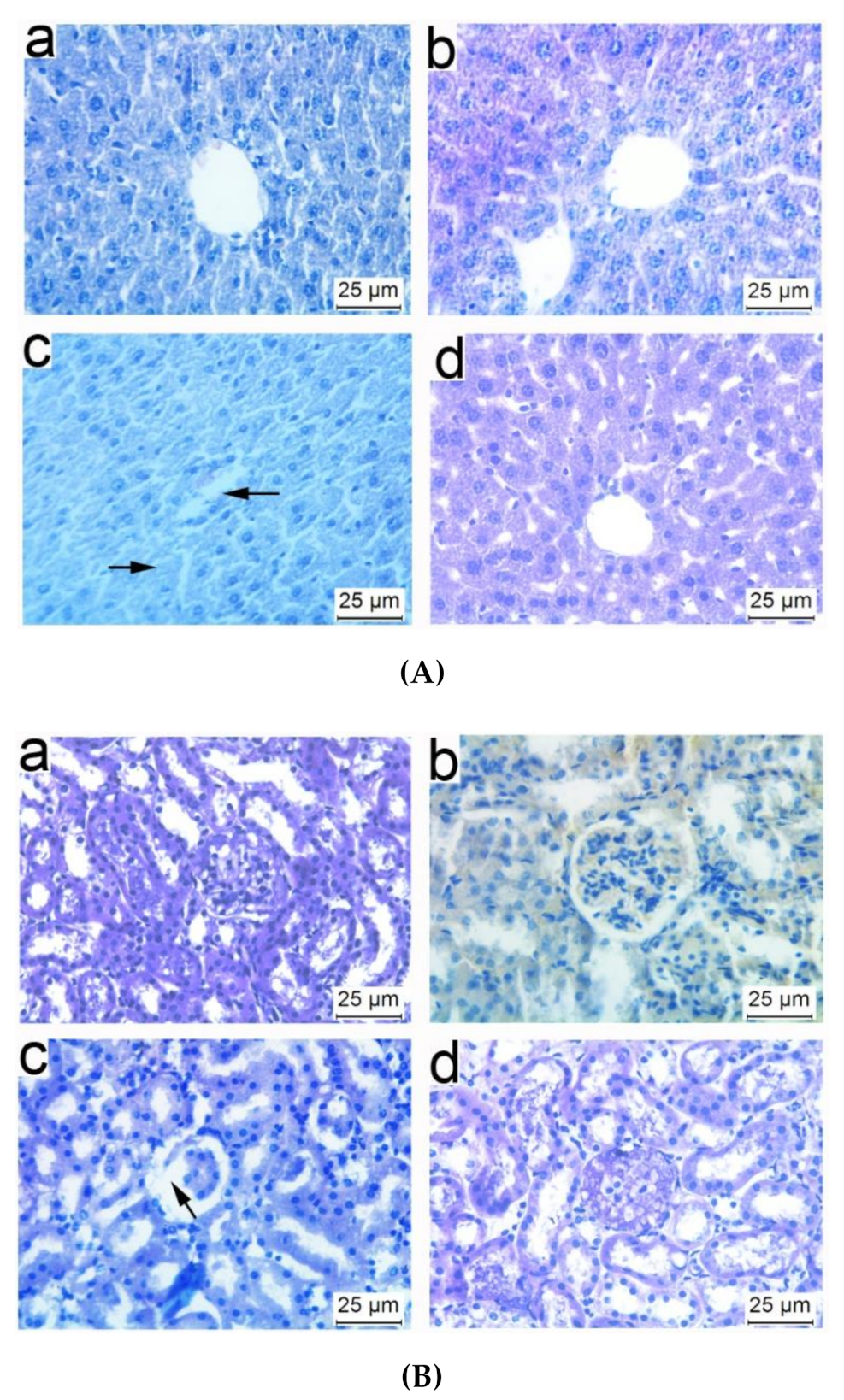
3.5. Histopathological Observations of Tissues from Mice Immunized with NP-OmpA
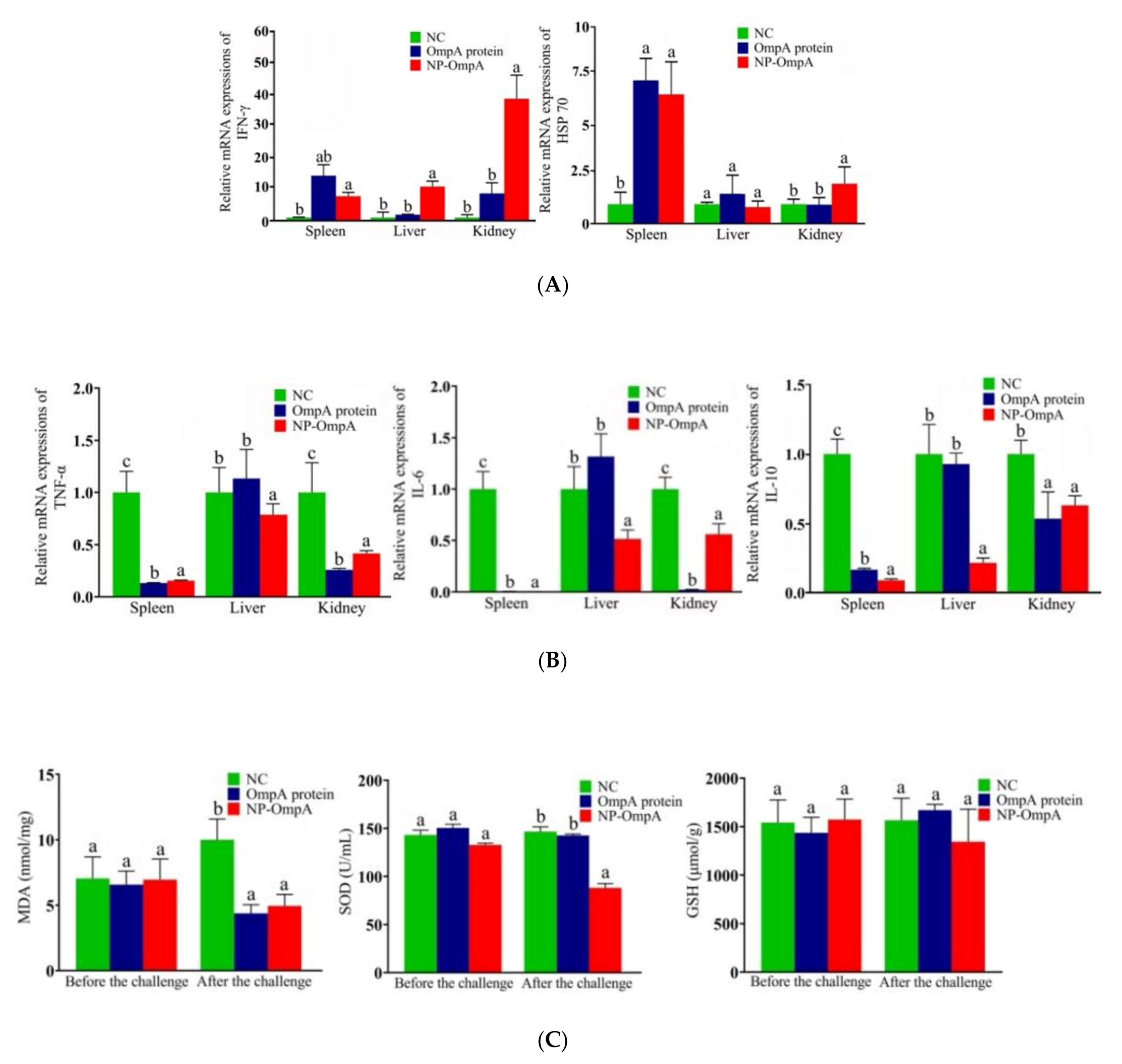
3.6. Immuno-Stimulating Activity of NP-OmpA
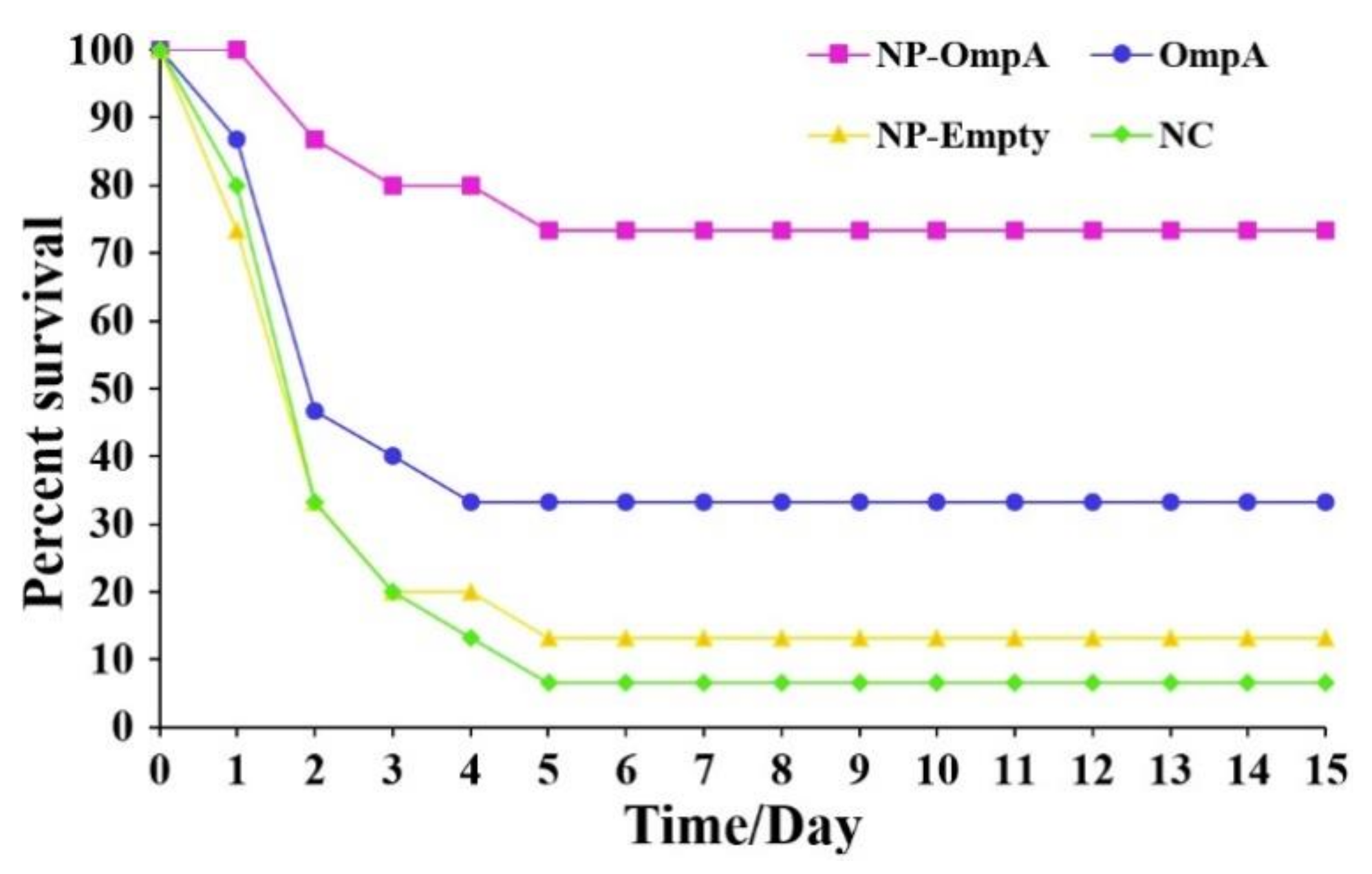
3.7. Immunoprotective Effect of NP-OmpA
3.8. Liver and Kidney Histopathology of Mice Challenged with E. coli
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Askari, N.; Ghanbarpour, R. Molecular investigation of the colicinogenic Escherichia coli strains that are capable of inhibiting E. coli O157: H7 In Vitro. BMC Vet. Res. 2019, 1, 14. [Google Scholar] [CrossRef]
- Fazel, F.; Jamshidi, A.; Khoramian, B. Phenotypic and genotypic study on antimicrobial resistance patterns of E. coli isolates from bovine mastitis. Microb. Pathog. 2019, 132, 355–361. [Google Scholar] [CrossRef]
- Lavon, Y.; Leitner, G.; Kressel, Y.; Ezra, E.; Wolfenson, D. Comparing effects of bovine Streptococcus and Escherichia coli mastitis on impaired reproductive performance. J. Dairy Sci. 2019, 11, 10587–10598. [Google Scholar] [CrossRef] [PubMed]
- Belotserkovsky, I.; Sansonetti, P.J. Shigella and enteroinvasive Escherichia coli. Curr. Top. Microbiol. Immunol. 2018, 416, 1–26. [Google Scholar] [PubMed]
- Sheen, S.; Huang, C.Y.; Ramos, R.; Chien, S.Y.; Scullen, O.J.; Sommers, C. Lethality prediction for Escherichia coli O157: H7 and uropathogenic E. coli in ground chicken treated with high pressure processing and trans-cinnamaldehyde. J. Food Sci. 2018, 3, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M. E. coli and colorectal cancer: A complex relationship that deserves a critical mindset. Crit. Rev. Microbiol. 2018, 5, 619–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowley, C.A.; Anderson, C.J.; Kendall, M.M. Ethanolamine influences human commensal Escherichia coli growth, gene expression, and competition with enterohemorrhagic E. coli O157: H7. mBio 2018, 5, 1429-18. [Google Scholar]
- Zhao, W.D.; Liu, D.X.; Wei, J.Y.; Miao, Z.W.; Zhang, K.; Su, Z.K.; Zhang, X.W.; Li, Q.; Fanf, W.G.; Qin, X.X. Caspr1 is a host receptor for meningitis-causing Escherichia coli. Nat. Commun. 2018, 1, 2296. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.I.; Równicki, M.; Wojciechowska, M.; Trylska, J. Antimicrobial synergy between mRNA targeted peptide nucleic acid and antibiotics in E. coli. Bioorg. Med. Chem. Lett. 2018, 18, 3094–3098. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibi-otics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global over-view. Poult. Sci. 2019, 4, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Confer, A.W.; Ayalew, S. The OmpA family of proteins: Roles in bacterial pathogenesis and immunity. Vet. Microbiol. 2013, 3-4, 207–222. [Google Scholar] [CrossRef]
- Boags, A.T.; Samsudin, F.; Khalid, S. Binding from both sides: TolR and full-length OmpA bind and maintain the local structure of the E. coli cell wall. Structure 2019, 4, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, W.S.; Yang, Y.Y.; Lin, P.H.; Chang, C.C.; Wu, H.H. Recombinant OmpA protein fragments mediate interleukin-17 regulation to prevent Escherichia coli meningitis. J. Microbiol. Immunol. Infect. 2016, 6, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; Liao, Y.; Zhang, J.; Wang, Y.; Liu, Z.; Cheng, P.; Wang, X.Y.; Zou, Q.M.; Gu, J. Rational de-sign and evaluation of an artificial Escherichia coli K1 protein vaccine candidate based on the struc-ture of OmpA. Front. Cell Infect. Microbiol. 2018, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wu, N.N.; Rong, N.; Kang, C.; Chen, C.L.; Wu, S.Q. Immunoprotective evaluation of Escherichia coli outer membrane protein A against the main pathogens of animal mastitis. Trop. J. Pharm. Res. 2020, 1, 155–162. [Google Scholar] [CrossRef]
- Liu, X.; Yang, M.J.; Wang, S.N.; Xu, D.; Li, H.; Peng, X.X. Differential antibody responses to outer membrane proteins contribute to differential immune protections between live and inactivated Vibrio parahemolyticus. J. Proteome. Res. 2018, 9, 2987–2994. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Kong, X.Y.; Shi, S.; Zheng, X.L.; Guo, G.; Wei, Y.Q. Preparation of alginate coated chi-tosan microparticles for vaccine delivery. BMC. Biotechnol. 2008, 8, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Marslin, G.; Prakash, J.; Qi, S.S.; Franklin, G. Oral delivery of curcumin polymeric nanoparticles ame-liorates CCl4-induced subacute hepatotoxicity in wistar rats. Polymers 2018, 5, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chen, C.L.; Chen, C.; Marslin, G.; Ding, R.; Wu, S.Q. Construction and evaluation of a novel triple cell epitope-based polypeptide vaccine against cow mastitis induced by Staphylococcus aureus, Escherichia coli and Streptococcus. Trop. J. Pharm. Res. 2017, 10, 2477–2486. [Google Scholar] [CrossRef]
- Meehan, T.L.; Yalonetskaya, A.; Joudi, T.F.; McCall, K. Detection of cell death and phagocytosis in the drosophila ovary. Methods Mol. Biol. 2015, 1328, 191–206. [Google Scholar] [PubMed]
- Liu, X.; Chen, C.; Zhang, X.Y. Drug-drug interaction of acetaminophen and roxithromycin with the cocktail of cytochrome P450 and hepatotoxicity in rats. Int. J. Med. Sci. 2020, 3, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paunovska, K.; Loughrey, D.; Sago, C.D.; Langer, R.; Dahlman, J.E. Using large datasets to under-stand nanotechnology. Adv. Mater. 2019, 43, e1902798. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Anselmo, A.C.; Huang, L. Nanotechnology intervention of the microbiome for cancer therapy. Nat. Nanotechnol. 2019, 12, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yang, S.; Jiang, Z.; Zhou, J.; Yao, J. Self-propelled gemini-like LMWH-scaffold nanodrugs for overall tumor microenvironment manipulation via macrophage reprogramming and vessel normaliza-tion. Nano Lett. 2020, 1, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Xu, M.; Zhao, T.; Kuang, L.; Hua, D. Smart oral administration of polydopa-mine-coated nanodrugs for efficient attenuation of radiation-induced gastrointestinal syndrome. Adv. Healthc. Mater. 2020, 13, e1901778. [Google Scholar] [CrossRef] [PubMed]
- Muxika, A.; Etxabide, A.; Uranga, J.; Guerrero, P.; de la Caba, K. Chitosan as a bioactive polymer: Processing, properties and applications. Int. J. Biol. Macromol. 2017, 2, 1358–1368. [Google Scholar] [CrossRef]
- Zhang, E.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Li, P. Advances in chitosan-based nanoparticles for on-cotherapy. Carbohydr. Polym. 2019, 222, 115004. [Google Scholar] [CrossRef] [PubMed]
- Matias, L.L.R.; Costa, R.O.A.; Passos, T.S.; Queiroz, J.L.C.; Serquiz, A.C.; Maciel, B.L.L. Tamarind trypsin inhibitor in chitosan whey protein nanoparticles reduces fasting blood glucose levels with-out compromising insulinemia: A preclinical study. Nutrients 2019, 11, 2770. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.Q.; Jiang, W.F. Preparation of chitosan-based nanoparticles for enzyme immobilization. Int. J. Biol. Macromol. 2019, 126, 1125–1132. [Google Scholar] [CrossRef]
- Dubey, S.; Avadhani, K.; Mutalik, S.; Sivadasan, S.M.; Maiti, B.; Girisha, S.K. Edwardsiella tarda OmpA encapsulated in chitosan nanoparticles shows superior protection over inactivated whole cell vaccine in orally vaccinated fringed-Lipped Peninsula Carp (Labeo fimbriatus). Vaccines 2016, 4, 40. [Google Scholar] [CrossRef]
- Lu, M.; Li, L.Y.; Lu, S.; Li, K.; Su, Z.; Wang, Y. The protective effect of dexmedetomidine on LPS-induced acute lung injury through the HMGB1- mediated TLR4/NF-κB and PI3K/Akt/mTOR pathways. Mol. Immunol. 2018, 94, 7–17. [Google Scholar]
- Ezz-Eldin, Y.M.; Aboseif, A.A.; Khalaf, M.M. Potential anti-inflammatory and immunomodulatory effects of carvacrol against ovalbumin-induced asthma in rats. Life Sci. 2020, 242, 117222. [Google Scholar] [CrossRef] [PubMed]
- Alhusaini, A.M.; Faddah, L.M.; El Orabi, N.F.; Hasan, I.H. Role of some natural antioxidants in the modulation of some proteins expressions against sodium fluoride-induced renal injury. Biomed. Res. Int. 2018, 2018, 5614803. [Google Scholar] [CrossRef] [Green Version]
- Benfaremo, D.; Manfredi, L.; Luchetti, M.M.; Gabrielli, A. Musculoskeletal and rheumatic diseases induced by immune checkpoint inhibitors: A review of the literature. Curr. Drug. Saf. 2018, 3, 150–164. [Google Scholar] [CrossRef]
- Aaby, P.; Benn, C.S. Developing the concept of beneficial non-specific effect of live vaccines with epidemiological studies. Clin. Microbiol. Infect. 2019, 12, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, A.; Noiret, L.; Higgins, J.M. White blood cell population dynamics for risk stratification of acute coronary syndrome. Proc. Natl. Acad. Sci. USA 2017, 46, 12344–12349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solana, J.C.; Ramírez, L.; Cook, E.C.; Hernández-García, E.; Sacristán, S.; Martín, M.E. Subcutane-ous immunization of Leishmania HSP70-II null mutant line reduces the severity of the experimental visceral leishmaniasis in BALB/c mice. Vaccines 2020, 1, 141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.M.; Zhao, H.L.; Wang, J.J.; Liao, Y.; Na, R.X.; Wang, L.C. Evaluation of immune respons-es and related patho-inflammatory reactions of a candidate inactivated EV71 vaccine in neonatal monkeys. Zhonghua Yi Xue Za Zhi 2011, 28, 1977–1981. [Google Scholar]
- Santos, K.O.; Costa-Filho, J.; Riet, J.; Spagnol, K.L.; Nornberg, B.F.; Kütter, M.T. Probiotic express-ing heterologous phytase improves the immune system and attenuates inflammator response in zebrafish fed with a diet rich in soybean meal. Fish Shellfish Immun. 2019, 93, 652–658. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| IFN-γ | TCAAGTGGCATAGATGTGGAAGAA | TGGCTCTGCAGGATTTTCATG |
| HSP70 | GAAGGTGCTGGACAAGTGC | GCCAGCAGAGGCCTCTAATC |
| TNF-a | TATGGCTCAGGGTCCAACTC | GCTCCAGTGAATTCGGAAAG |
| IL-6 | GACAAAGCCAGAGTCCTTCAGAGAGATACAG | TTGGATGGTCTTGGTCCTTAGCCAC |
| IL-10 | AACATACTGCTAACCGACTC | ATGCTCCTTGATTTCTGG |
| GAPDH | ACAGTCCATGCCATCACTGCC | GCCTGCTTCACCACCTTCTTG |
| Group | Compound | WBC No. (×109/L) | Thymus Index | Spleen Index | Phagocytic Percentage (PP %) | Phagocytic Index (PI %) |
|---|---|---|---|---|---|---|
| 1 | NP-OmpA | 6.83 ± 0.97 a | 2.67 ± 0.12 b | 4.67 ± 0.38 b | 5.76 ± 1.10 b | 3.56 ± 0.62 a |
| 2 | Control 1 (OmpA) | 6.93 ± 0.89 a | 2.25 ± 0.10 a | 4.01 ± 0.41 ab | 4.85 ± 0.33 ab | 3.67 ± 0.42 a |
| 3 | Control 2 (NP-Empty) | 6.73 ± 0.48 a | 2.08 ± 0.07 a | 3.48 ± 0.43 a | 4.68 ± 1.02 a | 3.08 ± 0.43 a |
| 4 | Control 3 (Normal saline) | 6.68 ± 0.67 a | 2.03 ± 0.11 a | 3.31 ± 0.25 a | 4.21 ± 0.98 a | 3.16 ± 0.58 a |
| Group | Compound | Nos | Survival No. | Death No. | ADR, % | RPS, % |
|---|---|---|---|---|---|---|
| 1 | NP-OmpA | 15 | 11 | 4 | 26.67 | 71.43 ** |
| 2 | Control 1 (OmpA) | 15 | 5 | 10 | 66.67 | 28.57 |
| 3 | Control 2 (NP-Empty) | 15 | 2 | 13 | 86.67 | 7.14 |
| 4 | Control 3 (Normal saline) | 15 | 1 | 14 | 93.33 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Sun, W.; Wu, N.; Rong, N.; Kang, C.; Jian, S.; Chen, C.; Chen, C.; Zhang, X. Synthesis of Escherichia coli OmpA Oral Nanoparticles and Evaluation of Immune Functions against the Major Etiologic Agent of Cow Mastitis. Vaccines 2021, 9, 304. https://doi.org/10.3390/vaccines9030304
Liu X, Sun W, Wu N, Rong N, Kang C, Jian S, Chen C, Chen C, Zhang X. Synthesis of Escherichia coli OmpA Oral Nanoparticles and Evaluation of Immune Functions against the Major Etiologic Agent of Cow Mastitis. Vaccines. 2021; 9(3):304. https://doi.org/10.3390/vaccines9030304
Chicago/Turabian StyleLiu, Xiang, Wei Sun, Nana Wu, Na Rong, Chao Kang, Sijie Jian, Chunlin Chen, Chen Chen, and Xiaoying Zhang. 2021. "Synthesis of Escherichia coli OmpA Oral Nanoparticles and Evaluation of Immune Functions against the Major Etiologic Agent of Cow Mastitis" Vaccines 9, no. 3: 304. https://doi.org/10.3390/vaccines9030304